- Futterkette
-
Der Begriff Nahrungskette bezeichnet in der Ökologie und in der Ökotoxikologie die linearen energetischen und stofflichen Beziehungen von aufeinanderfolgenden Organismenpopulationen, bei denen sich die Organismen des jeweils folgenden Trophieniveaus von den Organismen des vorangegangenen Trophieniveaus ernähren.
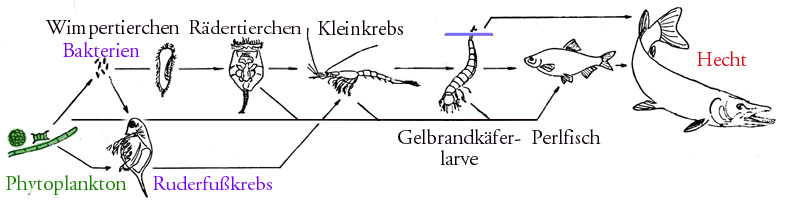
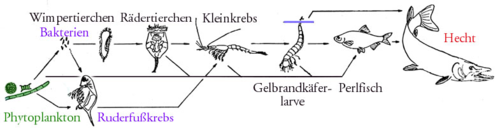 Schematische Nahrungskette in einem europäischen See (ohne Destruenten)
Schematische Nahrungskette in einem europäischen See (ohne Destruenten)Inhaltsverzeichnis
Das Konzept der Nahrungskette und der Stoffkreislauf
 Beispielhafte Darstellungen festländischer und mariner Nahrungsketten
Beispielhafte Darstellungen festländischer und mariner NahrungskettenDas Konzept der „Nahrungskette“ ist ein in verschiedener Hinsicht vereinfachtes Konzept linearer Energie- und Stoffflüsse im Ökosystem. Der Begriff wurde vor allem durch den britischen Zoologen und Ökologen Charles Elton in der ersten Hälfte des 20. Jahrhunderts propagiert. Auf ihn geht auch das Konzept der Elton’schen Zahlenpyramide zurück, die als Biomassepyramide oder als Energiepyramide der verschiedenen Trophieniveaus eines Ökosystems charakterisiert. Je nach Fragestellung werden daher entweder die gespeicherte und die in der Nahrungskette weitergegebene Energie im Rahmen von Energieflussmessungen oder aber einzelne Stoffe von Stoffflüssen (Kohlenstoff, Stickstoff usw.) untersucht.
Die Arten eines Ökosystems lassen sich verschiedenen Trophieniveaus (Nahrungskettenebenen) zuordnen. Das vereinfachte und meist idealisierte Modell der Nahrungskette unterscheidet zwischen Primärproduzenten (grüne Pflanzen oder chemosynthetische Bakterien), Konsumenten und Destruenten. Ein (vereinfachtes) Beispiel einer linearen Nahrungskette ist die Kette Gras – Zebra – Löwe.
Ein (vereinfachtes) Beispiel aus dem Süßwasser ist die Nahrungskette Kieselalge – Wasserfloh – Jungfisch – Raubfisch. Ein Beispiel aus dem Weltmeer ist die Nahrungskette Kleinalgen – Antarktischer Krill – Bartenwale.
Das älteste fossile Zeugnis einer dreigliedrigen Wirbeltier-Nahrungskette stammt aus den 290 Millionen Jahre alten frühpermischen Ablagerungen des „Humberg-Sees“ aus dem Saar-Nahe-Becken. Im Verdauungstrakt eines Fossils des etwa 70 Zentimeter[1] langen Xenacanthiformen Triodus, ein „Süßwasserhai“, fand sich die Larve einer temnospondylen Amphibie, die einen Stachelhai erbeutet hatte. Damit sind drei Trophieniveaus zugleich erhalten.[2] [3]
Destruenten (Pilze, Bakterien, aber auch an der mechanischen Zersetzung beteiligte Würmer, Gliedertiere usw.) entnehmen ihre Nahrung allen übrigen Trophieniveaus und führen dabei die Stoffe aus der Nahrungskette zurück, so dass sich die Form eines Stoffkreislaufs ergibt (vgl. z. B. Stickstoff- und Kohlenstoffkreislauf).
