- Nukleosom
-
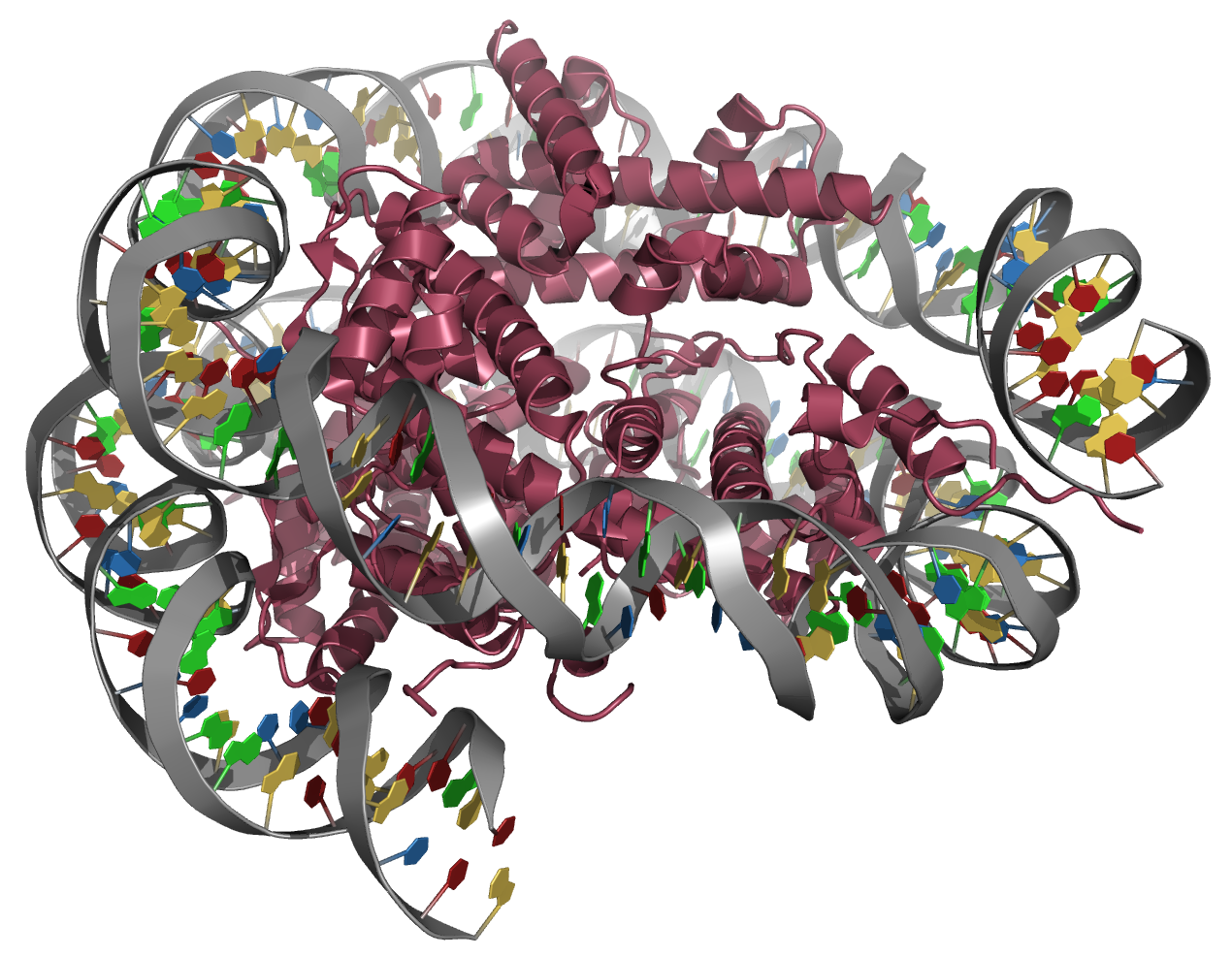
 Struktur eines Nukleosoms mit Histonen der Fruchtfliege: Die DNA (grau mit farbigen Nukleobasen) ist um den Kern aus acht Histon-Untereinheiten (bordeauxrot) gewickelt
Struktur eines Nukleosoms mit Histonen der Fruchtfliege: Die DNA (grau mit farbigen Nukleobasen) ist um den Kern aus acht Histon-Untereinheiten (bordeauxrot) gewickelt
Die erste Verpackungsstufe der DNA höherer Zellen ist ein Komplex aus DNA und Histonen, das so genannte Nukleosom. Von Ada und Donald Olins in elektronenmikroskopischen Darstellungen gequollener Zellkerne entdeckt und 1973 erstmals auf dem "Third Annual Meeting of the American Society for Cell Biology" als "ν-body" (neues Partikel) vorgestellt, wurde dieses nahezu umgehend als fundamentale Verpackungseinheit der DNA im Chromatin akzeptiert.
