- Evolutionäre Entwicklungsbiologie
-
Die evolutionäre Entwicklungsbiologie oder kurz Evo-Devo (abgeleitet vom englischen Begriff evolutionary developmental biology) ist eine Forschungsrichtung der Biologie, die untersucht, wie die Systeme der Individualentwicklung (Ontogenese) der Lebewesen evolvieren und wie die Prozesse der Embryonalentwicklung die Evolution der Organismen beeinflussen.
Die evolutionäre Entwicklungsbiologie, entstanden in den 1980er Jahren, stellt die genetischen und epigenetischen Erkenntnisse der Entwicklungsbiologie in einen evolutionären Kontext. Dazu bedient sich Evo-Devo entwicklungsbiologischer und molekularbiologischer Labormethoden, um zu zeigen, welche Faktoren und Steuerungsmechanismen für die Ausbildung und den evolutionären Wandel von Geweben, Organen und morphologischen Strukturen verantwortlich sind und versucht, das Auftreten solcher Veränderungen im Verlauf der Stammesgeschichte der Organismen zu rekonstruieren. Auf der theoretischen Ebene integriert Evo-Devo die Konzepte der Entwicklungsbiologie in die Evolutionstheorie. Eine wesentliche Erkenntnisquelle ist dabei die Entschlüsselung der genetischen Basis für zahlreiche bis in die 1980er Jahre vollkommen rätselhafte Entwicklungsvorgänge, welche mit der Entdeckung der sog. Hox-Gene begonnen hatte.
Die Bedeutung der Entwicklung für die Evolution
Zum Zeitpunkt der Befruchtung ist die phänotypische Form eines Individuums noch nicht gegeben; das Genom (DNA) wird nur durch Vorgänge während der Entwicklung in die Form (den Phänotyp) „übersetzt“. Der Embryo muss also seine Form erst aus einer einzelnen, undifferenzierten Zelle in der individuellen Entwicklung (der Ontogenese) erzeugen. Weil jede Veränderung des Phänotyps nur über die Veränderung der embryonalen Entwicklung erfolgen kann und diese ihre eigenen Gesetzmäßigkeiten besitzt, ist für die Vertreter von Evo-Devo die Entwicklung ein zentraler Schlüssel für das kausale Verständnis der organismischen Evolution.
Aus Sicht zahlreicher Vertreter dieser Forschungsrichtung kann die embryonale Entwicklung dabei spontane Variationen und Innovation hervorbringen. Die Selektion wirkt erst anschließend auf diese Variationsformen und wählt die geeignetsten Individuen aus. Morphologische Form und komplexe Strukturen (Körperbaupläne) entstehen nach dieser Sicht vor allem durch systemimmanente, sich selbst regulierende Umbauten des Organismus während der embryonalen Entwicklung.[1][2][3]
Geschichte der Erforschung evolutionärer Veränderungen in der Ontogenese
Darwin und das 19. Jahrhundert
Dass die Embryonalentwicklung Relevanz für die Evolution hat, war bereits Charles Darwin bewusst.[4] Zu seiner Zeit war es aber weder möglich, die Entwicklungsprozesse genauer zu untersuchen, noch waren der genaue Mechanismus der Vererbung oder Gene und DNA bekannt. Darwin konzentrierte sich daher auf die natürliche Selektion als den primären Evolutionsfaktor.
Aus dem 19. Jahrhundert sind neben Darwin einige Embryologen zu nennen, die ebenfalls evolutionäre Gesichtspunkte behandelten:
- Karl Ernst von Baer stellte an Wirbeltieren fest, dass Embryonen verschiedener Arten um so schwieriger unterscheidbar sind, je früher in ihrer Entwicklung sie angetroffen werden (Baer-Regel)
- Fritz Müller kombinierte in seinem Buch Für Darwin (1864) natürliche Selektion und Embryologie und demonstrierte an Entwicklungsphasen von Krebstieren, dass ihre Stammesgeschichte ohne Darwins Theorie nicht erklärt werden könne. Seine Studien inspirierten wie jene von Baers
- Ernst Haeckel. Auf ihn geht die heute nicht mehr gebräuchliche biogenetische Grundregel zurück, die in Schärfung eines damals weit verbreiteten Rekapitulationsgedankens angibt, dass die beobachteten Parallelen zwischen Ontogenese und Phylogenese der Organismen auf der embryonalen Wiederholung von Merkmalen beruht, die in der Stammesgeschichte der Arten schon im Erwachsenen-Stadium ausgebildet waren.
- Wilhelm Roux war Schüler von Haeckel und Begründer der Entwicklungsmechanik. Er war bereits der Ansicht, dass kein fertiger Bauplan vererbt wird, sondern dass „den einzelnen Zellen ein gewisser Spielraum bleibt, innerhalb dessen sich das Geschehen gegenseitig selbst reguliert“ (1881).
Synthetische Evolutionstheorie seit 1930
In den letzten Jahrzehnten des 19. und den ersten des 20. Jahrhunderts wurden Darwins Erkenntnisse in die neue Fachrichtung der vergleichenden Embryologie aufgenommen. Entdeckungen wie diejenige der Keimblätter (Endo-, Meso-, Ektoderm) waren wesentlich zur Entschlüsselung der Homologie der Körperbaupläne. Nach den großen Entdeckungen der Anfangszeit erlahmte der Elan aber dadurch, dass zwischen renommierten Forschern Meinungsverschiedenheiten über die Bedeutung zahlreicher Einzelheiten ausbrachen, die mit den damaligen Methoden nicht entscheidbar waren. Das Hauptinteresse der Forschung wandte sich daraufhin neuen Disziplinen wie der Entwicklungsmechanik und der Genetik zu[5][6]
Auch während des Entstehens der Synthetischen Evolutionstheorie in den 1930er und 1940er Jahren gab es vereinzelt Wissenschaftler, die sich um eine stärkere Thematisierung der Entwicklung bemühten (z. B. Richard Goldschmidt, Conrad Hal Waddington, Iwan Iwanowitsch Schmalhausen). Die Synthetische Evolutionstheorie war jedoch mit dem dominierenden Fundament der Populationsgenetik (Ronald A. Fisher, Sewall Wright, J.B.S. Haldane) unter der Mithilfe anderer Disziplinen (u.a. Zoologie, Systematik: Ernst Mayr) stark auf statistisch-deskriptives Denken ausgerichtet, so dass Prinzipien der Individualentwicklung keine Aufnahme in den Kanon fanden. Thomas Hunt Morgan, einer der frühen Vertreter der Synthetischen Evolutionstheorie, selbst auch Embryologe, stellte 1932 die Behauptung auf, die Genetik sei der einzige wissenschaftlich gültige Ansatz für das Studium der Evolution.[7] Vorstellungen, die sich mit der direkten Wirkung von Umwelteinflüssen auf den entstehenden Organismus und seine evolutionäre Änderung befassten (Epigenetik), wurden nicht weiter verfolgt, weil diese dem neodarwinistischen Dogma widersprachen, wonach kein Informationsfluss möglich ist, der von außen auf die DNA wirkt und sie vererbbar verändert (Weismann-Barriere). Vor diesem Hintergrund kann verstanden werden, dass ein Forscher wie Conrad Hal Waddington, der 1942 eine Umweltinduzierung von Entwicklungsveränderungen und die Kanalisierung von Entwicklungsprozessen theoretisch beschrieb und deswegen als ein wichtiger Vorläufer der Evolutionären Entwicklungsbiologie gilt, seitens der Synthetischen Evolutionstheorie nicht beachtet wurde. Waddingtons Thesen gelangten erst in den vergangenen zwei Jahrzehnten zu einer Renaissance.[8][9]
Die Synthetische Evolutionstheorie, das Standardmodell der heutigen, auf Darwin zurückgehenden Evolutionstheorie, sieht die Abfolge von zufälliger Variation, natürlicher Selektion und resultierender Adaption von Populationen als ausreichend an, um die Entstehung organismischer Vielfalt zu erklären.[10] Ihre Vertreter vorrangig der 1930er bis 1950er Jahre nahmen dabei im Vergleich zu Darwin zum Teil restriktiver Einschränkungen vor, die sich aus der gerade neu entdeckten Genetik ergaben. Wichtig sind hier folgenden Grundannahmen der Synthetischen Theorie:
- Gradualismus ist die Annahme, dass sich evolutionäre Entwicklungen stets in kontinuierlichen, kleinen Veränderungen vollziehen, die sich zu größeren summieren.[11] (das Konzept des Gradualismus geht dabei ursprünglich auf Charles Darwin selbst zurück).
- Die Weismann-Barriere. Demnach besteht keine Möglichkeit einer vererbbaren Beeinflussung des Genoms bzw. der Keimzellen durch individuelle Erfahrung der Organismen.
- Die Ansicht der deterministischen Genotyp-Phänotyp-Beziehung besagt, Gene und phänotypische Merkmale stünden in einem festen Verhältnis zueinander, wobei der Genotyp den Phänotyp definiere.
Forscher wie Mary Jane West-Eberhard, Marc Kirschner, Gerd B. Müller und andere versuchen gegenwärtig diese Annahmen zu modifizieren und weiter zu entwickeln.[12][13]
Constraints und Heterochronie
Seit den achtziger Jahren mehrten sich die Stimmen, die für eine stärkere Beachtung der Entwicklung für die Evolution plädierten (Stephen Jay Gould u.a.): Man untersuchte verstärkt die entwicklungsbiologischen Beschränkungen, die das Spektrum der evolutionären Variation begrenzen (Constraints). Andere Forscher beschäftigten sich mit den zeitlichen Verschiebungen der modularen Komponenten im Entwicklungsablauf (Heterochronie), wodurch z.B. Verschiebungen in den Größenverhältnissen zwischen verschiedenen Organen erklärbar wären. Gavin de Beer hatte bereits 1954 die These aufgestellt, dass Änderungen im Timing von Entwicklungsereignissen die Variation von Merkmalen verursachen können, etwa längere oder kürzere Beinen oder die Ausbildung oder Nichtausbildung eines Schwanzes auslösen können.[14]
Entdeckung der Master-Kontrollgene und Erforschung der Genregulationen
Nach Scott F. Gilbert kann 1977 als Konzeptionsjahr der neuen Forschungsrichtung Evo-Devo angesehen werden, begründet durch das Erscheinen dreier bedeutender Publikationen in diesem Jahr[15]: Stephen J. Goulds Ontogeny and Phylogeny, Francois Jacobs Evolution by Thinkering sowie eine technische Arbeit von A. Maxam und Walter Gilbert zu DNA-Sequenzierung. 1982/83 entdeckte man wichtige Master-Kontrollgene[16], die an der Regulierung grundlegender Körperbaupläne beteiligt sind, darunter die Hox-Gene, die für die Spezifikation der Körperlängsachse (in entwicklungsbiologischer Terminologie: der Anterior-Posterior-Körperachse) hauptverantwortlich sind, später die Pax-Gene mit grundlegender Bedeutung für die Augenentwicklung sowie die Mkx-Gene, die an der Herz-Formation beteiligt sind. Es stellte sich heraus, dass die Gruppe der Hox-Gene „in abgewandelter Form in bisher allen untersuchten vielzelligen Tieren vorkommt“,[17] sie sind homolog und müssen daher über einen sehr großen Zeitraum in der Evolution konserviert sein; mindestens seit der „kambrischen Explosion“ vor 530 Millionen Jahren, Paul Layer spricht sogar von rund einer Milliarde Jahre.[18] Die Entdeckung der Hox-Gene und ihrer Homologie für die Tierstämme zählt zu den herausragenden Entdeckungen der modernen Biologie der letzten Jahrzehnte. (siehe hierzu auch Homöobox).
In der Folge ermöglichte eine immer einfachere, schnellere und kostengünstigere Sequenzierung von Genomen und der vergleichenden Genetik einen verbesserten Einblick in die Genregulationsprozesse während der Entwicklung. Dies hatte zur Folge, dass sich jene Thematik zu einem der stärksten Forschungsfelder von Evo-Devo entwickelte.[19]
Epigenetische Mechanismen
Gemäß der Synthetischen Theorie ist Evolution als Veränderung der Allelfrequenz (Genfrequenz) in Populationen definiert.[20] Damit wird jedoch nicht erklärt, wie Variation und Innovation entstehen. Wurde die embryonale Entwicklung von den Hauptvertretern der Synthetischen Theorie noch als nicht relevant für die Evolution angesehen, steht sie heute im Fokus der kausalen Evolutionsforschung. Während die gängige Theorie auf das survival of the fittest fokussiert ist, versucht die evolutionäre Entwicklungsbiologie das arrival of the fittest zu erklären, also die Frage: wie entstehen die Geeignetsten?[21] Evo-Devo erweitert also den Forschungshorizont der Evolutionstheorie um neue Fragen und neue Erklärungen. Die von Evo-Devo neu beschriebenen epigenetischen Mechanismen der Reaktionsfähigkeiten der Entwicklung sind u.a.:
- Kanalisierung (siehe Kapitel 3, Kapitel 6 und 9.1)
- Heterochronie (siehe Kapitel 3)
- konservierte zelluläre Kernprozesse, die adaptiv kombiniert werden können (siehe Kapitel 5 ):
- schwache regulatorische Kopplungen bei Zell-Zell-Kommunikation
- explorative Prozesse / Verhalten
- Kompartimentierung
- Reaktionsfähigkeit der Entwicklung auf initiierende Umweltstressoren (siehe Kapitel 3 bis Kapitel 6 und Kapitel 8)
- nicht-lineare Effekte der Entwicklung mit diskontinuierlichem, phänotypischem Output (siehe Kapitel 4, Kapitel 6 und Kapitel 8).