Reale Nahrungsketten und Nahrungsnetze
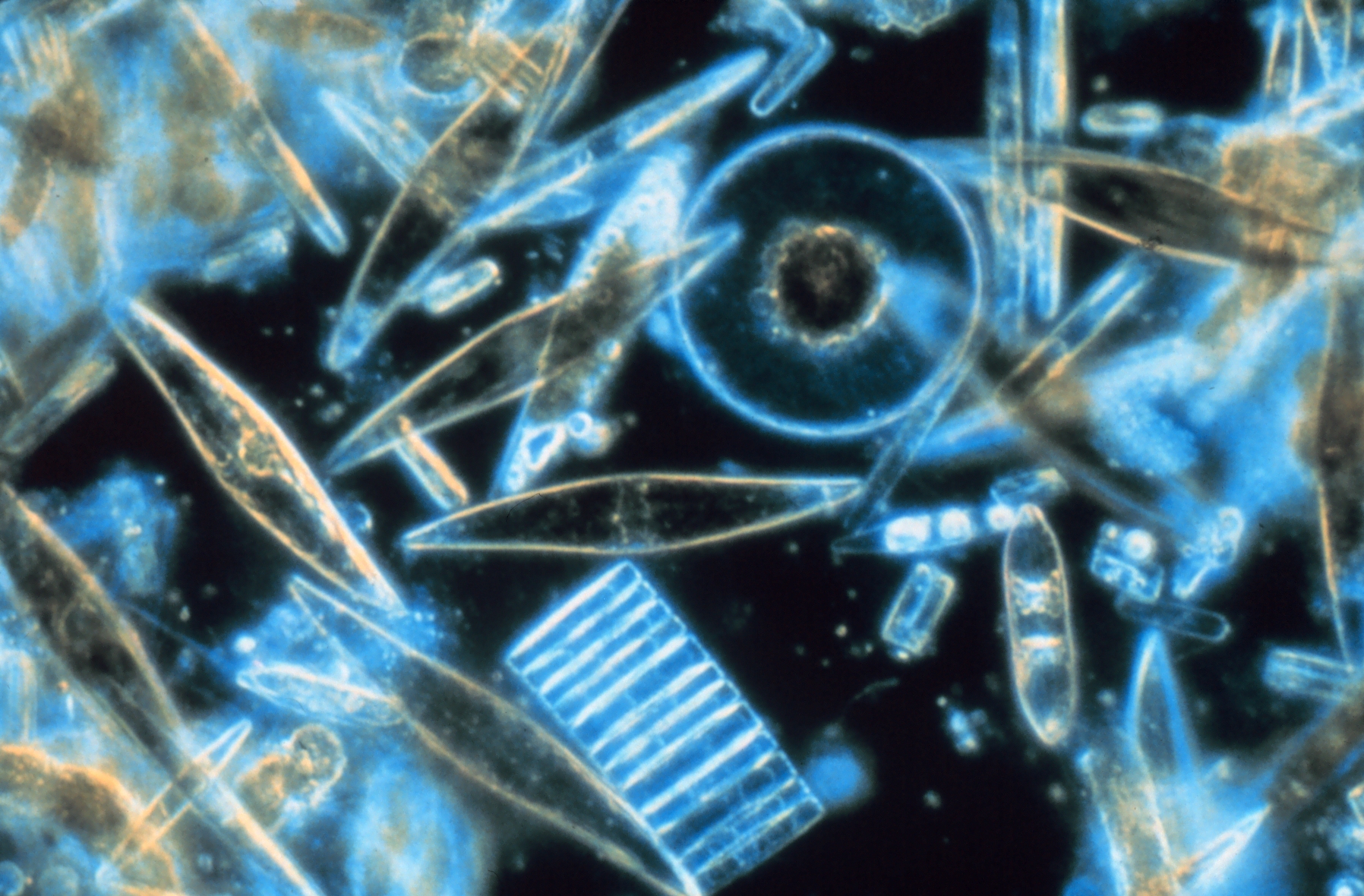
 Mikroskopisches Phytoplankton wie diese Kieselalgen stehen an der Basis der Nahrungsketten der marinen Ökosysteme.
