- Innovation (Evolution)
-
Eine phänotypische Innovation ist ein neues Merkmal oder eine neue Verhaltensweise in der evolutionären Stammesgeschichte, das sich nicht allein als Variation bestehender Merkmale der Vorfahren erklären lässt. Innovation steht somit in Abgrenzung zu Variation, der graduellen Veränderung eines bereits vorhandenen Merkmals.
Unterscheidung Variation und Innovation
Innovation in der Evolution wird unterschieden von Variation, jeweils bezogen auf den Phänotyp. Variation wird von Charles Darwin und der der Synthetischen Evolutionstheorie hauptsächlich mit natürlicher Selektion und Adaptation in Verbindung gebracht. Die Synthetische Theorie analysiert auf Basis populationsgenetischer Betrachtung statistische Änderungen der Genfrequenz in Populationen und interessiert sich in ihrer klassischen Form nicht für Fragen der Entstehung spezifischer Merkmale des Organismus. Phänotypische Variation wird in der Standardtheorie als gegeben angenommen.
Im Gegensatz dazu wird besonders von Evo-Devo seit Anfang der 1990er Jahre untersucht, wie die Embryonalentwicklung hilft, das Entstehen innovativer Merkmale unter ökologischen Bedingungen zu erklären und ihre Realisierung im Organismus sowie ihre dauerhafte Verankerung (genetisch/epigenetische Integration) zu verstehen.
Allgemein wird der Begriff Innovation jedoch oft synonym mit Variation verwendet, wenn wichtige evolutionäre Veränderungen beschrieben werden. Wegen der oft unscharfen Abgrenzungen kann es zu Missverständnissen kommen[1]. Innovation meint immer eine qualitative Abweichung von den Vorgängereigenschaften, und nicht nur eine quantitative (Müller)[1].
Beispiele
 Abb. 1 Der Schildkrötenpanzer ist eine evolutionäre Innovation vom Typ II. Sie erforderte parallel komplizierte Skelettumbauten.
Abb. 1 Der Schildkrötenpanzer ist eine evolutionäre Innovation vom Typ II. Sie erforderte parallel komplizierte Skelettumbauten.
Wirkliche Innovationen treten in der Evolution seltener als Variationen auf. Ihre Bedeutung für die Evolution ist jedoch oft groß, da in der Evolutionsgeschichte mit einer Innovation, speziell mit einer Schlüsselinnovation (s.u.), vielfach eine lang anhaltende adaptive Radiation auf höheren taxonomischen Ebenen auftritt. Folgende Merkmale werden in der Literatur als phänotypische Innovation bezeichnet:
- Feder
- Milchdrüse bei Säugetieren
- Außenskelett
- Innenskelett und spezifische Skelettelemente, z.B. Schulterblatt
- Zweibeinigkeit (Bipedie)
- Schildkrötenpanzer
- Backentasche von Nagetieren
- Gliedmaßen und Fortsätze von Gliederfüßern (Arthropoden) wie Antennen/Fühler, Greif-/Beiß-Schneidewerkzeuge und Gliedmaßen für unterschiedliche Fortbewegung
- Insektenflügel
- Flügelmuster bei Schmetterlingen
- Blütenorgane (Blüte)
Physiologie[1]:
- Konstante Körpertemperatur bei Warmblütern
Verhalten:
- Werkzeugverwendung bei Vögeln, Primaten, Delfinen u.a[4]
- Laube der Laubenvögel
- Brutpflege
- Gemeinsame elterliche Aufzucht der Jungen bzw. Aufzucht in der Gruppe (cooperive breeding)[5]
- Pilzfarmbetrieb von Blattschneiderameisen
- Sprache
- Kultur des Menschen[6]
Definitionen und Abgrenzungen
Ernst Mayr, einer der Hauptvertreter der Synthetischen Theorie, sieht 1963als evolutionäre Innovation „jede neu erworbene Struktur oder Eigenschaft, die es erlaubt eine neue Funktion auszuüben, die ihrerseits eine neue adaptive Zone eröffnet“ [7]. Diese Definition lässt auch zu, dass eine neue Funktion durch reine quantitative Anhäufung bereits bestehender Merkmalselemente zustande kommt und nicht nur durch qualitative Neuerung. Heute wird eher letzteres mit Innovation gleichgesetzt.
Stärker auf die Entwicklung, also auf den Entstehungsweg der Innovation (novelty) heben Müller/Wagner 1991 ab. Sie definieren Innovation als „ein Konstruktionselement in einem Bauplan, das weder ein homologes Gegenstück in der Vorläuferart noch im selben Organismus hat“[8]. Diese Definition schließt rein quantitative Veränderung von schon bestehenden Merkmalen aus. Sie erlaubt den Blick auf Merkmale, die entweder gänzlich neu entstehen oder aus neuen Kombinationen oder Unterteilungen bereits bestehender Strukturen hervorgehen (z.B. der Daumen des Panda). Ferner erlaubt die Definition vom Müller/Wagner die Anwendung auf klar abgegrenzte, eindeutige Fälle nicht nur in der Morphologie sondern auch in der Physiologie oder im Verhalten[9]. Ähnlich spricht West-Eberhard von „einem neuen Merkmal, das auf einer qualitativ neuen Entwicklungsvariante basiert“ [10]. Um die Ökologie einzubeziehen und zu berücksichtigen, dass nicht nur auf diskontinuierliche Merkmale eingegangen wird, erklärt Massimo Pigliucci 2008 Innovationen wie folgt: „Evolutionäre Innovationen sind neue Merkmale oder Verhaltensweisen, oder neue Kombinationen zuvor existenter Merkmale oder Verhaltensweisen, die in der Evolution einer Abstammungslinie entstehen, und die eine neue Funktion in der Ökologie dieser Abstammungslinie zeigen“ [11]. Diese Definition lässt wiederum den Funktionswechsel stärker in den Vordergrund treten. Ferner soll die aus Sicht von Evo-Devo hervorgehobene Diskontinuität nicht überbetont werden und auch eher kontinuierliche Konstruktionsänderung in der Evolutionstheorie beinhaltet sein.