Solche Mechanismen stellen für die Evo-Devo-Forschung neben Selektion und den genetischen Faktoren von Mutation, Rekombination und Gendrift eigenständige Evolutionsfaktoren dar, die es erlauben, das Entstehen von Variation und Innovation auch mechanistisch zu erklären.
Die Evo-Devo-Forschungsthemen
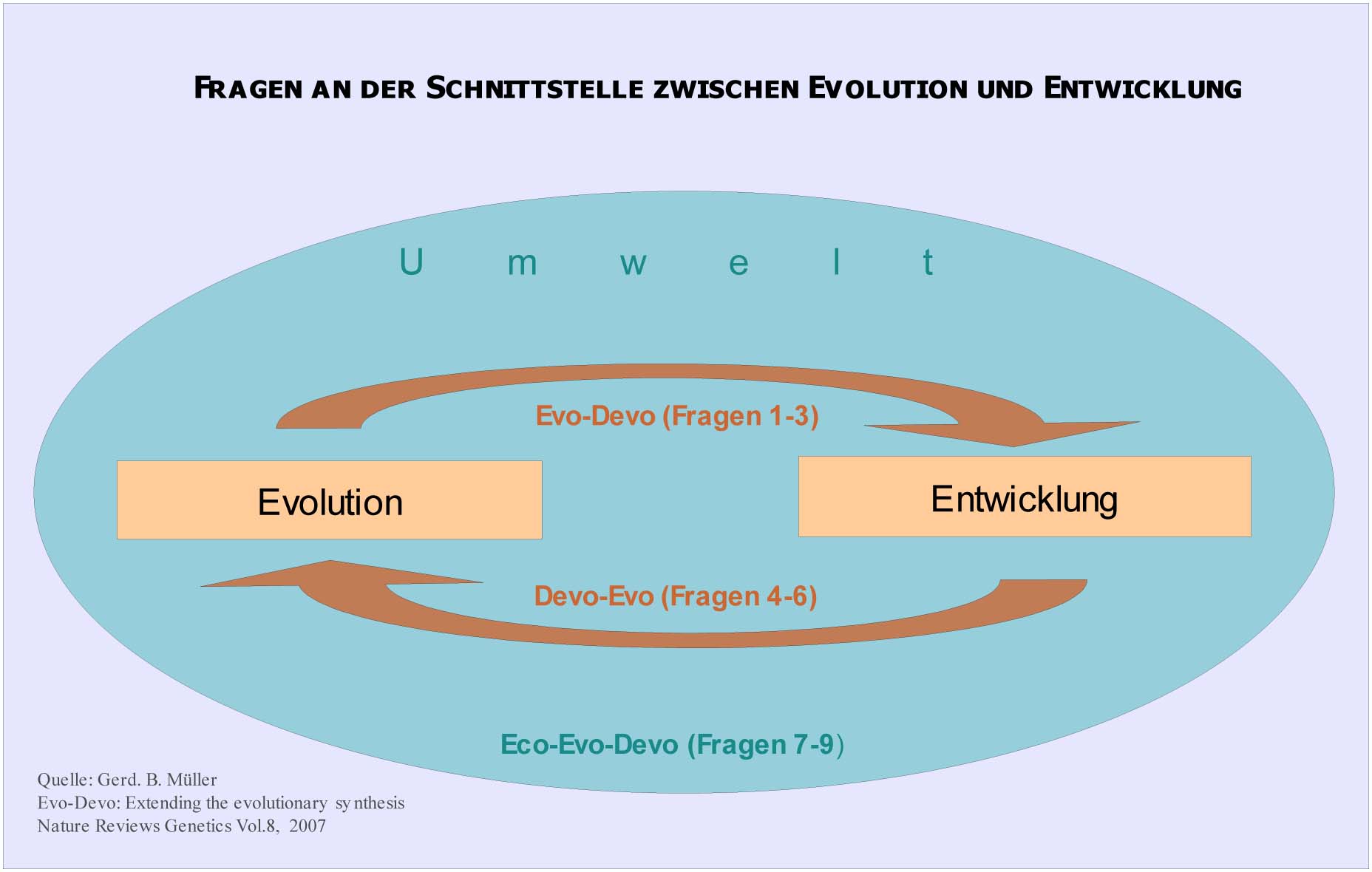
Die Evolutionäre Entwicklungsbiologie betrachtet das gesamte System der embryonalen Entwicklung selbst als ein evolviertes System und sieht es in einem komplexen systemischen Zusammenhang mit der Umwelt. Müller teilt das Forschungsgebiet in folgende drei Themenblöcke ein,[22] von deren Einzelfragen manche erst am Beginn wissenschaftlicher Bearbeitung stehen (s. Abb.2):
Zunächst als erster Block die Evo-Devo-Fragen, die sich von der Evolution auf die Entwicklung richten:
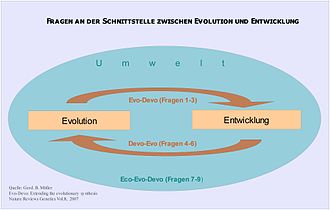 Abb.2 Evo-Devo fragt auch nach Wirkungsmechanismen der Entwicklung auf die Evolution und Interaktion zwischen Entwicklung, Evolution und Umwelt. Mit dieser Betrachtung wird die Evolutionstheorie methodologisch komplex.
Abb.2 Evo-Devo fragt auch nach Wirkungsmechanismen der Entwicklung auf die Evolution und Interaktion zwischen Entwicklung, Evolution und Umwelt. Mit dieser Betrachtung wird die Evolutionstheorie methodologisch komplex.
1. Der erste Themenkreis beschäftigt sich damit, wie Entwicklung bei rezenten Arten in der Evolution entstehen konnte. Was wir heute in höher entwickelten Arten in der Embryonalphase sehen und analysieren, dieser in seinem Zusammenspiel mit der Außenwelt noch nicht ganz verstandene Prozess, war nicht immer in diesem (wie Müller es nennt) routinierten, fein justierten Wechselspiel vorhanden. Es muss einen langen Evolutionsprozess bis heute gegeben haben. Ausgehend von ersten Metazoen sind erst viel später die selektive Fixierung und genetische Routinierung in den robusten Formen der Entwicklung und in den zuverlässigen mendelschen Vererbungsformen aufgegangen, wie wir sie in heute existierenden Organismen beobachten (Gerd B. Müller).
2. Unter der Evolution des Entwicklungsrepertoires versteht man zum einen die genetischen Werkzeuge. Carroll[23] spricht vom genetischen Werkzeugkasten (genetic toolkit). Man fragt und erforscht, wie dieser entstehen und evolvieren konnte oder wie zum Beispiel genetische Redundanz, neue Genfunktionen, Modularität auf Genomebene entstehen konnten. Zum andern gehört zum Entwicklungsrepertoire aber auch eine komplexe Vielfalt epigenetischer Prozesse (s. Abb.5: Das integrierte evolutionäre Entwicklungssystem). Diese Prozesse waren vor hunderten Millionen Jahren einfacher. Heute enthalten sie ausgefeilte, eingespielte Mechanismen, die zum Beispiel Zellwechselwirkungen genau regulieren. Das Entwicklungsrepertoire selbst entstand also durch Millionen Jahre Evolution. Es selbst vermehrt sich laut Müller in der Evolution.
3. Die Frage: Wie wirkt Evolution auf spezielle Entwicklungsprozesse ein? Hier gibt es z. B. die Heterochronie, die zeitliche Verschiebung von Entwicklungsprozessen. „Evolutionäre Modifizierungen in der Segmentierung und regionalen Differenzierung größerer Körperbausektionen wird begleitet von Verschiebungen in Domänen der Hox-Gen-Expressionen“[24]
Der zweite Block betrifft DevoEvo-Fragen, die sich von der Entwicklung auf die Evolution richten, sozusagen die Gegenfragen zum ersten Block. Diese Fragen sind das spezifisch Neue in Evo-Devo, die die kausalen Wechselwirkungen zwischen Entwicklung und Evolution erst sichtbar machen und die die herrschende Evolutionstheorie verändern.
4. Wie beeinflusst Entwicklung phänotypische Variation? Um zu unterbinden, dass in der Entwicklung zu große, unerwünschte evolutionäre Variationen auftreten, haben sich Entwicklungsconstraints gebildet. Solche Constraints sind physikalischer, morphologischer und phylogenetischer Natur. Sie führen zu einer Kanalisierung der Entwicklung (Conrad Hal Waddington), zu Robustheit. Man spricht auch von der development reaction norm, eine Bandbreite innerhalb derer sich phänotypische Plastizität vollziehen kann.
5. Was trägt Entwicklung zu phänotypischer Innovation bei? Wenn die Selektion allein gesehen keine Form bilden kann, muss es einen anderen Weg geben, wie organismische Innovation entsteht. Die Antwort kann für Evo-Devo nur in der Entwicklung liegen.
6. Wie wirkt Entwicklung auf die Organisation des Phänotyps? Die Frage nach der Organisation der Körperbaupläne in der Entwicklung ist nicht auf die Entstehung oder Variation bestimmter Körpermerkmale gerichtet, sondern darauf, wie der Organismus als ein integriertes System hergestellt werden kann.
Schließlich der dritte Block, das ist der Eco-Evo-Devo-Fragenkreis, der die kausale Beziehung von Entwicklung und Evolution mit der Umwelt betrifft, ebenfalls neu von Evo-Devo eingebracht, da die Synthetische Evolutionstheorie keine derartigen Wirkungsmechanismen erklären kann.
7. Wie interagiert die Umwelt mit Entwicklungsprozessen?
8. Wie beeinflussen Umweltänderungen die phänotypische Evolution?
9. Wie wirkt die evolutionäre Entwicklung auf die Umwelt?
Ein zentrales Konzept dieses Fragenkreises ist die phänotypische Plastizität. Plastizität bedeutet, dass ein Genotyp unter verschiedenen Umweltbedingungen verschiedene, unter Umständen stark abweichende, Phänotypen erzeugen kann. Ein bekanntes Beispiel ist, dass aus Eiern bestimmter Schildkrötenarten je nach Temperatur und damit umweltabhängig, männliche oder weibliche Nachkommen schlüpfen.[25] Schmetterlinge erzeugen abhängig von der Jahreszeit unterschiedliche Flügelfärbung. Mehr Tageslicht und geringere Temperatur bringen einen dunklen Typ hervor, weniger Licht einen orangen Typ.[26]
Die Theorie der erleichterten Variation
Die Entschlüsselung der genetischen Basis der Entwicklungsvorgänge beim Wachstum hat gezeigt, dass die wesentlichen Prozesse in Entwicklungs-Modulen organisiert sind. Dutzende bis hunderte genetisch codierte Strukturen und Struktureinheiten werden über gemeinsame regulatorische Einheiten synchron gesteuert. „Master-Kontrollgene“ an Schlüsselstellen können die Entwicklung ganzer Organe induzieren, z.B. kann das pax6-Gen überall die Entwicklung funktionsfähiger Augen induzieren. Die auslösenden regulatorischen Einheiten, meist zelluläre Signalwege und (über Transkriptionsfaktoren gesteuerte) cis-regulatorische Abschnitte im Genom abseits der proteinkodierenden Sequenzen, steuern dabei die Entwicklung keinesfalls bis ins Detail, sondern bilden gleichsam Schalter, die in sich koordinierte Entwicklungspfade ein- oder ausschalten können. Die genetische Basis des Steuerungswegs ist also unterschiedlich zu derjenigen der damit gesteuerten Struktur selbst. Dies bedeutet, dass sie unabhängig davon variieren und selektiert werden kann. Sean Carroll prägte dafür das Bild des „genetischen Werkzeugkastens“.
Andere Prozesse werden gar nicht bis ins letzte Detail genetisch festgelegt. Das Entwicklungsprogamm stellt hier lediglich eine noch weitgehend gestaltlose Grundstruktur bereit, die dann erst durch Einflüsse der Umwelt im Detail gestaltet wird: beispielsweise die Reifung des Zentralnervensystems, bei dem von den zahllosen angelegten synaptischen Verbindungen zwischen den Nervenzellen die benötigten verstärkt werden und die nicht genutzten zugrunde gehen. Dadurch braucht die Detailarchitektur nicht genetisch vorgegeben zu werden.
