- Artenpool-Hypothese
-
Die Artenpool-Hypothese (engl. Species Pool Hypothesis) entstand Ende des letzten Jahrhunderts als alternativer Top-down-Ansatz zu den zuvor propagierten Bottom-up-Erklärungsversuchen für die Artenvielfalt auf Gemeinschaftsebene (Biozönosen). Sie besagt generell [1], dass die Artenanzahl auf einer kleineren Skala im Wesentlichen durch die verfügbaren Arten (Artenanzahl) auf der nächsthöheren Skala bestimmt wird.
Inhaltsverzeichnis
Entstehung der Hypothese
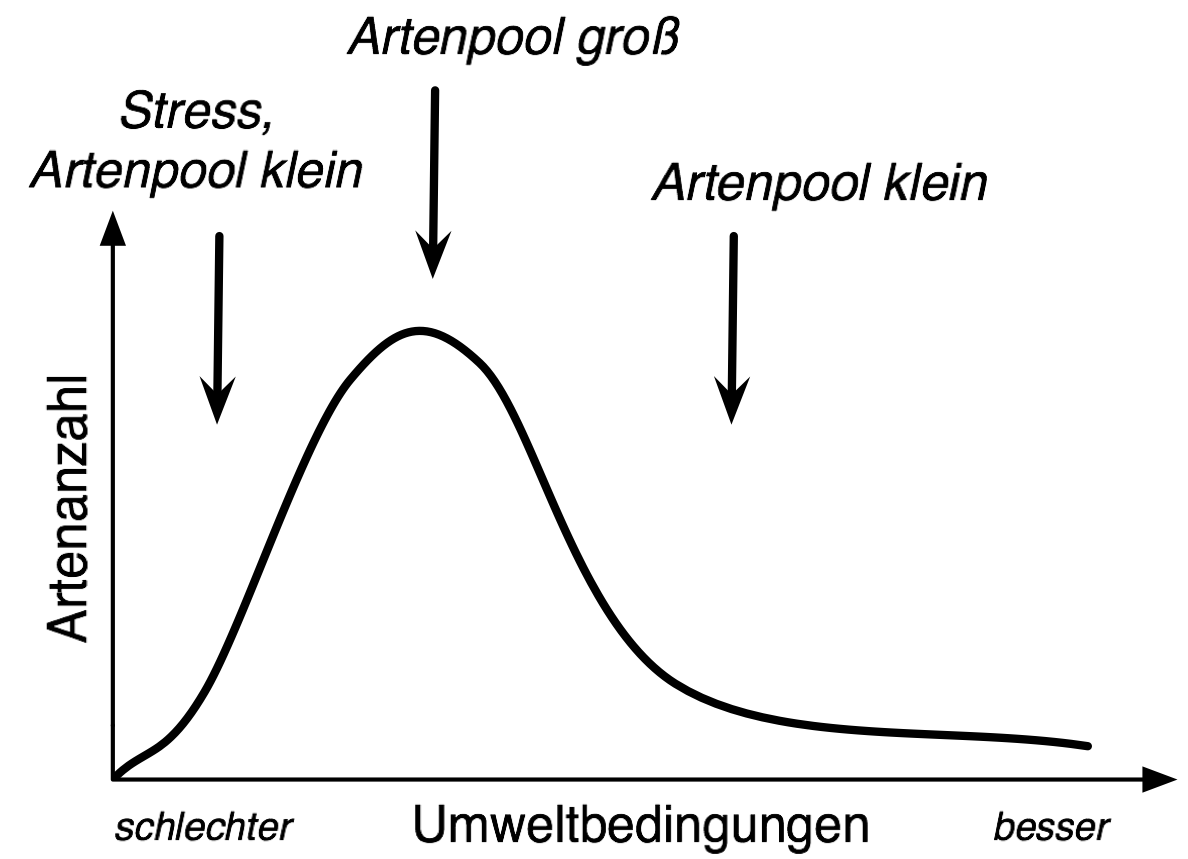
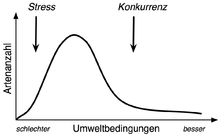 „Buckelkurve“: Muster des Zusammenhangs zwischen der Artenzahl von Gemeinschaften und den Umweltbedingungen, unter denen diese vorkommen.