Der Begriff Innovation wird im Umfeld von Evolution neben den genannten Feldern von Morphologie, Physiologie und Verhalten auch in folgenden Gebieten verwendet:
- Genprodukte (Proteine)
- Genetik (genetische/genomische Innovation) und
- Gennetzwerke
- Genregulation
Schlüsselinnovation
Schlüsselinnovation (key innovation) bezieht sich auf die herausragende Rolle bestimmter Innovationen im makroevolutionären Prozess und — wie bei Mayr ebenfalls — auf die adaptive Radiation. Beispiele für Schlüsselinnovationen sind die Zweibeinigkeit, das Auge, Milchdrüsen bei Säugetieren und andere. Schlüsselinnovation ist vom Begriff her stets in einer wichtigen reziproken Verbindung zwischen ihrer internen Konstruktion, den entsprechenden Umweltfaktoren und der Population zu sehen[1]. In diesem Zusammenhang ermöglichen sie die Verwertung neuer Energieresourcen und die radiative Exploration neuer ökologischer Nischen[1]. Das wird deutlich am Beispiel der Vögel in der Folge der Innovation der Flügel und der Zahl von mehr als 9000 rezenten Vögeln. Nennt man als Schlüsselinnovation taxonomisch neutral den Flügel, ist die Bedeutung noch umfangreicher, da Flügel mehrmals unabhängig evolutionär entstanden sind (Vogel, Fledertiere, Insekt, Schmetterling) und so zur adaptiven Radiation von noch weit mehr Nischen geführt haben.
Innovation zielt also stärker auf die konstruktive Entstehung ab, während Schlüsselinnovation stärker die Diversität in der Folge kaskadierter Effekte einer Innovation meint [12] [13].
Klassifizierung von Innovationsformen
Gerd B. Müller unterscheidet 3 Typen phänotypischer Innovation[14]:
- Typ I Innovation: Primäre anatomische Architektur von Metazoa
- Typ II Innovation: Diskretes neues Element, das einem existierenden Bauplan hinzugefügt wird
- Typ III Innovation: Extreme Variation eines existierenden Bauplanmerkmals
Typ I Innovation beruht auf physikalischen Grundvoraussetzungen für die unterschiedlichen Aggregationsformen von Zellen in multizellulären Organismen (Adhäsion, Diffusion, Oszillation etc.) wie von Stuart A. Newman als Dynamic Patterning Modules (DPM´s) beschrieben [15]. Solche multizellulären Grundformen können evolutionär als prämendelisch verstanden werden, die erst ab einer bestimmten Skalierung emergent auftreten. Strukturen werden noch nicht strikt an den Genotyp vererbt, sondern ihre Realisierung in jeder Generation wäre stark von den jeweils herrschenden Umweltbedingungen bestimmt.
Typ II – Aus der Sicht von Evo-Devo ist besonders dieser Innovationstyp von Bedeutung wegen der qualitativen Aspekts[16]. Beispiele für Innvotation vom Typ II sind der Schildkrötenpanzer oder die Feder.
Typ III Innovation: Hier geht das neue Merkmal aus einem bestehenden phänotypischen Merkmal hervor, z.B. der Narwal-Stoßzahn. Die Innovation kann größere neue Funktion besitzen und auf taxonomische Apomorphie hinauslaufen. Wenn in der Entwicklung neue Konstruktionselemente erscheinen, kann in einer solchen qualitativen, phänotypischen Innovation Standardvariation nicht als Ursache gesehen werden; es bedarf vielmehr einer spezifischen Erklärung für ihr Entstehen[17]
Die Entwicklung als Hauptfaktor der phänotypischen Innovation
Es werden drei Faktoren evolutionärer phänotypischer Innovation unterschieden:[18]
Initiierungsbedingungen
"Die Selektion kann nicht an Merkmalen angreifen, die noch nicht existieren und somit nicht unmittelbar Innovation verursachen" [19]. West-Eberhard betont: In den überwiegenden Fällen sind es veränderte Umweltbedingungen, die den Anstoß für evolutionäre Innovation geben[20]). Das gilt sowohl für die Entstehung des Skeletts der Wirbeltiere [21] als auch für die Entstehung des Außenskeletts von Meeresbewohnern (Seeigel), letzteres bei Anreicherung des Wassers durch Calcium [22]. Die Begründung dafür, dass Umweltfaktoren als initiierende Stressoren überwiegen, sieht West-Eberhard darin, dass sie oft viele Generationen lang anhalten und breit wirken, unter Umständen auf die gesamte Population lange und gleichzeitig. Das erleichtert die evolutionäre Entstehung von Innovation.