Die Autoren Kirschner und Gerhart fassen die Auswirkungen dieser Erkenntnisse auf die Evolutionstheorie zusammen, sie sprechen sprechen von erleichterter Variation (facilitated variation).
Konservierte Kernprozesse
Die grundlegenden Strukturen der Zellorganisation und zahlreiche der dem Körperbauplan und seinen Organen zugrunde liegenden Strukturen werden demnach als konservierte Kernprozesse betrachtet. Sie dienen der Feinsteuerung durch die Entwicklungsmodule danach gleichsam als Rohmaterial. Die Einzelprozesse ändern sich dabei nicht. Zellverhaltensweisen können also evolutionär neu kombiniert werden oder in neuem Ausmaß eingesetzt werden. Wichtige Beispiele für solche konservierten Kernprozesse sind nach Kirschner und Gerhart:
- der einheitliche genetische Code aller Lebewesen
- die selektiv durchlässige Zellmembran zur Kommunikation zwischen Zellen sowie
- die identische Funktion der Hox-Gene.
Die stabilen Kernprozesse erlauben aus Sicht der evolutionären Entwicklungsbiologie Ausprägungsformen oder Eigenschaften, die eine erleichterte phänotypische Variation ermöglichen. Das sind nach Kirschner/Gerhart:
- explorative Prozesse,
- schwache regulatorische Kopplungen und
- Kompartimentbildung beim Embryo.
Exploratives Verhalten
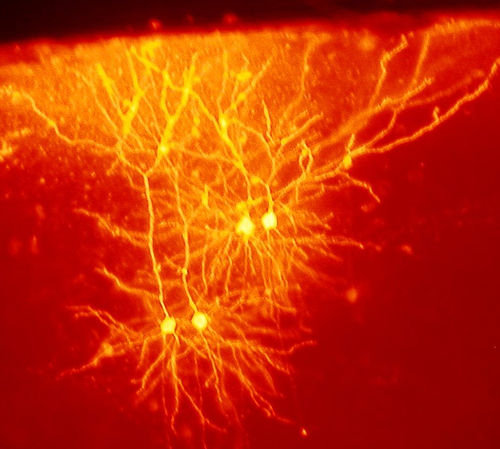
Die differenzierte Ausbildung etwa von Sehnen, Muskeln, Nerven und Blutgefäßen während der Entwicklung wird nicht im Detail vom Genom vorgegeben. Ihre Entstehungsweise kann als explorativ bezeichnet werden. Dabei zeigen Zellen je nach ihrer zellularen Umgebung alternative Reaktionen. So können Zellen „verzweigte Strukturen“[27] schaffen, die den gewünschten Geweberaum optimal ausfüllen[28] (z. B. Blutgefäß, Nervensystem). Explorative Strukturen sind im Verlauf der Entwicklung hochgradig anpassungsfähig.
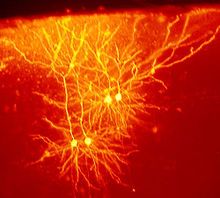 Abb.4 Exploratives Verhalten: Das Nervensystem (hier Maus-Cortex) ist nicht im Detail im Genom abgelegt. Axone und Dendriten „suchen und finden“ sich in der Entwicklung.
Abb.4 Exploratives Verhalten: Das Nervensystem (hier Maus-Cortex) ist nicht im Detail im Genom abgelegt. Axone und Dendriten „suchen und finden“ sich in der Entwicklung.Schwache regulatorische Kopplungen zwischen Zellen
Die für evolutionäre Variation notwendigen Neukombinationen der Kernprozesse sind durch Zellkommunikation vermittelt. Die Autoren sprechen hier von schwachen regulatorischen Kopplungen. „Schwach“, weil das Zellsignal die Entwicklung nicht im Detail steuert, sondern nur einschaltet, also eine nur schwache Beziehung zu den Spezifika des Outputs im Zielbereich hat. In der Regel bestimmt der Signalstoff an der Zieladresse das „An“ oder „Aus“ für die Expression eines dort vorhandenen Gens, dieser kann in derselben Zelle oder in einer anderen Zelle sein. Was dann jedoch genau geschieht, ist durch die eigene Regulation im Zielbereich festgelegt. Der zelluläre Mechanismus im Zielbereich ist schon früher entwickelt worden und braucht für die spezifische Reaktion nur aktiviert werden.[29] Ein Beispiel für schwache regulatorische Kopplungen sind etwa der durch Zellen mehrstufig gesteuerte Prozess der Insulinabgabe des Organismus nach Glucosezufuhr sowie viele andere Stoffwechselprozesse.
Kompartimentierung
Erst im Verlauf der Entwicklung kommt es zur Ausbildung differenzierter Zellen für spezifische Gewebetypen (Haut, Muskel, Nerven, Organe etc.). Es gibt also Regionen des Embryos, in denen in einer ganz bestimmten Phase der Entwicklung ein oder wenige ganz bestimmte Gene der Zellen exprimiert und bestimmte Signalproteine produziert werden.[30] Die Fähigkeit, unterschiedlich konservierte Kernprozesse an unterschiedlichen Orten im Organismus zu aktivieren und diese Reaktionsräume eigentlich erst zu schaffen, nennen sie Kompartimentierung. Ein Insektenembryo bildet in der mittleren Phase der Entwicklung ca. 200 Kompartimente aus. Kompartimentkarten dienen als Gerüst für Anordnung und Bau komplexer anatomischer Strukturen von Lebewesen. Jeder Tierstamm hat seine typische Karte.[31] Die Ausprägung dieser Kompartimente ist die eigentliche Aufgabe der hox-Gene.
Die Organismen, d.h. der Phänotyp, spielen demnach eine Hauptrolle bei der Festlegung von Natur und Maß der Variation.[32] Phänotypische Variation kann nicht beliebig sein. Vielmehr bedingt erleichterte Variation einen beeinflussten „vorsortierten“ Output phänotypischer Variation durch einen Organismus.[33] Variation wird vorwiegend deshalb erleichtert, weil so viel Neuheit in dem verfügbar ist, was Organismen bereits besitzen.[34]
Die Rolle epigenetischer Vererbung
Dass es Möglichkeiten gibt, wie phänotypische Merkmale abseits der DNA mit der Abfolge ihrer Basenpaare von einer Generation zur nächsten vererbt werden können, ist erst seit den 1980er Jahren nach und nach klar geworden. Im Gegensatz zur Genetik werden diese Prozesse Epigenetik genannt. Die Vertreter der synthetischen Evolutionstheorie haben sich mit solchen Prozessen nur widerstrebend befasst, weil sie für sie einen „lamarckistischen“ Beiklang hatten, d.h. an die gerade überwundenen Vorstellungen von der Vererbung erworbener Eigenschaften erinnerten. Ausmaß und Rolle epigenetischer Vererbung sind aktive Forschungsfelder[35][36] West-Eberhard hat in ihrem umfangreichen Werk über die phänotypische Evolution (2003) als eine der ersten zahlreiche Beispiele geliefert, die belegen, dass die Umwelt direkte Wirkung auf die Entwicklungsprozesse und auf diesem Weg auch auf die Evolution haben kann. Eine umfangreiche Theorie zur besonderen Rolle epigenetischen Vererbung stammt von Jablonka und Lamb.[37]
Bekannte Mechanismen epigenetischer Vererbung[38] umfassen:
- DNA-Methylierung. Werden Basen des DNA-Strangs enzymatisch methyliert, werden sie quasi „stumm geschaltet“. Dasdurch können einzelne Gene oder Teile des Genoms (z.B. das gesamte väterliche oder mütterliche Erbgut) von der Expression ausgenommen werden. Vererbung von Methylierungsmustern findet offensichtlich häufiger bei Pflanzen als bei Tieren statt (Pflanzen haben keine Keimbahn), spielt aber in beiden eine Rolle. Methylierungsmuster spielen sowohl bei der Steuerung der Entwicklung wie bei der Reaktion auf Stressfaktoren in der Umwelt[39] eine große Rolle.
- Regulatorische RNA. Teile des Genoms kodieren nicht für Proteine, sondern für (meist kurze) RNA-Sequenzen, die eine wichtige Rolle bei der Steuerung der Genexpression haben können.
- Histon-Komplexe. Histone sind DNA-begleitende Proteine, die bei der Stützung und „Verpackung“ eine Rolle spielen. In einzelnen Fällen wurde nachgewiesen, dass verschiedene Modi von DNA-Histonkomplexen existieren können. Besonders „dicht“ verpackte DNA wird seltener transkribiert.
Einige Autoren wollen ein weites Feld weiterer, nicht durch DNA vererbter Faktoren, bis hin zu menschlicher Kultur, als epigenetische Faktoren mit behandelt wissen.
Wichtige mögliche Auswirkungen epigenetisch vererbter Prozesse umfassen:
- Epigenetisch determinierte Merkmale sind labiler und variieren deutlich stärker. Durch epigenetische Varianten kann also die Reaktionsnorm einer Population auf Umweltstress erheblich erhöht sein. Möglicherweise liefert die epigenetische Variation eine zusätzliches Feld für kurzzeitige Variationsmöglichkeiten, das schneller reagieren kann als die Mutation der DNA.
- Epigenetisch gesteuerte Merkmalsausprägungen liefern einen Weg, über den sich die Merkmalsvariation innerhalb einer Population schnell und synchron in eine bestimmte Richtung verändern kann, wenn sich die Umweltbedingungen verändern.
- Es erscheint denkbar, dass innerhalb solcher epigenetisch geprägter Populationen eine Mutation auftritt, die den bisher modifikatorisch erzeugten Phänotyp als erbliche Variation fixiert. Dadurch können vorteilhafte Mutationen, die anfangs notwendigerweise sehr selten sind, einen entscheidenden Startvorteil erhalten und nicht ihr Auftreten als solches, aber ihre Fixierung in der Population stark gefördert werden. Dieser Vorgang wird als „genetische Assimilation“ bezeichnet[40] (vgl. dazu auch: Baldwin-Effekt).
Heute existieren bereits einige empirische Versuche, die eine genetische Assimilation bestätigen (Waddington 1953 mit Veränderung der Adern an Fliegenflügeln, Nihjout 2006 mit Farbvariation der Tabakschwärmer-Raupe). Durch die beschriebene Existenz von Umweltfaktoren können solche Konstruktionsänderungen zunächst angestoßen werden. Das System ist dann in der Lage zur Selbstorganisation, auf solche Einflüsse zu reagieren. Dennoch ist darauf hinzuweisen, dass der vorgeschlagene Mechanismus bisher noch weitgehend spekulativ ist.