„Buckelkurve“: Muster des Zusammenhangs zwischen der Artenzahl von Gemeinschaften und den Umweltbedingungen, unter denen diese vorkommen.
Die Hypothese wurde im Kontext einer Diskussion des C-S-R-Modells entwickelt [2]. Ausgangspunkt ist der unimodale Kurvenverlauf, der sich beim Zusammenhang zwischen Artenanzahl von Gemeinschaften und den Umweltbedingungen (Standortbedingungen), unter denen diese vorkommen, zeigt. Dieses, als „Buckelkurve“ (engl. hump-backed curve) bezeichnete Muster wird im Allgemeinen durch kleinskalige Prozesse erklärt: Unter „schlechten“ (weniger fruchtbar/unfruchtbar) Bedingungen ist der (Umwelt-)Stress so groß, dass sich nur wenige Arten ansiedeln können. Verbessern sich die Bedingungen finden sich zunehmend Arten ein. Verbessern sich die Bedingungen weiter, so kommt es zunehmend zu zwischenartlicher Konkurrenz und zum Konkurrenzausschluss – die Artenanzahl sinkt, da sich wenige konkurrenzstarke Arten durchsetzen.
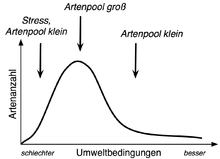 Erklärung der „Buckelkurve“ durch die Artenpool-Hypothese.
Erklärung der „Buckelkurve“ durch die Artenpool-Hypothese.Alternativ ist es allerdings ebenso möglich, dass es schlicht weniger Arten gibt, die an fruchtbare Standorte angepasst sind. Auf der linken Seite der Kurve werden Arten somit nicht durch Konkurrenten verdrängt, sondern es kommen deshalb weniger Arten vor, weil der Artenpool (eine Gruppe von Arten, die potentiell unter den betrachteten Bedingungen vorkommen können) geringer ist. Als Begründung wird angeführt, dass fruchtbare Standorte seltener sind und deshalb im Laufe der Evolution nur eine geringe Möglichkeit zur Ausbildung von Anpassungsmerkmalen (z. B. hohe potentielle Biomasse) an diese Bedingungen bestand. Da intermediäre Standorte weiter verbreitet sind, konnten sich hier entsprechend mehr Arten an die Bedingungen anpassen – der Artenpool ist somit größer. Arme Standorte, auf der anderen Seite, waren zwar zu Beginn der Besiedlung des Landes dominierend (bzw. die überall herrschenden Standortverhältnisse), wurden aber zunehmend in intermediäre bis hin zu fruchtbaren Standorten umgewandelt und sind heute ebenfalls „selten“. Außerdem sind bei armen Standorten weiterhin die Ressourcen limitierend.
Neben der Fläche die ein Standort- oder Habitattyp einnimmt, ist auch das Alter einer solchen Fläche wichtig. Mit zunehmendem (geologischem) Alter steigt ebenfalls die Wahrscheinlichkeit einer Anpassung und damit die Größe des Artenpools (Artenakkumulation). Beispiele sind hier der tropische Regenwald und die Sonora-Wüste. Beides sind sehr alte, bzw. über lange Zeit relativ stabile Lebensräume und unabhängig von ihrer unterschiedlichen Fruchtbarkeit sehr artenreich. Ein weiteres Beispiel ist die Artenarmut europäischer temperater Laubwälder in Relation zu den Wäldern der gleichen Vegetationszone in Nordamerika und Ostasien. Durch die Vereisung im Pleistozän kam es in Europa zu größeren Artverlusten als in den beiden anderen Regionen, da hier Gebirge wie die Alpen und Pyrenäen, sowie das Mittelmeer, Barrieren für den Rückzug und die Wiedereinwanderung von Arten bildeten.