 Abb. 2 Der vier bis fünf Meter lange Stoßzahn des Narwals ist eine Innovation vom Typ III
Abb. 2 Der vier bis fünf Meter lange Stoßzahn des Narwals ist eine Innovation vom Typ IIIMögliche initiierende Faktoren, durch die phänotypische Innovation ausgelöst werden kann[1]:
1. Seltene genetische Mutationen, die das Rohmaterial bilden für einen spontanen Struktur-Effekt, der sich in der Entwicklung herausbildet. Möglicherweise können solche Mutationen der Entstehung der Feder zugrunde gelegen haben, neue makroskopische Eigenschaften, die auf neuartigem Material (φ-Keratine) basieren.
2. Symbiotische Vereinigung von zuvor separierten genetischen oder entwicklungsbiologischen Komponenten. Endoysymbiose und auch horizontaler Gentransfer sind vor allem bei der evolutionären Entstehung von Eukaryoten bekannt. Der Transfer beschränkt sich nicht nur auf kurze Gensegmente sondern kann auch die Übertragung vollständiger genetischer Module bzw. Entwicklungsmodule zwischen zwei Arten beinhalten.
3. Epigenetische Nebenprodukte (side effect hypothesis) sind Strukturen. die entstehen,wenn Selektion an anderen Merkmalen oder auf anderen Ebenen ansetzt als an jenen, die zur Innovation selbst führen. Auf Nebeneffekte der Selektion hat bereits Darwin 1859 hingewiesen[23]. Selektion kann zum Beispiel auf Parameter wie Zellteilungsraten, Zell- und Gewebe-Signalaustausch oder das Timing unterschiedlicher entwicklungsbiologischer Prozesse wirken und dabei das System an einen Schwellenwert bringen, an dem neue Strukturen automatisch aus den Reaktionsmöglichkeiten der betroffenen Entwicklungsmodule hervorgehen. Beispiele epigenetisch entstandener Nebenprodukte sind etwa neue Skelettelemente. Die Entstehung der Vogelfeder wird von Josef H. Reichholf auf diese Art erklärt[24]. Reichholf betont z. B. den Überschuss an Nahrungsbestandteilen als möglichen Initiator für die Innovation.
Realisierungsbedingungen
Sofern die Initiierungsbedingungen für die Innovation unspezifisch und allgemein sind (Ernährung etc.), müssen die Realisierungsbedingungen zwangsläufig in der Entwicklung liegen [25]. Hier sieht man das Phänomen von Schwellenwerteffekten, das bedeutet, dass kleine Ausgangsbedingungen nicht-lineare, phänotypische Variation/Innovation hervorrufen können [26]. Die Veränderung von einem einzigen Parameter führt zur Antwort des ganzen Systems Entwicklung auf den Störfaktor. Diese Antwort des Systems Entwicklung kann Innovationen erzeugen. Der Innovations-/Konstruktionsprozess bis einem der oben genannten Beispiele wird dabei in vielen Fällen mehrstufig sein, d.h die Phasen Initiierung - Realisierung - Integration werden mehrmals durchlaufen und Funktionswechsel (s.u.) treten gegebenenfalls mehrfach auf.
Integrations- und Fixierungsbedingungen
Hier fragt man, wie das neue phänotypische Element (zusätzl. Finger etc.) genetisch und epigenetisch fixiert bzw. assimiliert wird, so dass es dauerhaft Bestand hat und von dem auslösenden (Umwelt-)Stressor ganz oder nahezu entkoppelt wird. "Das Innovationsmerkmal muss in das bereits bestehende Konstruktions-, Entwicklungs- und Genom-System verankert werden, um Funktionalität und Vererbung sicherzustellen" [27]. Dabei "scheint die Regel zu sein, dass die epigenetische Integration der genetischen Integration vorausgeht"[28] oder wie West-Eberhard es ausdrückt: "Genes are followers in Evolution" [29]. "Die genetische Integration stabilisiert und fixiert neu generierte Merkmale und resultiert in einem immer engeren Mapping zwischen Genotyp und Phänotyp" (Müller)[30].
Umweltfaktoren spielen eine wesentliche Rolle in allen Phasen der Innovation. Müller spricht daher auch von epigenetischer Innovation[31]. Ein geringfügiger Störfaktor, entweder ein Umweltstressor oder eine genetische Mutation, kann eine Antwort des gesamten Systems Entwicklung provozieren, wenn die kanalisierte Plastizität an ihre Grenzen stößt. Diese Antwort kann auf Grund der Fähigkeit des Entwicklungssystems zur Selbstorganisation nicht-linear bzw. nicht-graduell (diskontinuierlich) sein. Die spezifische Ausprägungsform des morphologischen Ergebnisses wird in diesem Szenario nicht durch die Selektion diktiert, sondern durch die Reaktionsnorm des Entwicklungssystems[32].
Die Entstehung von Innovationen ist demnach kein adaptiver Prozess sondern natürliche Selektion wirkt erst sekundär auf ein Konstruktionsergebnis der Entwicklung. Das ist eine Kernaussage der epigenetisch verstandenen Evo-Devo-Forschung und -Theorie, wie sie von Müller, Kirschner und Gerhard, West-Eberhard vertreten wird.