Mikroskopisches Phytoplankton wie diese Kieselalgen stehen an der Basis der Nahrungsketten der marinen Ökosysteme.
(McMurdo-Sund, Antarktis)Der Begriff der Nahrungskette ist für fast alle natürlichen Ökosysteme eine sehr stark vereinfachende Abstraktion, so dass sich üblicherweise eher das Konzept des Nahrungsnetzes eignet, die Komplexität im Ernährungsgefüge annähernd richtig darzustellen.
Arten, die sowohl pflanzliche als auch tierische Nahrung aufnehmen (z. B. Schaben, Möwen, Schweine, Schimpansen) oder die sich teilweise von lebenden Tieren und teilweise von Kadavern ernähren (z. B. Hyänen, auch Löwen), lassen sich allerdings weder in einem Schema der Nahrungskette, noch einem solchen des Nahrungsnetzes übersichtlich darstellen, weshalb ein „reales Nahrungsnetz“ aus der Natur ein sehr komplexes und schwer durchschaubares Gebilde darstellen würde. Viele Arten ändern ihre Nahrungszusammensetzung im Laufe des Lebens auch je nach Entwicklungsstadium (bei vielen Insekten) oder nach Jahreszeit (viele Singvögel).
Warum ist die Länge der Nahrungsketten limitiert?
Schon der Biologe Charles Elton stellte in den 1920er Jahren fest, dass Nahrungsketten in den meisten Fällen auf eine Länge von vier bis fünf Glieder beschränkt sind, also in der Regel über nur vier bis fünf Stationen (Trophieebenen) reichen.
Für die Erklärung dieser scheinbar konstanten Einschränkung der Nahrungskettenlänge liegen zwei Hypothesen vor:
Der Energiehypothese zufolge ist der Energieverlust von Trophiestufe zu Trophiestufe der einschränkende Faktor. Da die geringe trophische Effizienz, also der zur nächsten Stufe weitergegebene in Biomasse gespeicherte Energiegehalt, nur 10 % beträgt, nimmt die Energie, die jeder Ebenen zur Verfügung steht exponentiell ab. Dadurch ist die Kette in ihrer Länge eingeschränkt. Stehen auf einem kleinen Weidegebiet 100 kg Pflanzenmasse, so reicht dies nur für die Erzeugung von 10 kg Herbivorenbiomasse und 1 kg Carnivorenbiomasse. Erhöht sich die Primärproduktion in einer Nahrungskette, so könnte die Kette, gemäß dieser Hypothese, länger werden, denn die Energie reiche aus, trotz des Verlusts, eine weitere Trophiestufe zu versorgen.
Die dynamische Stabilitätshypothese hingegen schreibt der Stabilität einer Nahrungskette das einschränkende Potential zu. Je länger eine Nahrungskette ist, desto instabiler ist sie auch. Schwankungen in den unteren Stufen verstärken sich nach oben und können letztlich zur Auslöschung der Spitzenräuber führen. Ist eine Nahrungskette jedoch kürzer, so können die höheren Konsumenten leichter auf Fluktuationen, z. B. Umweltkatastrophen, reagieren, die das Nahrungsangebot auf allen Trophiebenen beeinflussen. In einem sehr variablem Nahrungsgefüge sind die Nahrungsketten demzufolge kürzer. [4]
Bedeutung für die Ökotoxikologie
Ins öffentliche Interesse rückte der Begriff in Zusammenhang mit der Beobachtung einer Anreicherung von Schadstoffen bei aufeinander folgenden Gliedern (Nahrungsketten- oder Trophieebenen). Die in diesem Zusammenhang am häufigsten zitierte Arbeit ist diejenige von Woodwell und Mitarbeitern aus dem Jahre 1967[5]. Tatsächlich können sich vor allem fettlösliche und nicht oder nur langsam abbaubare Stoffe (z. B. persistente Chlorkohlenwasserstoffe, Schwermetallionen) in aufeinander folgenden Nahrungskettengliedern unter bestimmten Bedingungen anreichern. Man spricht in diesem Zusammenhang auch von Biomagnifikation.