Definitionen
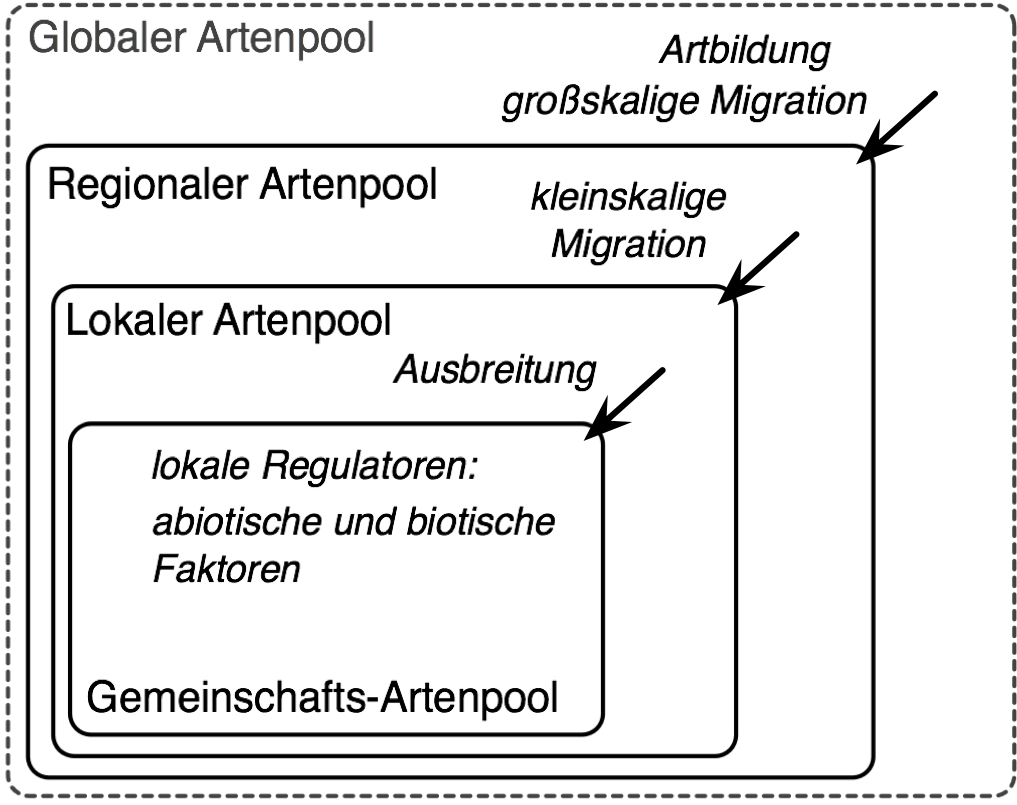
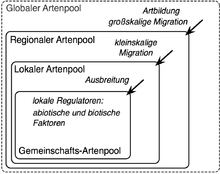 Verschiedene Skalen der Artenpools mit ihren bestimmenden Prozessen.
Verschiedene Skalen der Artenpools mit ihren bestimmenden Prozessen.Das Artenpool-Konzept kann man als in sich verschachtelte Pools auf verschiedenen Skalen verstehen. Die Größe des äußeren Pools bestimmt dabei die Größe des jeweils nachfolgenden Pools, wobei jede Ebene durch bestimmte Prozesse charakterisiert ist. Die höchste Ebene stellt der globale Artenpool dar. Dieser wurde bisher nicht in der Literatur benannt, soll hier aber als weltweit alle Arten umfassender Pool ergänzt werden (je nach Intension kann hier eine bestimmte Organismengruppe wie Pflanzen, Tiere oder Gefäßpflanzen) betrachtet werden. Auf dieser globalen Skala sind Artbildung und großskalige Migration grundlegende Prozesse, sie bestimmen welche Arten in den regionalen Artenpool (die darunterliegende Ebene) einwandern können. Der regionale Artenpool ist eine Gruppe von Arten, die in einer bestimmten Region vorkommt und welcher es möglich ist in der betrachteten Zielgemeinschaft zu koexistieren. Eine Region ist in diesem Zusammenhang ein physiografisch und klimatisch relativ einheitliches Gebiet, aus welchem die Zielgemeinschaft erreichbar ist. Für die Region existiert des Weiteren eine Flora. Die nächste Ebene wird als lokaler Artenpool bezeichnet, d. h. eine Gruppe von Arten, die in der Landschaft rund um die Zielgemeinschaft vorkommen. Den Arten ist es möglich in der Zielgemeinschaft zu koexistieren und sie können in diese relativ schnell (innerhalb weniger Jahre) einwandern. Der Gemeinschafts-Artenpool (auch aktueller Artenpool) umfasst schließlich alle Arten, die aktuell in der Zielgemeinschaft präsent sind. An der Grenze zwischen dem lokalen und Gemeinschafts-Artenpool bestimmten abiotische und biotische Bedingungen als ökologische Filter welche Arten in die betrachtete Gemeinschaft einwandern.