Inhaltsverzeichnis
Entdeckung und Eigenschaften
1974 gelangen mehreren Teams, darunter jenem von Roger Kornberg, Analysen, die den Aufbau dieser Partikel aus einem Histon-Oktamer, einem Linker-Histon und etwa 160-200 Basenpaaren an DNA zeigten. 1975 wurde diese Einheit als Nukleosom eingeführt. 1974 gilt heute als Geburtsjahr der molekularen Epigenetik.
Neben Wechselwirkungen, die zur Kompaktierung der DNA führen, gehen die Histone Interaktionen untereinander ein. So wird der Nukleosomenkern (das "core particle") aus jeweils zwei Exemplaren der Histone H2a, H2b, H3 und H4 gebildet, um das in 1,65 Windungen 147 Basenpaare an DNA gewickelt sind. Der Bereich zwischen zwei Nukleosomen (der variable "linker", der zwischen 160 Basenpaaren in Hefe und 200 Basenpaaren in höheren Organismen umfassen kann; beim Menschen sind es 50-60 Basenpaare) wird durch ein weiteres Histon, H1, besetzt, welches am Aufbau höherer Strukturen (der sog. 30-nm-Faser, erklärt z. B. im "Solenoid"-Modell) beteiligt ist. Die Komponenten des Nukleosomenkerns wurden in der Evolution hoch konserviert (nur zwei Aminosäurereste unterscheiden das Histon H3 des Menschen von jenem der Erbse), was die fundamentale Bedeutung dieser Einheit (und ihrer Modifikationen - siehe unten) unterstreicht.
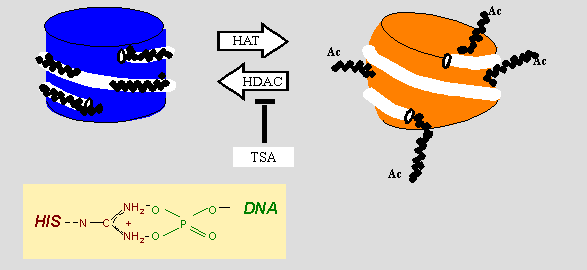
 Nukleosomen und ihre Dynamik: links ist DNA (weißer Schlauch) eng mit dem inaktiven Nukleosomenkern verbunden. Die Kontakte werden über Wasserstoffbrücken und elektrostatische Wechselwirkungen hergestellt (schwarze Punkte; herausvergrößert ist ein Phosphat-Arginin-Kontakt zwischen DNA und Histon, HIS). Histon-Methyltransferasen (HMTs) und Histon-Acetyltransferasen (HATs) sind Enzyme, die bestimmten Aminosäuren der Histone H2a, H2b, H3 und H4 posttranslational modifizieren können; ihre Gegenspieler sind Histon-Deacetylasen (HDACs) und Histon-Demethylasen. Stärkere Acetylierung, und (durch Öffnung der Nucleosomenstruktur) erhöhte Genexpression wird durch HDAC Inhibitoren wie Trichostatin A (TSA) bewirkt. Dieses Wirkprinzip wird mittlerweile in einigen Krebstherapien eingesetzt. In der Zelle ist die Modifizierung definierter Aminosäurereste strikt reguliert und mit analogen Modifikationen der DNA-Basen koordiniert, siehe Epigenetik.
Nukleosomen und ihre Dynamik: links ist DNA (weißer Schlauch) eng mit dem inaktiven Nukleosomenkern verbunden. Die Kontakte werden über Wasserstoffbrücken und elektrostatische Wechselwirkungen hergestellt (schwarze Punkte; herausvergrößert ist ein Phosphat-Arginin-Kontakt zwischen DNA und Histon, HIS). Histon-Methyltransferasen (HMTs) und Histon-Acetyltransferasen (HATs) sind Enzyme, die bestimmten Aminosäuren der Histone H2a, H2b, H3 und H4 posttranslational modifizieren können; ihre Gegenspieler sind Histon-Deacetylasen (HDACs) und Histon-Demethylasen. Stärkere Acetylierung, und (durch Öffnung der Nucleosomenstruktur) erhöhte Genexpression wird durch HDAC Inhibitoren wie Trichostatin A (TSA) bewirkt. Dieses Wirkprinzip wird mittlerweile in einigen Krebstherapien eingesetzt. In der Zelle ist die Modifizierung definierter Aminosäurereste strikt reguliert und mit analogen Modifikationen der DNA-Basen koordiniert, siehe Epigenetik.Die Abbildung gibt den schematischen Aufbau des Nukleosoms und seine strukturelle Variabilität wieder. Der Histonkern bildet einen Zylinder von 11 nm Durchmesser und 5 nm Höhe (blau). Die überwiegend Lysin- und Arginin-reichen und damit positiv geladenen N-terminalen Enden der Histone (die "histone-tails") binden an das negativ geladene Rückgrat der DNA-Phosphatreste, wodurch die Zugänglichkeit für Transkriptionsfaktoren eingeschränkt wird. Acetylierung der Lysinreste durch Acetyltransferasen beseitigt deren positive Ladung und gibt die zuvor verdeckte Information frei; gleichzeitig erfährt der Nukleosomenkern eine Strukturänderung (rot). Dieses stark vereinfachende Ladungsmodell ist mittlerweile zumindest umstritten. Neuere Forschungen zeigen, dass die modifizierten Motive durch verschiedene Proteine ausgelesen werden können, und dies zur Aktivierung oder Reprimierung der betroffenen Gene führt. Der Acetylierungsstatus selbst wird durch das Gleichgewicht der Histon-Acetyltransferasen (HATs) bzw. Histon-Deacetylasen (HDACs) bestimmt - werden letztere durch einen spezifische Inhibitoren wie z.B.: Trichostatin A (TSA) oder Butyrat gehemmt, so dominieren die ersteren. Derartige Versuchsansätze haben zum experimentellen Nachweis dieses Sachverhaltes geführt.