Constraints
 Abb. 3 Der Insektenflügel ist eine evolutionäre, phänotypische Innovation Typ II. Er konnte erst nach mehrfachem Funktionswechsel als Flügel dienen (s. Evolution des Insektenflügels)
Abb. 3 Der Insektenflügel ist eine evolutionäre, phänotypische Innovation Typ II. Er konnte erst nach mehrfachem Funktionswechsel als Flügel dienen (s. Evolution des Insektenflügels)Constraints begrenzen die phänotypische Evolution und wirken richtungsbestimmend für ihren Verlauf. Sie können physikalisch, morphologisch oder phylogenetischer Natur sein. Es werden äußere und innere Constraints unterschieden. Im letzteren Zusammenhang sind besonders die Entwicklungsconstraints zu nennen. Conrad Hal Waddington nennt das Phänomen Kanalisierung[33]. Art und Umfang, wie Constraints aufgebrochen und überwunden werden können, spielen eine maßgebliche Rolle dafür, wie evolutionäre Innovation in der Entwicklung entstehen kann. Sind Entwicklungspfade stark kanalisiert, besteht im Sinn Waddingtons eine Pufferung genetischer Mutation, die auf die Erhaltung des status quo im Phänotyp hinwirkt. Das kann bedeuten, dass die Entwicklung auch bei hohem Selektionsdruck unfähig ist, mit Variation zu antworten und genau deswegen gezwungen ist, Schwellenwerte zu überschreiten, was zu erhöhten Chancen für Innovation führt[34].
Funktionswechsel
Komplexe Innovation sind vielfach nicht in den Funktionen entstanden, die sie bei rezenten Organismen erfüllen. So sind Insektenflügel mehrstufig entstanden, zunächst vermutlich als rudimentäre Körperfortsätze, die aus überschüssigem Material gewachsen sein könnten und im ersten Stadium aquatischen Arthropoden als Lamellen dienten. Später konnten sie als Segel beim Gleiten auf glatten Oberflächen verwendet werden und erst darauf aufbauend, nach weiteren Transformationen, als Flügel[35]. Stephen J. Gould schreibt zum Thema Funktionswechsel[36] „Überall in der Natur hat beinahe jeder Bestandteil jedes lebenden Wesen unter leicht veränderten Bedingungen wahrscheinlich unterschiedlichen Zwecken gedient und ist in der lebendigen Maschinerie vieler älterer und ganz spezieller Formen tätig geworden.“
Während solche Beispiele langfristige Prozesse beschreiben, lässt das oben genannte Evo-Devo-Konzept mit den Phasen Initiierung – Realisierung – Integration auf der Grundlage von Schwellenwerteffekten und der nicht-linearen Reaktionsfähigkeit der Entwicklungssysteme auch rasche morphologisch Übergänge zu (siehe dazu Evo-Devo).
Es ist davon auszugehen, dass die hier genannten Innovationsbeispiele, wie etwa Milchdrüsen oder Zweibeinigkeit, komplexe, mehrstufige Innovationsprozesse darstellen, die jeweils einzeln die spontane Konstruktionseigenschaft der Entwicklung erfüllen, aber erst in kaskadierten Stufen, gegebenenfalls mit mehreren Funktionswechseln, zu dem führen, als was sie in den Beispielen erkennbar sind [37]. Für Gerhard Vollmer ist es dabei unerlässlich, dass ein Merkmal zeitweise zwei oder mehr Funktionen gleichzeitig ausübt[38]. Vor allem Lewontin/Gould haben darauf aufmerksam gemacht, wie schwierig es ist, einzelnen Merkmalen und Verhaltensweisen eindeutige evolutionäre Funktion zuzuordnen und dass die Gefahr besteht, adaptionistisches Denken für jedes Merkmal und seine Funktionen unkritisch zu übernehmen[39] [40].
Verhaltensinnovation bei Tieren
 Abb. 4 2005 wurde erstmals beobachtet, wie ein Gorilla einen Stab zum Abtasten der Wassertiefe und als Gehhilfe verwendet.
Abb. 4 2005 wurde erstmals beobachtet, wie ein Gorilla einen Stab zum Abtasten der Wassertiefe und als Gehhilfe verwendet.Tiere erfinden innovative Verhaltensformen. Hier will man verstehen, wie diese evolutionär entstehen können und ob es gemeinsame Erklärungsmuster gibt. Unterschieden wird emotionales, kognitives, sozial und ökologisch neues Verhalten [41]. Raeder und Laland betrachten Verhaltensinnovation als Prozess und Produkt und definieren Innovation als "einen Prozess, der in neuem oder modifizierten erlernten Verhalten mündet und der die neuen Verhaltensvarianten in das Repertoire einer Population überführt"[42]. Innovatives Verhalten als Prozess beinhaltet individuelles und soziales Lernen. Hierbei wird soziales Lernen als chancenreicher für eine Etablierung in der Population gesehen. Raeder und Laland berichten daher bei innovativem Verhalten auch überwiegend über Primaten und Singvögel. Das generelle mentale und emotionale Makeup eines Individuums ist dabei relevant dafür, ob und wie oft Individuen mit neuen Verhaltensformen in Erscheinung treten [43]. Der niederländische Verhaltensforscher Carel van Schaik[4] nennt drei herausragende Muster für das Auftreten von Verhaltensinnovation bei sozialen Tieren:
- die Veranlagung eines Individuums
- das soziale Umfeld
- die ökologischen Umstände
Damit neue Verhaltensweisen entstehen können, ist laut van Schaik ökologischer Druck notwendig. Er ist eine mögliche Initiierungsbedingung (s.o.). Neue Verhaltensformen können dann unter Umständen überlebenswichtig für die Art werden. Das gilt vor allem dann, wenn eine Art nicht in der Lage ist, ihr Habitat zu verlassen oder zu vergrößern. "Eine große ökologische Verbreitung ist wahrscheinlich positiv korreliert mit Innovation, weil in einem ökologisch großen Gebiet neue Herausforderungen und Möglichkeiten bestehen. Dennoch muss Innovation nicht mit einer extensiven geografischen oder ökologischen Weite korelliert sein: Manchmal kann nämlich Innovation die Migrations- oder Erkundungstendenz der Besiedlung neuer Gebiete limitieren, weil Innovation die existierende Nische ausweitet bzw. eine effizientere Ausbeutung eröffnet und es so Individuen erlaubt zu bleiben. Es wird daher angenommen, dass eine asymmetrische Korrelation besteht zwischen der Ausbreitung einer Art und Innovation"[44].