Dieser Effekt wurde zunächst als allgemein gültig betrachtet, erwies sich aber auf Basis einer Vielzahl an Untersuchungen als insbesondere für luftatmende und/oder terrestrisch lebende Organismen (Vögel, Robben usw.) bedeutsam[6]. Auch der Mensch kann dieser Form der Schadstoffanreicherung unterliegen (z.B. in der Muttermilch). Bei primären Wassertieren (z. B. Wasserschnecken, Wasserflöhe, Fische) in rein aquatischen Nahrungsketten ist der direkte Austausch der Stoffe aus der Wasserphase über die Epithelien der Organismen (der Vorgang der so genannten Biokonzentration in den Organismus hinein und die Elimination über Kiemen oder Hautoberflächen aus dem Organismus heraus) bedeutsamer als die durch Fressen kontaminierter Nahrung hervorgerufene Biomagnifikation[7], wenngleich starke Unterschiede zwischen den verschiedenen Stofftypen und den einzelnen Organismengruppen auftreten.
Eine nähere Analyse ist im Einzelfall nur durch Messung realer Stoffflüsse und durch Anwendung geeigneter Kompartimentmodellierung möglich. Eine wichtige Stoffeigenschaft bei fettlöslichen Stoffen ist dabei z. B. der Oktanol-Wasser-Verteilungskoeffizient; ein wichtiges Charakteristikum der untersuchten Organismen ist die Größe der Fläche sowie die Durchlässigkeit ihrer Membranen bzw. Epithelien, welche den Stoffaustausch mit der Umwelt ermöglichen[8].
Einzelnachweise
- ↑ V. Storch, U. Welsch, M. Wink: Evolutionsbiologie, Springer-Verlag, Berlin Heidelberg, 2. Aufl. 2007
- ↑ Kriwet, J., Witzmann, F., Klug, S. & Heidtke, H.J. (2008): „First direct evidence of a three-level trophic chain in the fossil record.“ Proceedings of the Royal Society B: Biological Sciences 275: 181-186. Vollständiger Artikel (HTML) (Abstract)
- ↑ Fressen und gefressen werden in einem urzeitlichen See - einmalige Einblicke in eine 290 Millionen Jahre alte Nahrungskette (Innovations-report.de, Stand: 6.11.2007)
- ↑ Campbell, Neil A. / Reece, Jane B. (2003): Biologie, Spektrum akademischer Verlag Berlin Heidelberg, 6. Auflage
- ↑ George M. Woodwell, Charles F. Wurster Jr., Peter A. Isaacson: "DDT residues in an East coast estuary: A case of biological concentration of a persistent insecticide." Science 156: 821-824 (1967)
- ↑ Winter, S., Streit, B. (1992): "Organochlorine compounds in a three-step terrestrial food chain". Chemosphere 24: 1765-1774
- ↑ Streit, B. (1979): "Uptake, accumulation and release of organic pesticides by benthic invertebrates. 3. Distribution of 14C-atrazine and 14C-lindane in an experimental 3-step food chain microcosm". Arch. Hydrobiol./ Suppl. 55: 374-400)
- ↑ Streit, B. (1998): "Bioaccumulation of contaminants in fish". S. 353-387 in: Braunbeck, T., Hinton, D.E., Streit, B.: Fish Ecotoxicology. Birkhäuser Publ., Basel etc.

Wikimedia Foundation.