Die Entwicklung erzeugt phänotypische Innovation
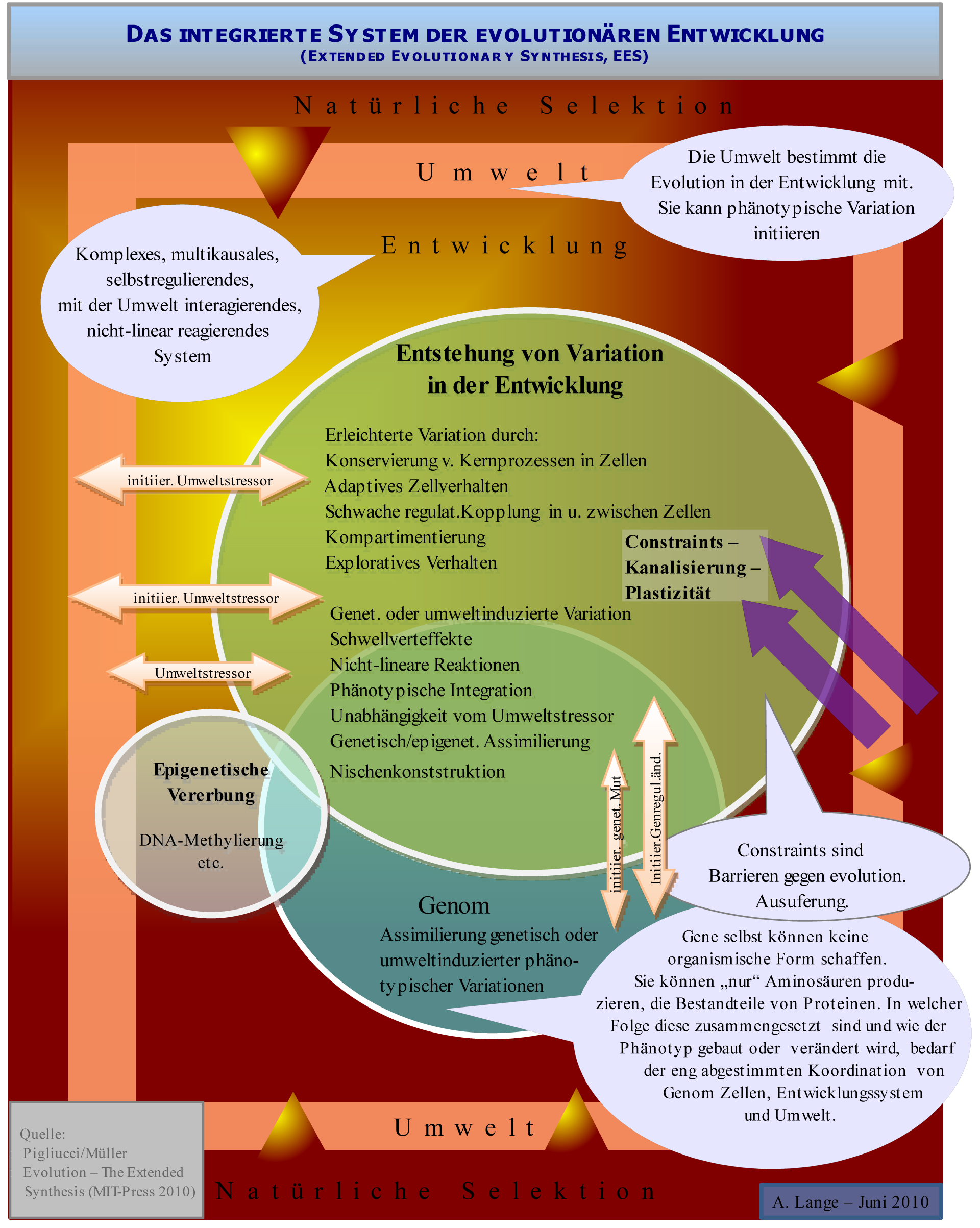
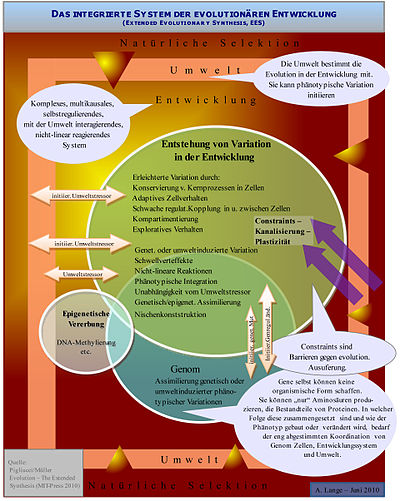 Abb.5 Das integrierte, evolutionäre Entwicklungssystem: Umwelt, natürliche Selektion, Genom und Entwicklung interagieren auf komplexe Weise zur Erzeugung phänotypischer Variation.
Abb.5 Das integrierte, evolutionäre Entwicklungssystem: Umwelt, natürliche Selektion, Genom und Entwicklung interagieren auf komplexe Weise zur Erzeugung phänotypischer Variation.Nach der klassischen synthetischen Evolutionstheorie sind phänotypische Innovationen, also völlig neue Strukturen im Körperbauplan, nicht prinzipiell von Variationen unterscheidbar. Wichtigster Mechanismus zur Erzeugung völlig neuer Strukturen ist danach die Funktionsänderung ursprünglich für einen anderen Zweck adaptiv entstandener Strukturen (sog. Präadaptionen, auch: Exaptationen), z.B. der ursprünglich als Isolation entstandenen Feder für den Flügel der Vögel. Einige Vertreter der evolutonären Entwicklungsbiologie halten zur Entstehung von Innovationen andere, zusätzliche Mechanismen für erforderlich[41][42] Müller definiert drei Typen von Innovationen.[43] Aus der Sicht von Evo-Devo ist besonders Novelty Typ-2 von Bedeutung: Eine phänotypische Innovation ist ein neues Konstruktionselement in einem Bauplan, das weder eine homologe Entsprechung in der Vorgängerart noch im selben Organismus hat. Es werden drei Phasen im Entstehen solcher evolutionärer Innovation unterschieden:[44][43]: Initiierung (meist durch veränderte Umweltbedingungen), Realisierung (durch die Steuerung im Entwicklungsprozess ermöglichte Umstellungen im Körperbauplan) und Akkkomodierung.
 Abb.6 Der Insektenflügel ist eine evolutionäre phänotypische Innovation (s.Flügel (Insekt)).
Abb.6 Der Insektenflügel ist eine evolutionäre phänotypische Innovation (s.Flügel (Insekt)).Innerhalb der Theorie nimmt man an, dass neue phänotypische Elemente zunächst epigenetisch fixiert und anschließend assimiliert werden. „Das Innovationsmerkmal muss in das bereits bestehende Konstruktions-, Entwicklungs- und Genom-System akkommodiert werden, um Funktionalität und Vererbung sicherzustellen“.[45] Dabei „scheint es die Regel zu sein, dass die epigenetische Integration der genetischen Integration vorausgeht“[46] oder wie West-Eberhard es ausdrückt: „Genes are followers in Evolution“.[47] „Die genetische Integration stabilisiert und überdeterminiert den generativen Prozess (Innovationsprozess) und resultiert in einem immer engeren Mapping zwischen Genotyp und Phänotyp“.[48]
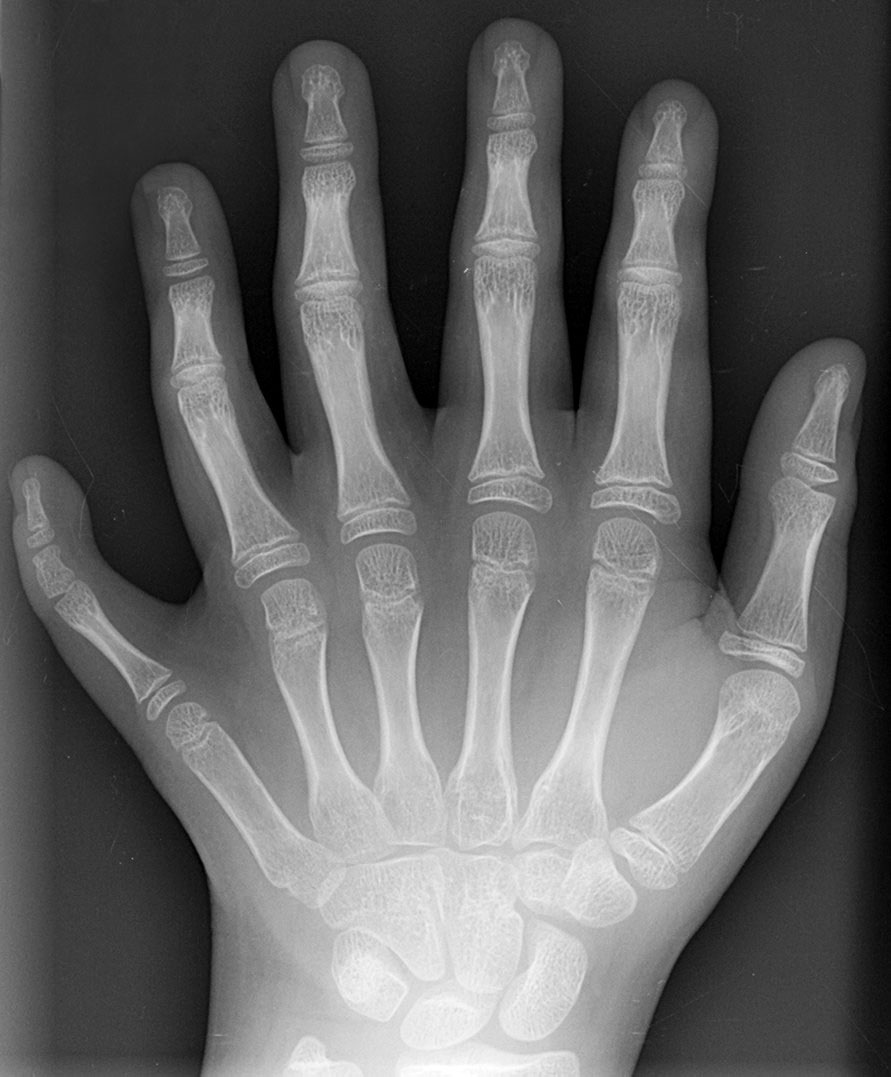
Kleine Ursachen (sowohl Umwelt wie Mutation) können phänotypisch starke Veränderung zur Folge haben. Für solche nicht-graduelle, diskontinuierliche Reaktionen sind die selbstorganisatorischen Eigenschaften des Entwicklungssystems verantwortlich. Ein Beispiel für eine spontane Konstruktion des Entwicklungssystems ist eine Hand mit sechs Fingern (Polydaktylie) (Abb.3).[49]
 Abb.3 Sechs Finger: Knochen, Muskeln, Blutgefäße und Größe des zusätzlichen Fingers können durch die Entwicklung vollständig in die Anatomie der Hand integriert sein.
Abb.3 Sechs Finger: Knochen, Muskeln, Blutgefäße und Größe des zusätzlichen Fingers können durch die Entwicklung vollständig in die Anatomie der Hand integriert sein.Das Beispiel belegt, dass die embryonale Entwicklung in der Lage ist, solche phänotypische Variation zu erzeugen und dass sechs Finger an einer Hand oder an beiden Händen eine mögliche evolutionäre Variation darstellen könnten. Die Entwicklung kann hierbei ein vollständig integriertes Merkmal erzeugen: Nerven, Muskeln, Gelenk, Skelett des Fingers und seine Größe sind vollständig funktionsfähig in die Anatomie der Hand integriert.
Verhältnis von genetischer und epigenetischer Dimension
Aufbauend auf fortschreitenden (vergleichenden) Genomsequenzierungen untersucht die evolutionäre Entwicklungsbiologie auch die umfangreichen Genregulationen während der Entwicklung. Sean B. Carroll[50] oder Wallace Arthur[51], aber auch Paul Layer[52] sehen entsprechend die Genregulationsprozesse mit wechselnden Kombinationen von Genschaltern als vorherrschende Einflussfaktoren auf die Entwicklung des Organismus sowie auf sein Veränderungspotenzial. Nach dieser Sicht sind Mutationen im regulatorischen Genom wesentlicher für die organismische Evolution als Mutation von Strukturgenen. Andere Forscher wie Marc Kirschner, Müller, Massimo Pigliucci oder West-Eberhard gehen weiter und betrachten den gesamten Entwicklungsapparat als komplexes System, das auf den verschiedenen genetischen und epigenetischen Ebenen (DNA, Zellkern, Zellen, Proteine, Zellkommunikation, Zellaggregate, Organismus, Umwelt) auf komplexe Weise mit den vorgestellten Mechanismen agieren und reagieren kann.[53]
Bildgebende Verfahren
Video 1: Maus Embryo (microCT) Theiler stage 21, stained with iodine (IKI)Evo-Devo bedient sich in der empirischen Forschung entwicklungsbiologischer und molekularbiologischer Labormethoden, um Faktoren und Steuerungsmechanismen für die Ausbildung und den evolutionären Wandel von Geweben, Organen und morphologischen Strukturen zu erkennen. Das Auftreten solcher Veränderungen wird im Verlauf der Stammesgeschichte der Organismen rekonstruiert. Einen wichtigen Beitrag für Evo-Devo haben bildgebende micro-CT-Verfahren, Computertomografie im Mikro- und Nanometerbereich.[54] Mit Kontrastmitteln lassen sich Genaktivitäten sichtbar machen, so dass der Beitrag eines Gens oder mehrerer beim raum-zeitlichen Entwicklungsvorgang beobachtbar wird.[55] Benötigt wird neben der Genexpressionsebene die „kalibrierte, dreidimensionale Darstellung anatomischer Strukturen in deren natürlichem Aussehen und räumlichen Beziehungen, so nahe am natürlichen Zustand wie für präparierte Specimen nur irgend möglich“ (Metscher).[56] Einen umfassenden Anspruch zur Kartierung des kompletten Embryonalverlaufs von Drosophila mit bildgebenden Verfahren hat das Projekt BDTNP (Barkeley Drosophila Transcription Network Project). Ziel ist dabei, vollständige Genexpressionsatlanten zu erstellen. Erzeugt wird ein Datensatz von 75.000 Bildern je Embryo mit sichtbar gemachten Aktivitäten von ca. 50% der Gene. Das „repräsentiert eine solide Beobachtungsgrundlage für „die Analyse der Beziehung zwischen Gensequenz, gewebespezifischer Genexpression und Entwicklung in der Tierwelt“ (Tomancak)[55]. Der komplette Atlas enthält die Daten aller Transkriptionsprodukte des Drosophila-Genoms in allen Phasen der Entwicklung. Das führt in Zukunft zur „automatisierten Erstellung und Speicherung der Expressionsmuster lebender Arten in vier Dimensionen"[55]. Das Projekt BDTNP zeigt beim heutigen Stand im Internet mit Videostreams die computergestützte statistische Auswertungsspezifischer Genexpressionen hunderter von Embryonalvergleichen von Drosophila (virtuelle Embryonen). Der Vergleich der Prozesse dient der Erzeugung statistischer Wahrscheinlichkeiten für das Entstehen phänotypischer Bandbreiten bestimmter Gewebe. Mit Stressoren (Hitze, Kälte, Ernährung etc.) lassen sich zukünftig Expressionsmuster verändern, statistisch auswerten und auf diese Weise mögliche evolutionäre Entwicklungspfade aufspüren.