Schwierigkeiten können sich nach Jablonka und Avital bei der eindeutigen Abgrenzung neuer kontextabhängiger, kognitiver Verhaltensformen von anderen vorhandenen Verhaltensformen und deren Funktionen ergeben. Lernfähigkeit und Bereitschaft für Innovation kann nicht losgelöst analysiert werden. Innovation resultiert aus dem hochintegrierten Zusammenspiel komplexer kognitiver Prozesse und Umstände. Deren zugrundeliegende physiologische und neurologische Mechanismen können nicht unterschieden werden von anderen solchen Prozessen, benötigen aber noch die Berücksichtigung einiger weiterer Prozesse, wie etwa bestimmte emotionale Zustände in spezifischen Lernsituationen[45].
Nicht geklärt ist heute die Frage, ob die Vererbung von sozialem Lernen eher konservatives oder innovatives Verhalten fördert, bzw. wann welches. Ausschlaggebend scheinen die Fragen nach den Entwicklungsdimensionen: In welchem Alter sind innovative Verhaltensformen am wahrscheinlichsten? Bei welchem Geschlecht, sozialen Status bzw. ökologischen Stress? Welche Anforderungen gibt es an kognitive Mechanismen, die nicht nur die Ausbildung erlernter neuer Innovationen erlauben sondern auch zu ihrer funktionalen Organisation führen? [46] So muss die fertige Laube des Laubenvogels zuerst bei älteren Tieren abgeschaut, dann selbst gebaut werden, anschließend für die Werbung des Weibchens genutzt werden, um nach mehrjährigen Misserfolgen und erst mit zunehmend verbesserter Bautechnik zu dem "Ziel" zu gelangen, dass eine Laube von einem Weibchen auch angenommen wird[47].
Die Verwendung eines Stocks durch einen einzelnen Gorilla (Abb. 4) ist noch keine evolutionäre Innovation. Dafür ist der Nachweis zu erbringen, dass die Verhaltensform in das Repertoire der Population übernommen wird (s.o. Definition). Die Vererbung (Weitergabe) von Verhalten beschreiben Jablonka und Lamb[48]. Dabei ist das Erkennen von kognitiven Fähigkeiten und Bereitschaftpotenzialen erforderlich, die sich wie oben gesagt, nicht losgelöst analysieren lassen. Tiere imitieren nicht in der Art und in dem Umfang, in dem Menschen imitieren, wie der US-amerikanische Anthropologe und Verhaltensforscher Michael Tomasello in seinen empirischen Studien belegen konnte[49]. Die Fähigkeit zu Imitation ist beim Menschen notwendig für die Entstehung von Kultur (s.u. Abschn. Kulturelle Evolution), Imitation kann demnach als eine menschliche Innovation gesehen werden. Tiere (Bsp. Laubenvogel) beobachten die Verhaltensprodukte und weniger die Verhaltensprozesse erfahrener Tiere und entwickeln daraus ihr eigenes ähnliches Verhalten[47]. Mit dem Aufzeigen derartiger Verhaltensformen und -unterschiede ist aber noch nicht nachgewiesen, dass oder wie das jeweilige Verhalten evolutionär (adaptiv) wirkt und wie es im darwinschen Sinn zur biologischen Fitness beiträgt. Die evolutionär adaptiven Eigenschaften von innovativen Verhaltensformen sind bei Tieren wenig untersucht. Die Evo-Devo-Sichtweise, dass Innovation ein Produkt der Entwicklung ist, erzwingt analog zu dem in Kap. 6 gesagten, auch bei innovativen Verhaltensweisen (novelties), ihre mögliche nicht-adaptive Entstehung im Zuge der Verhaltensentwicklung zu berücksichtigen. Den engen Zusammenhang von Entwicklung und Verhaltensänderung zeigt das Evo-Devo-Kapitel Ausgewählte empirische Forschungsergebnisse und dort der Abschnitt Belyaevs Zähmung von Silberfüchsen. Adaptives Verhalten ist jedoch Gegenstand der Theorie von Peter Richerson und Robert Boyd zur Evolution der menschlichen Kultur[6].
Kultur als Innovation
Die Kulturfähigkeit des Menschen wird als Alleinstellungsmerkmal unserer Spezies gesehen. Sowohl Sprache als auch Kultur selbst gelten als evolutionäre Innovationen[50]. Richerson und Boyd[6] beschreiben, wie Kultur evolutionär adaptiv entstehen kann. Es geht ihnen dabei nicht darum zu erklären, wie sich kulturelle Leistungen und Produkte entwickelt haben, wie etwa das Auto, das Internet oder gotische Kathedralen. Das wäre Evolution in der Kultur. Die Autoren meinen aber Evolution der Kultur. Auf den Unterschied hat Karl Eibl hingewiesen[51].