Arbeiten zur Struktur der Nukleosomen wurden durch Aaron Klug (Nobelpreis 1982[1] für Kristall-Strukturanalysen an Protein/Nucleinsäure-Komplexen) in London am Medical Research Council aufgenommen. Diese Arbeiten führten 1984 bei noch relativ geringer Auflösung zum ersten Strukturvorschlag.[2][3] Die Arbeiten werden seitdem systematisch durch Timothy Richmond[4], der bereits in der Gruppe von Klug 1984 an dem ersten Strukturvorschlag beteiligt war, am Institute for Molecular Biology & Biophysics der ETH Zürich vorangetrieben. 1997 publizierte die Arbeitsgruppe von Richmond eine Struktur des Nukleosoms mit einer Auflösung von 2.8 Å[5] und 2002 folgte die Publikation der Struktur mit einer Auflösung von 1.9 Å.[6][7]
Im Jahr 2005 publizierte die Arbeitsgruppe von Richmond eine Röntgen-Kristallstruktur des Tetranukleosoms.[8]
Einzelnachweise
- ↑ http://www.nobel.se/chemistry/laureates/1982/klug-autobio.html Aaron Klug Nobelpreis]
- ↑ Richmond, T.J., Finch, J.T., Rushton, B., Rhodes, D., and Klug, A. (1984): Structure of the nucleosome core particle at 7 Å resolution. In: Nature. 311, 532-537. PMID 6482966
- ↑ Struck, M. M., Klug, A., and Richmond, T.J. (1992): Comparison of X-ray structures of the nucleosome core particle in two different hydration states. In: J. Mol. Biol. 224, 253-264. PMID 1548703
- ↑ Gruppe T.J. Richmond
- ↑ Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ (1997): Crystal Structure of the Nucleosome Core Particle at 2.8 Å Resolution. In: Nature. 389 (6648): 251-260. PMID 9305837
- ↑ Davey CA, Sargent DF, Luger K, Maeder AW, Richmond TJ (2002): Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 Å resolution. In: J. Mol. Biol. 319 (5): 1097-1113. PMID 12079350
- ↑ Richmond T.J., Davey CA (2003): The structure of DNA in the nucleosome core. In: Nature. 423, 145-150. PMID 12736678
- ↑ Schalch T, Duda S, Sargent DF, Richmond TJ (2005): X-ray structure of a tetranucleosome and its implications for the chromatin fibre. In: Nature. 436(7047):138-141. PMID 16001076
Literatur
- Prior, C.P., Cantor, C.R., Johnson, E.M., Littau, V.C., Allfrey, V.G. (1983): Reversible changes in nucleosome structure and histone H3 accessibility in transcriptionally active and inactive states of rDNA chromatin. In: Cell. 34, 1033-1042.
- Bode, J.Gòmez-Lira M.M., Schröter, H. (1983): Nucleosomal Particles Open as the Histone Core Becomes Hyperacetylated. In: Eur. J. Biochem. 130, 437-445.
- Strahl, B.D., Allis, C.D. (2000): The language of covalent histone modifications. In: Nature. 403(6765): 41-45.
- Teif, V.B., Rippe K., (2009). Predicting nucleosome positions on the DNA: combining intrinsic affinities and remodeler activities. In Nucleic Acids Res. 37, 5641-5655. http://nar.oxfordjournals.org/content/37/17/5641
Weblinks
Kategorien:- Zellbestandteil
- Zellbiologie
- Chromatin
- Epigenetik
Wikimedia Foundation.