Ausgewählte empirische Forschungsergebnisse
Belege bei der Taufliege
Erkenntnisse, dass Umweltfaktoren auf Vererbung und Evolution wirken, hatte bereits der Brite Conrad Hal Waddington (1942). Er konnte seine Theorie später (1953) anhand von Veränderungen an den Adern der Fruchtfliege (Drosophila melanogaster) empirisch belegen, indem er die Fliegeneier einige Generationen lang jeweils kurzen Hitzeschocks aussetzte. Dabei blieben nach einigen Generationen die Querverstrebungen an den Flügeln aus. Wurden die Hitzeschocks in Folgegenerationen als externer Stressor aufgehoben, blieben die durch sie induzierten Variationen am Flügeladersystem aber weiterhin erhalten, das heißt die Querverstrebungen erschienen nicht wieder. Die Hitzeschocks waren ein ausreichend starker Anstoß, dass bisherige Entwicklungspfade verlassen wurden. Bereits angelegte, aber bis dahin nicht genutzte, maskierte Pfade traten durch den äußeren Einfluss zu Tage. Die Entwicklung wurde mit Waddingtons Worten in einem neuen Pfad kanalisiert. Der Umweltfaktor war nur so lange erforderlich, bis der neu kanalisierte Entwicklungsverlauf, wie Waddington es ausdrückte, nachträglich auch genetisch assimiliert ist.[57] Hier bleibt er wieder so lange kanalisiert bzw. stabil, und zwar auch bei neuen auftretenden Mutationen, bis entweder eine Mutation oder aber neue Umwelteinflüsse stark genug sind, dass die Kanalisierung an ihre Grenzen stößt. Gegebenenfalls führt das dann unter Einwirkung von Schwellenwerteffekten, wie oben erläutert, zu einer neuen Variation des Phänotyps.
Was Waddington mit den Laborversuchen nicht zeigen konnte, ist, wie ein adaptiver Weg entsteht, dass auf einen Umweltfaktor (hier: Hitze) geeignet reagiert wird. Die Variation der Flügeladern ist kein adaptives Merkmal auf Hitzeeinwirkung. „Es ist keineswegs sicher, dass er mit nennenswerter Häufigkeit auf irgendeine besondere adaptive Morphologie gestoßen wäre“.[58]
 Abb. 7 Flügeladern bei Drosophila
Abb. 7 Flügeladern bei DrosophilaVeränderung der Schnäbel bei Darwin-Finkenarten
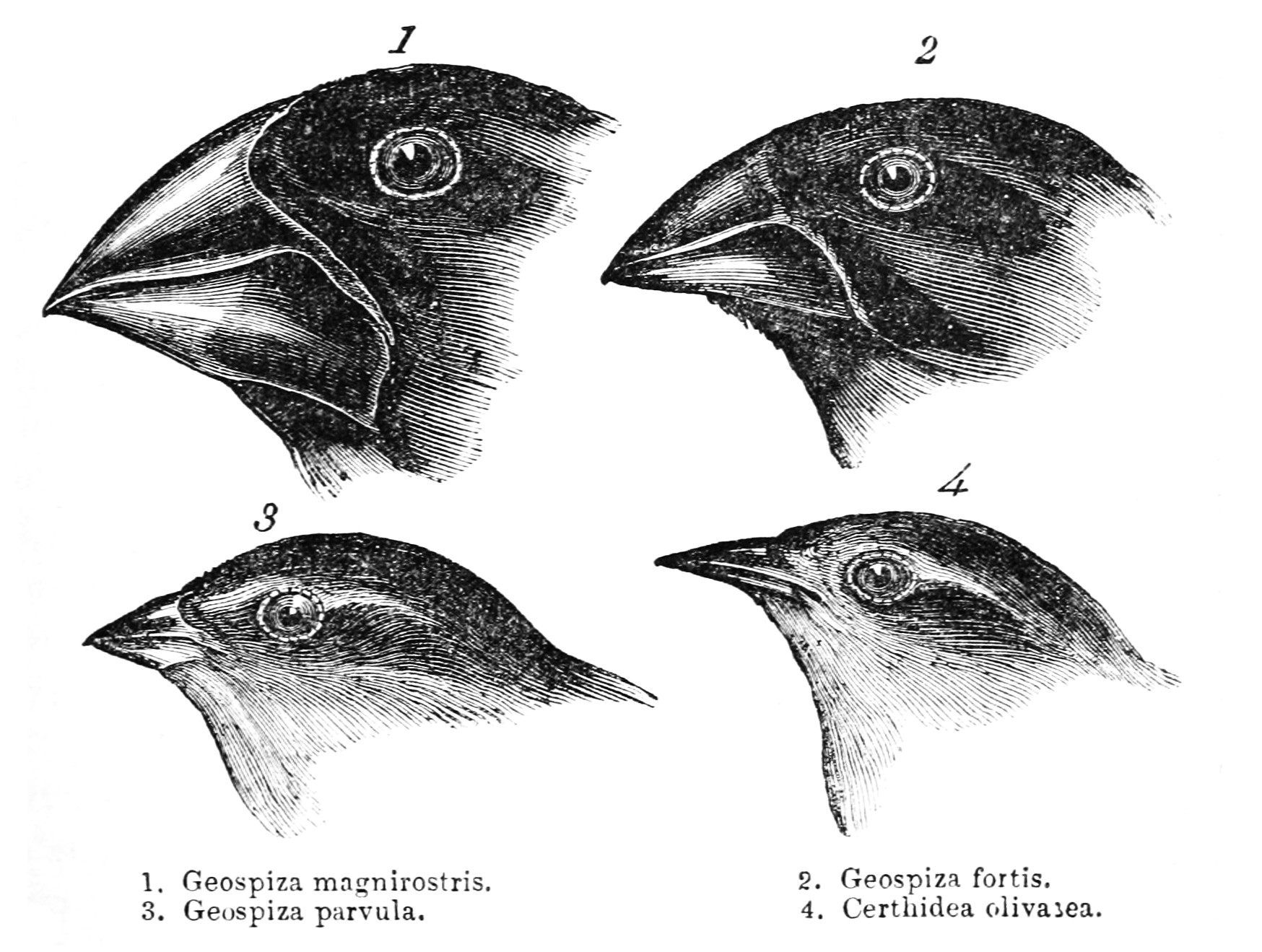
Sean B. Carroll konnte Veränderungen der Muster von Schmetterlingsflügeln im Labor vornehmen. Dabei gelang es durch Bestimmung der entsprechenden Genschalterkombinationen bei der Entwicklung des Schmetterlings, die Muster auf den Flügeln zu variieren.[59] 2007 haben Peter und Rosemary Grant[60] bei Darwin-Finkenarten auf den Galápagos-Inseln nachgewiesen, dass es in nur wenigen Generationen auf Grund von verändertem Nahrungsangebot (Initiator) zur Umbildung der Schnäbel kommt.[61]
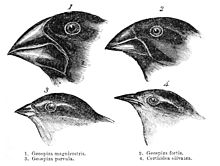 Abb.8 Evolutive Veränderung der Schnabelgröße und -form bei Darwinfinken. Eine Variation des Schnabels erfordert die vollständige Integration in die Anatomie des Kopfes. Das leistet die Entwicklung.
Abb.8 Evolutive Veränderung der Schnabelgröße und -form bei Darwinfinken. Eine Variation des Schnabels erfordert die vollständige Integration in die Anatomie des Kopfes. Das leistet die Entwicklung.In diesem Zusammenhang konnte man ein Wachstumsfaktor-Protein identifizieren, das an der Schnabelbildung im Embryo maßgeblich beteiligt ist, und konnte auch zeigen, dass dieses Protein bei verschiedenen Schnabelformen unterschiedlich stark oder unterschiedlich lange korreliert ausgebildet wird.[62] Kirschner/Gerhart erwähnen zudem, dass besagtes Protein (es heißt BMP4 und wird in Neuralleistenzellen produziert) experimentell in die Neuralleiste eines Hühnchens eingepflanzt wurde, wo sich ebenfalls die Schnabelform veränderte. Das Hühnchen entwickelte breitere und größere Schnäbel als normal. Andere Wachstumsfaktoren haben nicht diese Wirkung.[63] Obgleich also der experimentell manipulierte Schnabel seine Größe bzw. Form ändert, wird er dennoch in die Anatomie des Vogelkopfes integriert. „Es kommt nicht zu einer monströsen Fehlentwicklung“ (Kirschner/Gerhart).
Die Schnabelbildung ist ein komplexer Entwicklungsprozess, an dem fünf Nester von Neuralzellen beteiligt sind. Die Nester empfangen Signale von Gesichtszellen an den fünf Orten und reagieren auf sie. Daher beeinflussen Merkmale, die die Neuralleistenzellen beeinflussen, das Schnabelwachstum in koordinierter Weise.[64] Die herrschende Synthetische Evolutionstheorie müsste an diesem und anderen Beispielen plausibel erklären können, wie in nur wenigen Generationen allein durch das Wechselspiel von zufälliger Mutation und Selektion eine derartig umfangreiche, koordinierte phänotypische Variation entstehen kann, die eines wechselseitigen Zusammenspiels vieler separater Entwicklungsparameter bedarf.
Evo-Devo zeigt an diesem Beispiel die erklärte Wirkungsweise: Kleine Ursache (eine oder ein paar quantitative, regulatorische Proteinänderungen, hervorgerufen) führt zu großer Wirkung (integrierte Veränderung der Schnabelform), gesteuert durch epigenetische Prozesse der Entwicklung, insbesondere durch ein breites adaptives Zellverhalten der Neuralleistenzellen des Schnabels und des Gesichtsumfelds.[65] Aus der gut erforschten Kenntnis der Entwicklung des Schnabels und seiner Modifikationen kann geschlossen werden, dass sich „recht umfangreiche Veränderungen der Schnabelgröße und Schnabelform mit ein paar regulatorischen Mutationen eher erreichen lassen als mit einer Summierung von langen Folgen kleiner Veränderungen“.[66] Nicht final erforscht ist in diesem Beispiel, wodurch die Veränderungen des Bmp4-Spiegels in der Entwicklung ausgelöst werden. Eine Möglichkeit sind genetische Zufallsmutationen, wahrscheinlicher sind Reaktionswege der Entwicklung auf den Stress der Tiere, der durch die anhaltende Veränderung des Nahrungsangebots entsteht. Diese Veränderung wurde ja von den Grants im Zusammenhang mit der Variation der Schnäbel dokumentiert.[65][67]
Beljajew genetische Zähmung von Silberfüchsen
Bekannt geworden ist der Versuch des russischen Genetikers Dmitri Konstantinowitsch Beljajew,[68] dessen Team über einen Zeitraum von 40 Jahren Silberfüchse auf Zahmheit selektierte. Man wollte erfahren, welche Konsequenzen die Selektion auf Zahmheit (bzw. gegen Aggression) nach sich zieht. Beljajews Hypothese war, dass es zu vielfältigen morphologischen, physiologischen und Verhaltensänderungen kommt, ähnlich wie bei über 10.000 Jahre zurückliegenden Domestikation des Wolfs zum Hund beobachtet. Für das Experiment wurden besonders zahm wirkende Füchse aus Pelztierzuchten ausgewählt, eine Zuchtlinie begründet und im folgenden auf Zahmheit (und nur darauf, nicht auf andere morphologische oder physiologische Änderungen) selektiert.
Beljajews Team hat sehr darauf geachtet, dass Mechanismen wie Inzucht oder Polygenie die Ergebnisse nicht beeinflussen konnten. Dennoch traten etwa von der vierten Generation an neben Verhaltensänderungen auch typische Haustiermerkmale, wie weiße Fellzeichnungen, hängende Ohren, kurze Beine und Schnauzen auch bei den Füchsen auf. Dies konnte Beljajew anhand paralleler Versuche an anderen Arten (Ottern, Ratten) im Kern reproduzieren.[69]
 Abb. 9 Das Silberfuchs-Experiment in Sibirien hat eine Reihe zu erwartender evolutionärer Veränderungen in der Entwicklung hervorgerufen.