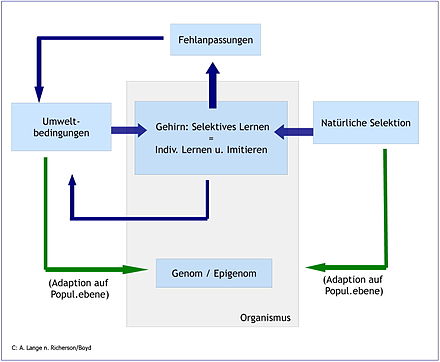 Abb. 5 Das interaktive System kultureller Evolution. Richerson und Boyd erklären, Kultur als Prozess komplexer Beziehungen von Genom/Epigenom, Umweltbedingungen, natürlicher Selektion und der Kumulation von vererbbarem Wissen.
Abb. 5 Das interaktive System kultureller Evolution. Richerson und Boyd erklären, Kultur als Prozess komplexer Beziehungen von Genom/Epigenom, Umweltbedingungen, natürlicher Selektion und der Kumulation von vererbbarem Wissen.Mögliche Initiierungsbedingungen für die Entstehung von Kultur werden in der Literatur auf breiter Basis beschrieben. Die Argumente reichen vom aufrechten Gang und der damit einhergehenden Verfeinerung der Hände, der Vergrößerung des Gehirns, dem Gebrauch von Werkzeugen, der Entwicklung der Sprache bis zur Erfindung der Landwirtschaft und Sesshaftigkeit. Alle Faktoren werden auch in ihrer Wechselwirkung diskutiert[52]. Für Richerson/Boyd sind vor allem die Fähigkeit des Menschen zu imitieren sowie die Fähigkeit, Wissen zu akkumulieren spezifisch menschlich und notwendige (nicht hinreichende) Voraussetzungen zur Entstehung von Kultur. Wolf Singer geht in dieser Frage weiter zurück auf die entscheidenden Schritte der Hirnevolution. Für ihn ist das Gehirn des "vorkulturellen" Menschen in seiner fundamentalen Struktur nicht anders als das des homo sapiens sapiens. Die Unterschiede in der "epigenetischen Gestaltung und Feinstruktur" und die "kognitiven Fähigkeiten, die durch diesen epigenetischen Weg erworben und tradiert werden" sieht Singer "mehr als eine Folge der kulturellen Evolution und des kollektiven Lernens und weniger (als) ihre Ursache"[53]. Somit gilt die Frage nach "der evolutionären Entwicklung von bestimmten kognitiven Funktionen..., die allein dem Menschen zukommen"[54]. Diese sieht Singer vor allem in der Fähigkeit, abstrakte, symbolische Fassungen. - sogenannte Metarepräsentation von kognitiven Inhalten zu erstellen""[54]. "Wir sind die einzige Spezies, die in der Lage ist, Dialoge zu führen, in denen es heißen kann: 'Ich weiß, dass du weißt, dass ich weiß' oder 'ich weiß, dass du weißt, was ich fühle'[55]. Singer beschreibt die neokortikalen Funktionen, die solches abstraktes Denken und ihre Repräsentation in unserem Gehirn möglich machen[56].
Wie Kultur realisierbar wird, beschreiben Richerson/Boyd wie folgt: Der Mensch ist zu kumulativer kultureller Evolution fähig. „Menschen können eine Innovation zu einer anderen hinzufügen, bis die Ergebnisse Organen mit extremer Perfektion ähneln wie etwa einem Auge“ [6]. Michael Tomasello hat das als Wagenhebereffekt bezeichnet[57]. Zweitens kombiniert der Mensch individuelles Lernen und soziales Lernen (Imitieren). Durch jede der beiden Lernformen kann Wissen akkumuliert werden. Erst aber durch die Möglichkeit des selektiven Lernens, das ist die jeweils unter Kosten-Nutzengesichtspunkten vorteilhafte Kombination der beiden Lernformen in Entscheidungssitutation, entsteht aber drittens akkumuliertes Wissen, das ein Objekt der natürlichen Selektion sein kann. Lernen per se ist also nicht als adaptiv interpretierbar. Mit anderen Worten: Lernen allein erhöht nicht die Fitness. Selbst wenn durch individuelles oder schulisches Lernen Wissen ständig vermehrt und weitergegeben wird, ist das nach Richerson und Boyd nicht nachweisbar adaptiv. Auf der Ebene der Art Mensch kann das allein die Selektion leisten, die die Individuen mit den jeweils vorteilhaftesten Kombinationen aus beiden Lernformen selektiert, wodurch die Population adaptiert.
Evolution durch Kultur definieren Richerson/Boyd wie folgt: „Unter kultureller Evolution verstehen wir Verhalten oder Artefakte, die über viele Generationen übermittelt und modifiziert werden und die zu komplexen Artefakten oder Verhalten führen“[6]. Durch Kultur verändert der Mensch seine eigenen Lebensbedingungen (Umwelt). Die veränderte Umwelt wirkt über epigenetische Prozesse auch auf das Genom zurück. Somit verändert und gestaltet der Mensch durch sein eigenes Handeln auch sein Genom und damit wiederum seine morphologische und verhaltensseitige Evolution (siehe dazu Nischenkonstruktion). Beide Mechanismen sind dicht verwoben und lassen sich nicht voneinander trennen. Somit kommen die Autoren zum Ergebnis: "Adaptation durch kumulative kulturelle Evolution ist kein Nebenprodukt der Intelligenz und des sozialen Lebens"[58].