Kritik
Ein wesentlicher Kritikpunkt ist, dass sich die Artenpool-Hypothese zu stark auf den Top-Down-Ansatz beschränkt. Der Einfluss des Artenpools sollte nicht als alleiniger bestimmender Faktor im Bezug zur lokalen Artenvielfalt angesehen werden. Vielmehr ist er als Ergänzung zu lokalen Mechanismen wie Einnischung und Konkurrenz zu sehen [3]. Es wird außerdem darauf hingewiesen, dass Richtung des Zusammenhangs zwischen lokaler und regionaler Artenvielfalt sowie die dahinter stehenden Mechanismen unbekannt sind [4], [5].
Als zweiter Schwachpunkt der Hypothese wird ihre Testbarkeit angesehen [6]. Aus statistischen Gründen dürfen die Pools der Gemeinschaften in Teststudien sich nicht überlappen (müssen unabhängig sein). Trotzdem müssen sie ähnliche Taxa mit ähnlichen Standortansprüchen enthalten um vergleichbar zu sein. Dies ist aber für die meisten Organismengruppen praktisch nicht umsetzbar. Teilweise könnte mit je einer Testgemeinschaft pro Kontinent gearbeitet werden, allerdings ist die Zahl an Kontinenten und somit Stichproben dann zu gering um statistischen Anforderungen zu genügen. Bisherige Studien (z. B. [7], [8], [9], [10]) sind diesbezüglich unbrauchbar. Wenn nicht testbar, wird allerdings auch die gesamte Hypothese unbrauchbar.
Literatur
- ↑ Zobel (1997): The relative role of species pools in determining plant species richness: an alternative explanation of species coexistance? Trends in Ecology & Evolution 12, S. 266-269
- ↑ Taylor, Aarssen, Loechle (1990): On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies. Oikos 58, S. 239-250
- ↑ Grace (2001): Difficulties with estimating and interpreting species pools and the implications for understanding patterns of diversity. Folia Geobotanica 36, S. 71-83
- ↑ Herben (2001): Correlation between richness per unit area and the species pool cannot be used to demonstrate the species pool effekt. Journal of Vegetation Science 11, S. 123-126
- ↑ Lepš (2001): Species-pool hypothesis: limits to its testing. Folia Geobotanica 36, S. 45-52
- ↑ Wilson, Anderson (2001): Species-pool relations: like a wooden light bulb? Folia Geobotanica 36, S. 35-44
- ↑ Pärtel, Zobel M., Zobel K., van der Maarel (1996): The species pool and ist relation to species richness: evidence from the Estonian plant communities. Oikos 75, S. 111-117
- ↑ Duncan, Buckley, Urlich, Stewart, Geritzlehner (1998): Small-scale richness in forest canopy gaps: the role of niche imitation versus the size of the species pool. Journal of Vegetation Science 9, S. 455-460
- ↑ Pärtel, Zobel (1999): Small-scale plant species richness in calcareous grasslands determined by the species pool, community age and shoot density. Ecography 22, S. 153-159
- ↑ Safford, Rejmánek, Hadač (2001). Species pools and the “hump-back” model of plant species diversity: an empirical analysis at a relevant spatial scale. Oikos 95, S. 282 – 290
Kategorie:- Theoretische Ökologie
Wikimedia Foundation.