Abb. 9 Das Silberfuchs-Experiment in Sibirien hat eine Reihe zu erwartender evolutionärer Veränderungen in der Entwicklung hervorgerufen.Mit Fortschreiten des Projekts konnten Veränderungen in den neurochemischen und neurohormonellen Mechanismen aufgedeckt werden, die für die genannten Bereiche eine wesentliche Rolle spielen, und zwar hauptsächlich der Cortisol-, Adrenalin- und Serotonin-Spiegel. Der stark abgesenkte Level der „Stresshormone“ Cortisol und Adrenalin und der abgesenkte Serotoninspiegel mit Zusammenhang zur Aggressivität sind plausible Erklärungsmuster für die zunehmende Zahmheit.[70] Diese Reaktionen in den Hormonenspiegeln können neben den Verhaltensänderungen auch zu den Entwicklungsänderungen in Beljajews Versuch beigetragen haben. Ein plausibles Modell geht davon aus, dass eine Verzögerung der angeborenen Furchtreaktion bei den Jungtieren die zunehmende Vertrautheit mit Menschen bewirkt. Diese Entwicklungsverzögerung ist durch entwicklungsbiologische Zwänge möglicherweise mit anderen Entwicklunhgsverzögerungen gekoppelt. Die typischen anatomischen Auswirkungen machen das Tier welpenähnlicher („Neotenie“), oft gekoppelt mit erhöhter Fruchtbarkeit. Besonders hat Trut darauf hingewiesen, dass der Fruchtbarkeitszyklus bei weiblichen Wildtieren genetisch normalerweise „fest verdrahtet“ ist und selbst in jahrzehntelangen direkten Züchtungsversuchen in der Vergangenheit nicht im Sinne der Züchter abgeändert werden konnte.[71] Solche Veränderungen gelangen aber im Versuch Beljajews als Beiprodukt, und zwar auf dem Weg ausschließlich über eine gezielte
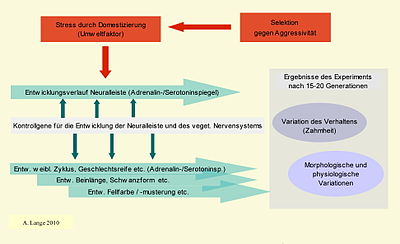 Abb. 10 Zusammenhang von Umwelt, Selektion, Kontrollgenen, physiologischen Prozessen, Entwicklung und Evolution im Experiment von Beljajew
Abb. 10 Zusammenhang von Umwelt, Selektion, Kontrollgenen, physiologischen Prozessen, Entwicklung und Evolution im Experiment von BeljajewVerhaltensselektion, es kam zu einer Reihe koordinierter Veränderungen im weiblichen Zyklus (um ca. 1 Monat früherer Eintritt der Fruchtbarkeit). Trut weist darauf hin, dass auch Schwangerschaften außerhalb der normalen Tragzeiten auftraten, aber keine der Jungen überlebt haben, die in saisonalen Zyklen geboren wurden, die vom natürlichen Zyklus abwichen. Ein Grund dafür konnte nicht gefunden werden.
Die beobachteten Veränderungen zeigen eine verblüffend schnell ablaufende Wandlung zum Haustier, wobei die meisten Merkmale unbeabsichtigt, als reines Beiprodukt der Selektion auf Zahmheit, auftraten. Für eine konventionelle genetische Erklärung über die Anhäufung von Punktmutationen ist der Wandel um ein Vielfaches zu schnell. Anzunehmen ist eine Änderung der Genexpression von (bisher nicht identifizierten) Mastergenen mit entwicklungssteuernder Wirkung. Möglich wäre ein rein epigenetischer Mechanismus über „Abschaltung“ (silencing) eines Entwicklungsgens durch DNA-Methylierung eines Entwicklungsgens bzw. seiner regulatorischen Sequenzen, die durch hormonelle Einflüsse möglich erscheint.
Evolution des Auges
Durch Analyse spontan entstandener Mutationen der Taufliege Drosophila, denen die Augen fehlen, ist es Genetikern gelungen, ein Schlüsselgen aus der Regulationskaskade der Augenentwicklung zu identifizieren.[72] Dieses Gen erwies sich als ein Transkriptionsfaktor, das heißt es kodiert ein Protein, welches an die DNA bindet und dadurch die Transkription anderer Gene verstärkt bzw. verhindert. Das pax6 genannte Gen gehört zu einer ganzen Familie regulatorischer Gene, die alle Entwicklungsvorgänge steuern. In einem aufsehenerregenden Versuch ist es den Forschern gelungen, durch künstlich induzierte Expression des Gens auch in anderen Körperteilen (funktionsfähige!) Augen zu erzeugen: An den Antennen, an der Flügelbasis, am Thorax usw. Durch heute fast routinemäßige Vergleiche mit dem Genom anderer Organismen erwies sich: Gene ähnlicher Sequenz, die aller Wahrscheinlichkeit nach homolog sind, wurden bei Tierarten aus nahezu allen daraufhin untersuchten Tierstämmen gefunden: z.B. bei Wirbeltieren (Maus, Mensch), Weichtieren (Muscheln, Tintenfische), Fadenwürmern uv.a., und in allen Fällen war es (neben einigen anderen Aufgaben) an der Entwicklung von Augen beteiligt.[73] Sogar die primitiven Augenflecken des Plattwurms Dugesia und die Linsenaugen am Schirmrand der Würfelqualle Tripedalia cystophora wurden von dem gleichen bzw. einem homologen Gen gesteuert. Dies ist deswegen verblüffend, weil sich diese Tiere in der Evolution mindestens seit dem Kambrium vor über 540 Millionen Jahren auseinanderentwickelt haben. Trotzdem war es möglich, mit dem Gen der Maus bei der Taufliege Augen zu induzieren. Die Augenentwicklung erfordert das fein abgestimmte Zusammenspiel einiger hundert Effektorgene. Dies lässt sich am ehesten dadurch erklären, dass diese Gene, die im Jargon der Genetiker „stromabwärts“ (engl: downstream) von pax6 liegen, Bindungsstellen (sog. cis-regulatorischen Elemente) für das Pax6-Protein enthalten. Pax6 ist dabei nur ein einziger Faktor in einem fein austarierten Netzwerk von Signalketten und Steuerungswegen, welches im Detail noch kaum bekannt ist. Für solche in der Evolution über hunderte von Millionen Jahren konservierte Entwicklungswege hat Sean Carroll den Ausdruck Tiefenhomologie geprägt.[74]
Bei Betrachtung der Augen im Detail zeigt es sich allerdings, dass es nicht unbedingt wahrscheinlich ist, dass eine einfache Weiterentwicklung eines einmal entstandenen Auges bereits die ganze Geschichte ausmacht. Zwar haben alle Augen aller Tiere dasselbe lichtempfindliche Molekül, eine Variante des Sehpigments Rhodopsin (welches bereits bei Einzellern und auch bei Prokaryonten vorkommt). Neben dem Sehpigment gehört bereits zu den einfachsten Augen ein lichtabschirmendes Pigment (zum Richtungssehen), außer bei den allereinfachsten Konstruktionen auch ein lichtdurchlässiger „Glaskörper“, der aus einem durchsichtigen Protein („Crystallin“ genannt) besteht. Beim Vergleich verschiedener Augentypen zeigt es sich, dass unterschiedliche Organismen unterschiedliche Pigmente (Melanin, Pterin, Ommochrom) und vor allem völlig unterschiedliche Crystalline verwenden. Beinahe alle Crystalline sind darüber hinaus Enzyme oder Abkömmlinge von solchen, die andernorts im Körper noch völlig andere, essenzielle Aufgaben zu erfüllen haben.[75] Außerdem gibt es die Rezeptorzellen in zwei Ausfertigungen, als „rhabdomerische“ und „ciliare“ Rezeptoren mit völlig unterschiedlichem Zellbauplan. Die rhabdomerische Ausfertigung findet sich bei den Arthropoden, die ciliare bei den Wirbeltieren, aber auch bei den Würfelquallen. Die wesentlichen Bestandteile des Auges sind also zwar unter in der Evolution hochgradig konservierter Entwicklungskontrolle, darunter aber beinahe zufällig „durcheinandergewürfelt“. Dies erscheint am ehesten dadurch erklärbar, dass beim Aufbau des immer komplexer werdenden Auges immer mehr ursprünglich unabhängig und für einen anderen Zweck entstandene Strukturen, aber auch Entwicklungspfade und Signalwege neben ihrer ursprünglichen Funktion auch bei der Augenentwicklung verwendet wurden. Dies ist am besten dadurch erklärbar, dass sie cis-regulatorische Sequenzen evolutiv neu erworben haben, die durch Kontrollgene der Augenentwicklung wie z.B. pax6 steuerbar sind[76] (die Gene sind ja in allen Körperzellen identisch vorhanden!). Der Entwicklungsweg insgesamt ist also homolog, die weiteren Strukturen wie Pigmente, Glaskörper, Linse etc. sind aber vermutlich konvergente Bildungen. Für ihre Entstehung sind aber weder neue Proteinfamilien oder auch nur neue Gene erfunden worden, sondern bereits bestehende wurden umfunktioniert („rekrutiert“ oder auch „ko-optiert“).[77]
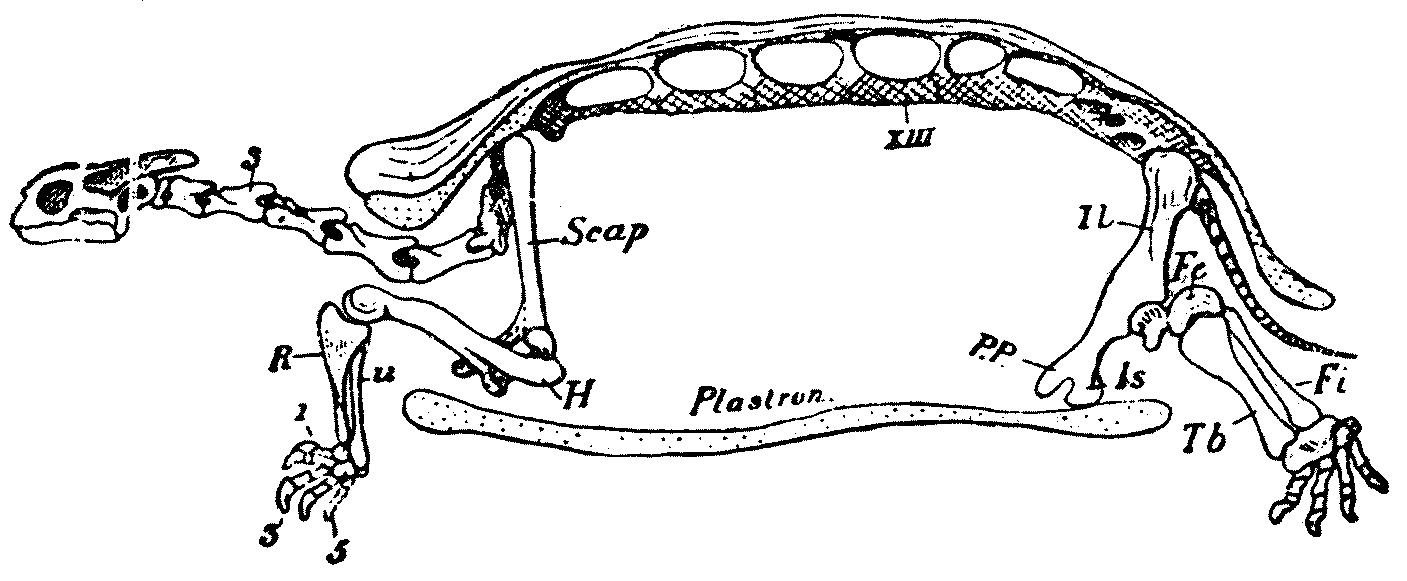
 Abb. 11 Die Entstehung des Schildkrötenpanzers erzwang eine komplizierte Verlagerung des Schultergürtels (Scapula) von außerhalb wie sonst bei Echsen üblich nach innerhalb der Rippenbögen, bzw. hier innerhalb des Panzers.