Kultur selbst ist ein evolutionär entstandenes menschliches Merkmal. Das ist im Kern das Resultat der Sichtweise von Richerson und Boyd auf die beteiligten interaktiven Prozesse (s. Abb. 5). Kultur ist in dem Sinne ein evolutionärer Prozess, als der Mensch durch selektives Lernen jeweils entscheidet, ob er eher individuell oder eher durch Schule, Training etc. sozial lernt und so andere imitiert, wenn das kostengünstiger und effizienter ist. Das Ergebnis der Kombination der beiden Lernformen in der Interaktion von Mensch - Umwelt - Genom ist es, das im darwinschen Sinn nach Richerson und Boyd der Selektion unterliegt und die biologische Fitness des Menschen auf Populationsebene adaptiv verbessern kann. Temporäre Fehlanpassungen (maladaptions) wie Rückgang der Geburtenrate, Vernichtung der Ressourcen etc. können dabei nicht ausgeschlossen werden und widersprechen auch nicht dem Verständnis des darwinschen Evolutionsprinzips.
 Abb. 6 Wurfspeer der Anazasi-Kultur. Bereits die Herstellung eines Speers wie diesem sowie seine geübte Handhabung verlangen in hohem Grad die Vererbung von kumuliertem Wissen aus individuellem und sozialem Lernen über viele Generationen. Der Schaft muss exakt gerade gezogen sein, eine behauene, schlanke Steinspitze muss haltbar montiert, Länge, Ausrichtung, Balance und Wurftechnik der Waffe geduldsam erprobt sein. Die evolutionären Voraussetzungen für solche innovativen kulturellen, menschlichen Leistungen sind in Abb. 5 und im Text beschrieben.
Abb. 6 Wurfspeer der Anazasi-Kultur. Bereits die Herstellung eines Speers wie diesem sowie seine geübte Handhabung verlangen in hohem Grad die Vererbung von kumuliertem Wissen aus individuellem und sozialem Lernen über viele Generationen. Der Schaft muss exakt gerade gezogen sein, eine behauene, schlanke Steinspitze muss haltbar montiert, Länge, Ausrichtung, Balance und Wurftechnik der Waffe geduldsam erprobt sein. Die evolutionären Voraussetzungen für solche innovativen kulturellen, menschlichen Leistungen sind in Abb. 5 und im Text beschrieben.Siehe auch
Evolutionäre Entwicklungsbiologie
Einzelnachweise
- ↑ a b c d e f Müller, Gerd B. (2002): Novelty and Key Innovations. In Mark Pagel (Ed.): Encyclopedia of Evolution. Oxford University Press S. 827-830
- ↑ Müller, G. B. Epigenetic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 311Press
- ↑ Pigliucci, Massimo (2008) What, if anything, Is an Evolutionary Novelty? Philosophy of Science 75 (Dec. 2008) S. 896
- ↑ a b Schaik, Carel van (2006): Die Evolution der Innovation.Technology Review-Archiv, 2/2006, Seite 88
- ↑ Carel van Schaik (2008): Gruppen machen schlau.DIE ZEIT, 6. November 2008 Nr. 46
- ↑ a b c d e Richerson, J.R. & Boyd, R. (2005): Not by Genes Alone. How Culture Tranformed Human Evolution. University of Chicago Press
- ↑ Pigliucci, Massimo (2008) What, if anything, Is an Evolutionary Novelty? Philosophy of Science 75 (Dec. 2008) S. 898
- ↑ Müller, Gerd B. & Wagner, Günter P. (1991): Novelty in Evolution: Restructuring the Concept S.243
- ↑ Müller, G.B. Epigenetic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 312
- ↑ West-Eberhard, Mary Jane (2003): Development Plastizity and Evolution. Oxford University Press S. 198
- ↑ Pigliucci, Massimo (2008): What, if Anything, Is an Evolutionary Novelty? Philosophy of Science Ass. S. 890
- ↑ Liem, Karel F. (1973): Evolutionary Strategies and Morphological Innovations: Cichlid Pharyngeal Jaws, Systematic Zoology, Vol. 22, No. 4. (Dec., 1973), pp. 425-441
- ↑ Galis, F., & Drucker, E.G. (1996): Pharyngeal Biting Mechanics in Centrarchid and Cichlid Fishes: Insights into a Key Evolutionary innovation. Journal of Evolutionary Biology 9 (1996) pp. 641–670. A detailed study of a key innovation in fish evolution
- ↑ Müller, G. B.: Epigenetic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 314ff
- ↑ Newman, Stuart A.: Dynamic Patterning Modules. in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 281ff
- ↑ Müller, G. & Newman S. the Innovation Triad. An Evo-Devo Agenda. in Journal of Experimental Zoology 304B: 387-503
- ↑ Müller, Gerd B. Epigenetic Innovation in Pigliucci, M. & Müller G.B. (Hg) (2010): Evolution The Extended Synthesis, MIT Press, S. 313
- ↑ Müller, G. B.: Epigenetic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 314ff
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005): The Innovation Triad. An EvoDevo Agenda in Journal of Experimental Zoology 304B S. 491