Abb. 11 Die Entstehung des Schildkrötenpanzers erzwang eine komplizierte Verlagerung des Schultergürtels (Scapula) von außerhalb wie sonst bei Echsen üblich nach innerhalb der Rippenbögen, bzw. hier innerhalb des Panzers.Skelettumbauten bei der Entstehung des Schildkrötenpanzers
Der Panzer der Schildkröten ist eine anatomisch äußerst komplexe Bildung, an der sowohl Knochen (v.a. die Rippen) als auch Hautverknöcherungen (Osteoderm) beteiligt ist. In jüngerer Zeit hat diese Struktur, die seit den anatomischen Studien des 19.Jh. als rätselhaft galt, erneut starke Aufmerksamkeit bei Evolutionsbiologen gefunden, weil eine einflussreiche Theorie postulierte, die bekannt gewordenen entwicklungsbiologischen Fakten würden eine Entstehung des Panzers durch eine Makromutation (auch Saltation genannt) nahelegen und dadurch die synthetische Evolutionstheorie partiell widerlegen.[78][79] Es hat sich allerdings gezeigt, dass eine plausible Erklärung auch ohne Saltation möglich ist. Neben neueren entwicklungsgenetischen Erkenntnissen war ein wichtiger Grund, dass in jüngster Zeit erstmals Fossilien von Schildkröten mit unvollständiger Panzerung neu entdeckt worden sind.[80][81]
Die Entstehung des Panzers kann man sich demnach in etwa so vorstellen: Entscheidendes morphologisches Merkmal ist die Entwicklung der Rippen, die nicht wie bei allen anderen Wirbeltieren den Brustraum mehr oder weniger umschließen, sondern die in die obere (anatomisch: dorsale) Körperwand einwachsen. Hier verbinden sie sich mit Hautverknöcherungen, die vermutlich unabhängig entstanden sind (entsprechende Bildungen sind bei Wirbeltieren weit verbreitet[82]). In der Embryonalentwicklung werden beide Bildungen über einen zellulären Signalweg (den sog. wnt-Weg) gesteuert, der sonst z.B. an der Bildung von Gliedmaßen beteiligt ist.[83] Eine entsprechende partielle Umbildung von bereits bestehenden Entwicklungspfaden (und ihren Genen) wird als „Kooptation“ bezeichnet und tritt offensichtlich häufiger auf, auch wenn die Einzelheiten nicht völlig geklärt sind. Wahrscheinlichste Erklärung ist die Umwandlung von „genetischen Schaltern“ (sog. cis-regulatorischen Sequenzen), durch die die Expression eines Gens in neuem Funktionszusammenhang möglich ist, ohne dass das Gen selbst sich verändert. Im Zuge dieser Umgestaltung faltete sich beim Schildkrötenembryo die seitliche Körperwand nach innen. Dadurch wurde der Schultergürtel, der sonst oberhalb der Rippen liegt, in das Innere verlagert (die ursprüngliche Lagebeziehung lässt sich an den an ihm ansetzenden Muskeln noch ablesen[84]). Der Bauchpanzer (das Plastron) ist parallel dazu auf noch nicht ganz geklärtem Wege entstanden. Vermutlich ist an seiner Entstehung ein zellulärer Signalweg beteiligt, der demjenigen entspricht, der zur Verknöcherung der Schädelknochen führt.[85] Interssanterweise besaß ein in China entdecktes, 220 Millionen Jahre altes Fossil einer Wasserschildkröte einen vollständigen Bauchpanzer, währen ein Rückenpanzer fehlte.
Obwohl damit noch nicht alle Einzelheiten geklärt sind, existiert ein überzeugendes Modell der graduellen Entstehung damit auch für eine zunächst rätselhafte Struktur wie den Schildkrötenpanzer. Daran zeigt sich schön, wie durch intensive Zusammenarbeit aller Disziplinen (hier: Paläontologie, Embryologie, Entwicklungsgenetik) auch zunächst unlösbar erscheinende Probleme nach und nach enträtselt werden können.
Evo-Devo liefert auch eine Erklärung für den Wegfall der Beine beim evolutionären Übergang von Echsen zu Schlangen. Nach Paul Layer war hierfür die Expressionshemmung von nur 2 Hox-Genen (c6 und c8) erforderlich, wie empirisch nachgewiesen wurde.[86]
Konsequenzen für die Evolutionstheorie
Evo-Devo erklärt die Evolution organismischer Form mit den kausal-mechanistischen Änderungen des Gesamtsystems Entwicklung (bestehend aus den teilautonomen Subsystemen Genom, Zellen, Zellverbänden, Organismus) und der Wirkung von Umwelteinflüssen. Demgegenüber ist die klassische darwinistische Evolutionstheorie eher statistisch-deskriptiv, da sie dem Organismus inhärente Faktoren für die Entstehung von Variation und Innovation nicht berücksichtigt. Vielmehr überlässt sie das evolutionäre Geschehen ausschließlich der zufälligen Mutation und der externalistisch wirkenden natürlichen Selektion. Richtung bekommt die Evolution aus klassischer Sicht ausschließlich durch das Wirken der Selektion.
Die Bedeutung des Zufalls zur Erzeugung von morphologischer Variation wird von der evolutionären Entwicklungsbiologie neu bewertet. Es existieren erklärbare, vorhersehbare, regelhafte Gesetzmäßigkeiten der Entwicklungsprozesse, die die (im Kern immer noch zufällige) Variation stark beeinflussen. Das ist einer der wesentlichen Unterschiede zwischen Evo-Devo und Synthetischer Evolutionstheorie. Die natürliche Selektion leistet nach wie vor ihren Beitrag (survival of the fittest), aber ihr Erklärungswert für die Entstehung organismischer Form und Komplexität wird relativiert und dem Erklärungswert von Konstruktion gegenübergestellt. Selektion bleibt dabei eine Rahmenbedingung der Evolution. Sie kann aber nur „digital entscheiden“ über das, was die Entwicklung ihr vorgibt.
Das von einigen Autoren als "genzentristisch" bezeichnete und das besonders durch Richard Dawkins forcierte, reduktionistische Denken wird von Evo-Devo kritisiert.[87] Für Forscher wie Gilbert, Kirschner, Müller, Pigliucci, West-Eberhard und andere sind Gene nicht der ausschließliche und auch nicht der Hauptadressat der natürlichen Selektion. Es wird auch nicht von einer streng deterministischen Beziehung zwischen genotypischer und phänotypischer Evolution ausgegangen. Die epigenetischen Entwicklungsprozesse gelten im Zusammenhang mit der Evolution organismischer Form als unabdingbar für die Erzeugung des Phänotyps und die Generierung morphologischer Variation und Innovation. Das beseitigt nicht die überragende Bedeutung der Gene, relativiert sie aber bis zu einem gewissen Punkt.
Die Evolutionstheorie befindet sich somit in einer post-genozentrischen Ära (Müller-Wille, Staffan und Rheinberger[88][89]), in der sie die komplexen, rekursiven, epigenetischen Zusammenhänge aufgreift und in einen kongruenten Theorierahmen einbinden muss. Als ein Beispiel wurde durch die Altenberg-16-Gruppe mit dem 2010 publizierten Konzept der "Extended Synthesis" ein neuer fächerübergreifender Ansatz vorgestellt.[90]
Die neue systemische Sicht auf die Entwicklung
Einige Forscher, unter ihnen Brian K. Hall, Rudy Raff, Gerd B. Müller, Walter Gilbert, Marc Kirschner, Massimo Pigliucci, Mary Jane West-Eberhard, gehen heute über die Analyse der Genregulationen hinaus und erweitern die Untersuchung auf das gesamte System Entwicklung als einen komplexen genetisch/epigenetisch evoluierten Apparat, der mit der Umwelt als ein integriertes, multikausales, nicht-lineares, offenes System interagiert. Kausale Erklärungen für die Entstehung und Variation organismischer Formen werden von diesen Forschern nicht ausschließlich auf der Gen-/Genregulationsebene gesucht, sondern auch bzw. primär in den epigenetischen Eigenschaften von Zellen, Zellaggregaten, Selbstorganisation, Organismus und Umwelt (siehe Abbildung 5). Epigenese wird von Evo-Devo nicht synonym mit Epigenetik gebraucht, d. h. es sind nicht genregulatorische Aspekte wie DNA-Methylierungen gemeint, sondern die Summe der nicht programmierten Faktoren in der Embryonalentwicklung, wie z. B. chemische, physikalische oder dynamische Faktoren, die den Entwicklungsprozess organisatorisch beeinflussen.
Eco-Evo-Devo
Die Betrachtung der Umwelt erweitert die Disziplin maßgeblich (s. Abb.2). Man spricht in diesem Kontext auch von Eco-Evo-Devo.[91] Leigh Van Valen definierte 1973 Evolution als „die Kontrolle der Entwicklung durch die Umwelt“. In dieses Feld fallen Überlegungen zur phänotypischen Plastizität oder Entwicklungsplastizität, d.h. die Fähigkeit des Organismus, in Abhängigkeit von wechselnden Umweltbedingungen unterschiedliche Phänotypen auszubilden. Zu nennen sind hier neben dem schon zitierten C. H. Waddington West-Eberhards Buch Development Plastizity and Evolution (2003) sowie Gilbert und Epels interdisziplinäres Buch Ecological Development Biology (2009). Die Einbeziehung der Umwelt und damit die Interaktionsfähigkeit der Evolution mit der Umwelt auf dem Weg über die Entwicklung wird zu einem wachsenden Forschungsthema (siehe Die Evo-Devo-Forschungsthemen). Erstmals kann in diesem erweiterten Zusammenhang eine kausale Beziehung zwischen Umwelt und Entwicklung hergestellt werden.
Literatur
Konzeptionelle Grundlagen
- Amundson, Ron (2005), The Changing Role of the Embryo in Evolutionary Thought
- Arthur, Wallace (2004), Biased Embryos and Evolution. Cambridge University Press
- Brigandt, Ingo (2010): Jenseits des Neodarwinismus? Neuere Entwicklungen in der Evolutionsbiologie. In: Sarasin, Philipp, Sommer, Marianne: Evolution – Ein interdisziplinäres Handbuch. J. B. Metzler, S. 115–126.
- Caroll, Sean B. (o. Jg., ersch. 2008) EvoDevo – Das neue Bild der Evolution. Berlin. (Orig.: Endles Forms Most Beautiful, USA 2006)
- Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology
- Kirschner, Mark C., Gerhart, John C. (2007) Die Lösung von Darwins Dilemma – Wie Evolution komplexes Leben schafft. Rowohlt. Orig.: The Plausibility of Life (2005)
- Layer, Paul G. EvoDevo: Die molokulare Entwicklungsbiologie als Schlüssel zum Verständnis der Evolutionstheorie.
- Minelli, Alessandro & Giuseppe Fusco (Hrsg.) (2008): Evolving pathways - key themes in evolutionary developmental biology. Cambridge University Press, Cambridge/ New York, 426 S., ISBN 9780521875004
- Müller, Gerd B., Newman, Stuart A. (2003) Origination of Organismal Form – Beyond the Gene in Development and Evolutionary Biology. MIT-Press.
- Müller, Gerd B. (2008) Evodevo as a discipline in Minelli & Fusco (2008)
- Nüsslein-Vollhard, Christiane (2006) Das Werden des Lebens – Wie Gene die Entwicklung steuern. dtv.
- Pigliucci, Massimo, Müller, Gerd (2010), Evolution – the Extended Synthesis. MIT Press
- West-Eberhard, Mary Jane (2003), Development Plastizity and Evolution. Oxford University Press.
Weiterführende Literatur und Internetartikel
- BDTNP Barkeley Transcription Drosophila Network Project
- Gilbert, Scott F., Epel, David (2009), Ecological Development Biology. Inegrating Epigenetics, Medicine and Evolution. Sinauer Ass. USA.
- Jablonka, Eva, Lamb, Marion J. (2005), Evolution in four Dimensions. Genetic, Epigenetic, Behaviroal and Symbolic Variation in the History of Lfe. MIT Press.[3]
- Müller, Gerd B., Newman, Stuart A. (2005) The Innovation Triad. An EvoDevo Agenda, in: Journal of Experimental Zoology 304B: 487-503
- Neukamm, M. (2009) Evolutionäre Entwicklungsbiologie: Neues Paradigma. Laborjournal 15(11), 24-27 (pdf).
- Nihjout, Frederic (2006), Researchers evolve a komplex genetic trait in the labratory
- Pigliucci, Massimo (2008), What, if anything, Is an Evolutionary Novelty? Philosophy of Science 75 (12/2008), S. 887–898
- Tomancak, Pavel et al. (2007) (Max-Planck-Ges.) Patterns of gene expression in animal development
- Trut, Lyudmila N. (1999), Early Canid Domestication: The Farm-Fox Experiment, American Scientist, Bd. 87
- Young, Emma (2008) Rewriting Darwin. The new non-genetic inheritance. New Scientist magazine.
Einzelnachweise
- ↑ Kirschner Kirschner, Mark C., Gerhart, John C. (2007) Die Lösung von Darwins Dilemma – Wie Evolution komplexes Leben schafft. Rowohlt. Orig. The Plausibility of Life (2005)
- ↑ Pigliucci/Müller (Hrsg.) (2010) Evolution - The Extended Synthesis. MIT Press. Kap. 10 u. 12
- ↑ West-Eberhard, Mary Jane (2003) Development Plastizity and Evolution. Oxford Univesity Press
- ↑ Einen umfassenden Überblick über die Rolle des Embryos in der Geschichte der Evolutionstheorie gibt das Buch von Ronald Amundson (2005) The Changing Role of the Embryo in Evolutionary Thought. Roots of Evo-Devo. Cambridge University Press
- ↑ Brian K. Hall (2000): Balfour, Garstang and de Beer: The First Century of Evolutionary Embryology. American Zoologist 40(5): 718-728
- ↑ Alan C. Love & Rudolf A. Raff (2003): Knowing your ancestors: themes in the history of evo-devo. Evolution and Development 5(4): 327–330.
- ↑ Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology S. 471
- ↑ Jablonka, Eva & Lamb, Marion J. (2005), Evolution in four Dimensions. Genetic, Epigenetic, Behaviroal and Symbolic Variation in the History of Lfe. MIT Press S. 261–266
- ↑ Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology S. 474
- ↑ M. Pigliucci, G. Müller (2010): Evolution. The Extended Synthesis, Kap.1: Elements of an Extended Evolutionary Synthesis
- ↑ Pigliucci, Müller 2010, Kap.1: Elements of an Extended Evolutionary Synthesis
- ↑ Pigliucci, Müller 2010 (Hrsg.) S.13
- ↑ West-Eberhard, Mary Jane (2003), S.6ff.
- ↑ Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology S. 470
- ↑ Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology S. 473
- ↑ Master-Kontrollgene oder Masterregulatorgene sind Gene, „die fortlaufend einen oder mehrere Transkriptionsfaktoren in gewissen sich differenzierenden Zellen exprimieren“ (Kirschner und Gerhard 2005 S.384)
- ↑ Nüsslein-Vollhard, Christiane (2006) Das Werden des Lebens – Wie Gene die Entwicklung steuern. S. 93
- ↑ Layer, Paul G. Evo-Devo: Die molekulare Entwicklungsbiologie als Schlüssel zum Verständnis der Evolutionstheorie S.7
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005): The Innovation Triad. An Evo-Devo Agenda in Journal of Experimental Zoology 304B
- ↑ West-Eberhard, Mary Jane (2003) S.7 mit Bezug auf Buss 1987
- ↑ Minelli, A., Fusco, G. (Hrsg.) (2008) Evolving Pathways – Key Themes in Evolutionary Developmental Biology S.2.
- ↑ Müller, Gerd B. (2009): Extending the Evolutionary Synthesis
- ↑ Caroll, Sean B. (o. Jg., ersch. 2008) Evo-Devo – Das neue Bild der Evolution. Berlin. (Orig.: Endless Forms Most Beautiful, USA 2006)
- ↑ Müller, Gerd B. (2007) Evo-devo: extending the evolutionary synthesis in Nature Review Genetics S.2
- ↑ Gilbert, Scott F. (2003) The reactive Genome in Müller/Newman (Hrsg.) (2003) Origination of Organismal Form. Beyond the Gene in Development and Evolutionary Biology S. 87–101f
- ↑ Gilbert in Müller-Newman 2003, S.93f
- ↑ Spektrum der Wissenschaft 03/2010. Meinhardt, Hans: Die Simulation der Embryonalentwicklung.
- ↑ Kirschner/Gerhard a.a.O. S. 264
- ↑ Kirschner/Gerhart a.a.O. S.302
- ↑ Kirschner/Gerhart a.a.O. S.382
- ↑ Kirschner/Gerhart a.a.O. S.382
- ↑ Kirschner/Gerhart a.a.O. S.328
- ↑ Kirschner/Gerhart a.a.O. S.333
- ↑ Kirschner/Gerhart a.a.O. S.369
- ↑ Christina L. Richards, Oliver Bossdorf, and Massimo Pigliucci (2010): What role does heritable epigenetic variation play in phenotypic evolution? BioScience 60(3): 232-237.
- ↑ Ryan A. Rapp, Jonathan F. Wendel(2005): Epigenetics and plant evolution. New Phytologist, 168: 81–91.[1]
- ↑ Eva Jablonka and Marion Lamb: Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life (MIT, 2005)
- ↑ Jablonka, E., Raz, G. (2009): Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution. Quarterly Review of Biology 84: 131–176.
- ↑ Finnegan, E.J. (2002): Epialleles—a source of random variation in times of stress. Current Opinion in Plant Biology 5: 101–106.
- ↑ West-Eberhard, M.J. (2003). Developmental Plasticity and Evolution. Oxford University Press.
- ↑ Pigliucci, Müller (2010) u. dort: G.Müller Kap.12 Epigenetic Innovation S. 311
- ↑ Pigliucci, Massimo (2008) What, if anything, Is an Evolutionary Novelty? Philosophy of Science 75 (Dec.2008) S. 887–898
- ↑ a b Müller, Gerd B. u. Newman, Stuart A. (2005): The Innovation Triad. An Evo-Devo Agenda in Journal of Experimental Zoology 304B: 487-503
- ↑ Pigliucci, Müller (2010) u. dort: G. Müller Kap.12 Epigenetic Innovation S. 314ff.
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) S.494
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005) S.494
- ↑ West-Eberhard, M.J. (2003): Development Plastizity and Evolution. Oxford University Press S. 157
- ↑ Müller, Gerd B. u. Newman, Stuart A. (2005): S.494
- ↑ Pigliucci, Müller (2010) u. dort: G.Müller Kap.12 Epigenetic Innovation S.320f
- ↑ Carroll, Sean B. (o. Jg., ersch. 2008) Evo-Devo – Das neue Bild der Evolution. Berlin. (Orig.: Endles Forms Most Beautiful, USA 2006)
- ↑ Arthur, Wallace (2004), Biased Embryos and Evolution. Cambridge University Press
- ↑ Layer, Paul G. EvoDevo: Die molokulare Entwicklungsbiologie als Schlüssel zum Verständnis der Evolutionstheorie S. 5f
- ↑ Komplex meint hier im Sinn der Komplexitätsforschung: interdependent (Genom – Entwicklung – Evolution – Umwelt), multikausal, nicht-linear, offen usw (Systemtheorie)
- ↑ Anm.: Die besten Geräte haben nach Metscher (Metscher 2009) heute eine Auflösung von 60 Nanometer. 1 nm = 1 Milliardstel m bzw. 1 Millionstel mm (z.Vgl.: Eine menschliche Zelle hat im Durchschnitt eine Größe von 10-20 Mikrometer, das ist um die Größenordnung 1000 größer.)
- ↑ a b c Tomancak, Pavel et al. (Max Planck Ges.) (2007): Patterns of gene expression in animal development. Kurzfassung des Artikels.: Global analysis of patterns of gene expression during Drosophila embryogenesis. Genome Biol.; 8, no. 7 S. 145.1–145.34
- ↑ Metscher, Brian (2009): MicroCT for comparative morphology: simple staining methods allow high contrast 3D imaging of diverse non-mineralized animal tissues. http://www.biomedcentral.com/1472-6793/9/11
- ↑ Waddington, Conrad Hal (1953): Genetic Assimilation of an Acquired Charakter. Evolution Vol.7 No.2, S. 118–126
- ↑ Kirschner/Gerhart a.a.O. S. 316
- ↑ Caroll, Sean B. (o. Jg., ersch. 2008) Evo-Devo. S. 192–209
- ↑ NZ Neue Zürcher Zeitung vom 12. Juli 2006 Die Darwinfinken – Evolution im Zeitraffer. Erkenntnisse eines britischen Ehepaars aus mehr als 30 Jahren Forschung auf den Galapagosinseln
- ↑ Kirschner Kirschner, Mark C., Gerhart, John C. (2007) Die Lösung von Darwins Dilemma S.321f.
- ↑ Kirschner/Gerhart a.a.O. S. 3121 mit Bezug auf Studien von Clifford Tabin an Darwinfinken
- ↑ Kirschner/Gerhart a.a.O. S. 3121 mit Bezug auf Studien von Clifford Tabin an Darwinfinken
- ↑ Kirschner/Gerhart a.a.O. S. 318
- ↑ a b Kirschner/Gerhart a.a.O. S. 319
- ↑ Kirschner/Gerhart a.a.O. S. 319
- ↑ NZ Neue Zürcher Zeitung vom 12. Juli 2006 Die Darwinfinken – Evolution im Zeitraffer. P. u. R. Grant sehen Rückkreuzungen (Introgression) als eine adaptive Ursache für die Variation der Schnäbel S. 3
- ↑ Trut, Lyudmila N. (1999) Early Canid Domestication: The Farm-Fox Experiment American Scientist Vol 87 "
- ↑ Trut, Lyudmila N. (1999) Early Canid Domestication: The Farm-Fox Experiment. American Scientist Vol. 87 S. 165f.
- ↑ Lyudmila Trut, Irina Oskina, Anastasiya Kharlamova (2009): Animal evolution during domestication: the domesticated fox as a model. Bioessays 31(3): 349–360
- ↑ Trut, Lyudmila N. (1999) Early Canid Domestication: The Farm-Fox Experiment. American Scientist Vol. 87 S. 167
- ↑ Georg Halder, Pstrick Callaerts, Walter J.Gehring (1995): Induction of ectopic eyes by targetes expression of the eyless gene in Drosophila. Science 267: 1788-1792
- ↑ Walter J.Gehring & Kazuho Ikeo (1999): Pax6: mastering eye morphogenesis and eye evolution. Trends in Genetics 15(9): 371-377.
- ↑ vgl.: Neil Shubin, Cliff Tabin, Sean Carroll (2009): Deep homology and the origins of evolutionary novelty. Nature 457: 818-823.
- ↑ Joram Piatigorsky, G.Wistow (1991): The recruitment of crystallins: new functions precede gene duplication. Science 252: 1078-1079.
- ↑ Pavel Vopalensky & Zbynek Kozmik (2009): Eye evolution: common use and independent recruitment of genetic components. Philosophical Transactions of the Royal Society B 364: 2819-2832
- ↑ Joram Piatigorsky (2008): A Genetic Perspective on Eye Evolution: Gene Sharing, Convergence and Parallelism. Evolution: Education and Outreach 1(4): 403-414.
- ↑ Gilbert, S.F., Loredo, G.A., Brukman, A., Burke, A.C. (2001): Morphogenesis of the turtle shell:the development of a novel structure in tetrapod evolution. Evolution and Development 3:47–58.
- ↑ Rieppel, Olivier (2001): Turtles as hopeful monsters. BioEssays 23(11): 987-991
- ↑ Chun Li, Xiao-Chun Wu, Olivier Rieppel, Li-Ting Wang & Li-Jun Zhao (2008): An ancestral turtle from the Late Triassic of southwestern China. Nature 456(27):497-501.
- ↑ Walter G. Joyce, Spencer G. Lucas, Torsten M. Scheyer, Andrew B. Heckert, Adrian P. Hunt (2009): A thin-shelled reptile from the Late Triassic of North America and the origin of the turtle shell. Proccedings of the Royal Society B 276: 507-513.
- ↑ Eine Übersicht in: Matthew K. Vickaryous and Jean-Yves Sire (2009): The integumentary skeleton of tetrapods: origin, evolution, and development. Journal of Anatomy 214: 441-464.
- ↑ Shigehiro Kuraku, Ryo Usuda, Shigeru Kuratani (2005): Comprehensive survey of carapacial ridge-specific genes in turtle implies co-option of some regulatory genes in carapace evolution. Evolution and Development 7(1): 3-17.
- ↑ Shigeru Kuratani, Shigehiro Kuraku, Hiroshi Nagashima (2011): Evolutionary developmental perspective for the origin of turtles: the folding theory for the shell based on the developmental nature of the carapacial ridge. Evolution and Development 13(1): 1-14[2]
- ↑ Scott F. Gilbert, Gunes Bender, Erin Betters, Melinda Yin, Judith A. Cebra-Thomas (2007): The contribution of neural crest cells to the nuchal bone and plastron of the turtle shell. Integrative and Comparative Biology 47(3): 401–408.
- ↑ Layer, Paul G. EvoDevo: Die molekulare Entwicklungsbiologie als Schlüssel zum Verständnis der Evolutionstheorie S. 8
- ↑ John Dupré (2009) Darwins Vermächtnis. Die Bedeutung der Evolution für die Gegenwart des Menschen, u. dort bes. Kap. „Der genozentrische Fehlschluss“ S.89ff. Suhrkamp
- ↑ S.Müller-Wille, H-J. Rheinberger (2009) Das Gen im Zeitalter der Postgenomik. Eine wissenschaftliche Bestandsaufnahme. Suhrkamp
- ↑ John Dupré (2009) Darwins Vermächtnis; u. dort bes. Kap.: Der genozentrische Fehlschluss S.89ff. Suhrkamp
- ↑ . M. Pigliucci, G. Müller (Hrsg.) (2010): Evolution. The Extended Syntesis.
- ↑ Gilbert, Scott F. (2003): The morphogenesis of evolutionary development biology S. 473




Wikimedia Foundation.