- ↑ West-Eberhard, M.J. (2003): Development Plastizity and Evolution. Oxford University Press S. 500ff
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 492
- ↑ West-Eberhard, M.J. (2003): Development Plastizity and Evolution. Oxford University Press S. 501
- ↑ Müller, G.B.: Epiegentic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Ed.) (2010): Ecolution - The Extended Synthesis, MIT Press S. 307
- ↑ Reichholf, Josef H.: Die Erfindung der Feder S.99ff in: Sentker, Andreas & Wigger, Frank (Hg.) (2008): Triebkraft Evolution - Vielfalt, Wandel und Menschwerdung
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 492
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) S. 493
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 494
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 494
- ↑ West-Eberhard, M.J. (2003): Development Plastizity and Evolution. Oxford University Press S. 157
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 494
- ↑ Müller, G. B.: Epigenetic Innovation in Pigliucci, Massimo & Müller, Gerd B. (Hg.) (2010): Evolution - The Extended Synthesis, MIT Press S. 307-332
- ↑ Müller, G. B.: Epigenetic Innovation in Pigliucci, Massimo u. Müller, Gerd B. (Hg.) (2010): Evolution – The Extended Synthesis, MIT Press, S. 323
- ↑ Waddington, Conrad Hal (1942): Canalisation of development and the inheritance of acquired characters. In: Nature. Band 150, 1942, S. 563-56
- ↑ Müller, Gerd B. & Newman, Stuart A. (2005) a.a.O. S. 493
- ↑ Müller, Gerd B. & Newman, Stuart A. (2005): a.a.O. S. 494 mit Bez. auf Averof & Cohen 1997 u. Marden & Kramer 1994
- ↑ Gould, Stephen J. (1980, dt. 1987): Der Daumen des Panda – Betrachtungen zur Naturgeschichte. Suhrkamp TB Wissenschaft
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) a.a.O. S. 497
- ↑ Vollmer, Gerhard: Wieso können wir die Welt erkennen? in Fischer, Peter & Wiegandt, Klaus (Hg.) (2003): Evolution. Geschichte und Zukunft des Lebens. Fischer Taschenbuch S. 286.
- ↑ Lewontin, Richard & Gould, Steven Jay (1979): The spandrels of San Marco and the Panglosssian paradigm: a critique of the adaptionist programme
- ↑ Richerson, P.J. & Boyd R. (2005): Not by genes alone. How culture transformed humane evolution. Univserity of Chicago Press S. 137
- ↑ Jablonka, Eva & Avital, Eytan (2006): Animal Innovation: The Origins and Effects of New Learned Behaviours. Biology and Philosophy (2006) 21, S. 135; book review of: Animal Innovation (2003): Reader, S.M. & Laland, K.N. (eds.) (2003), Oxford University Press
- ↑ Reader, S.M. & Laland, K.N. (eds.) (2003): Reader, S.M. & Laland, K.N. (eds.) (2003): Animal Innovation (2003): Oxford University Press, zit. n: Jablonka, Eva & Avital, Eytan (2006) a.a.O. S. 136
- ↑ Jablonka, Eva & Avital, Eytan (2006): a.a O. S. 135
- ↑ Jablonka, Eva & Avital, Eytan (2006): a.a O. S. 138 mit Bezug auf Raeder/Laland Kap.2-4
- ↑ Jablonka, Eva & Avital, Eytan (2006): a.a O. S. 137
- ↑ Jablonka, Eva & Avital, Eytan (2006): a.a O. S. 139
- ↑ a b Jablonka, Eva & Lamb, Marion (2006): Evolution in Four Dimensions. Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life, MIT-Press, S. 169
- ↑ Jablonka, Eva & Lamb, Marion (2006): Evolution in Four Dimensions. Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life, MIT-Press Kap. 5: The Behavioral Inheritance System, S. 155-191
- ↑ Tomasello, Michael (2007): The Chimpanzee Culture
- ↑ zu diesem Kap. siehe auch: O'Brien, Michael J. & Shennan, Stephen J. (Hg) (2009): Innovation in Cultural Systems: Contributions from Evolutionary Anthropology (Vienna Series in Theoretical Biology), MIT Press
- ↑ Eibl, Karl (2009): Kultur als Zwischenwelt. Eine evolutionsbiologische Perspektive. Suhrkamp. S. 99
- ↑ Singer, Wolf (2003): Die Evolution der Kultur. In Fischer, Ernst Peter & Wiegandt Klaus (2003): Evolution. Geschichte und Zukunft des Lebens. Fischer TB S. 301
- ↑ Singer, Wolf (2003): Die Evolution der Kultur. a.a.O. S. 303
- ↑ a b Singer, Wolf (2003): Die Evolution der Kultur. a.a.O. S. 304
- ↑ Singer, Wolf (2003): Die Evolution der Kultur. a.a.O. 305
- ↑ Singer, Wolf (2003): Die Evolution der Kultur. a.a.O. S. 305
- ↑ Eibl, Karl (2009): Kultur als Zwischenwelt. Eine evolutionsbiologische Perspektive. Suhrkamp. S.38
- ↑ Richerson, J.R. & Boyd, R. (2005): Not by Genes Alone. How Culture Tranformed Human Evolution. University of Chicago Press. S. 109





Wikimedia Foundation.