- Histon
-
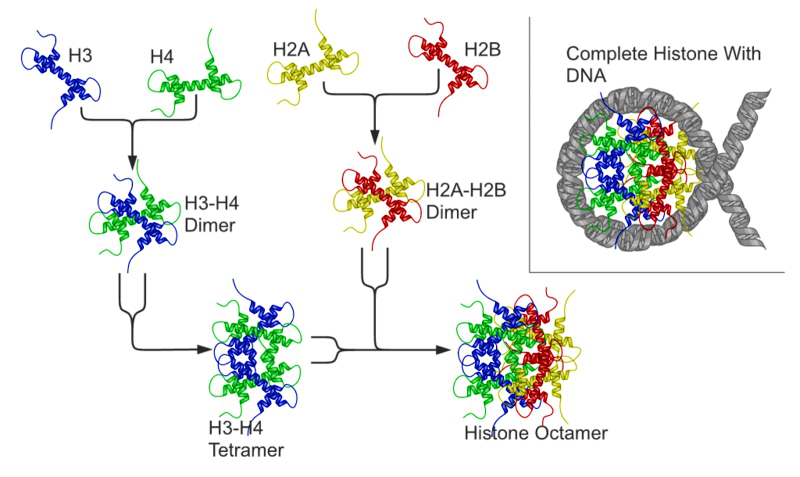
 Schematische Darstellung der Nukleosombildung aus den Histonproteinen.
Schematische Darstellung der Nukleosombildung aus den Histonproteinen.
Histone sind basische Proteine, die im Zellkern von Eukaryoten vorkommen. Sie sind als Bestandteil des Chromatins für die Verpackung der DNA (es sind Spulen, um welche sich die DNA windet), aber auch für die Expression mancher auf ihr codierten Gene von essentieller Bedeutung. Um das große Genom der Eukaryoten im Zellkern unterbringen zu können, ist es nötig, die DNA besonders zu verpacken. Bei Eukaryoten geschieht dies in Chromosomen, deren kleinste Verpackungseinheit ein Nukleosom ist. Ein Nukleosom besteht aus acht Histonen (lat. Oktamer), aus je zwei Kopien der Histone H2A, H2B, H3 und H4. Die DNA ist 1,65x um ein Nukleosom gewickelt, was einer DNA-Länge von 146 Basenpaaren entspricht. Die nicht assoziierte DNA, die benachbarte Nukleosomen verbindet, wird "Linker-DNA" genannt. Das Histon H1 bindet DNA direkt neben Nukleosomen und erlaubt die nächsthöhere Verpackungseinheit der DNA.
Histone bestehen aus einem globulären Zentrum und flexiblen endständigen Armen (engl. histone tails), die viele basische, also positiv geladene, Aminosäuren besitzen. Die DNA ist hingegen negativ geladen, so dass eine elektrostatische Anziehung besteht.
Inhaltsverzeichnis
Entdeckung
Die Histon-Proteine wurden 1884 vom deutschen Mediziner und Physiologen Albrecht Kossel entdeckt. Der Begriff Histon stammt aus der deutschen Sprache. Unklar ist, ob sich der deutsche Begriff vom griechischen histanai oder histos herleitet. Bis in die frühen 90er Jahre wurden Histone als reines Packmaterial nukleärer DNA verkannt. Erst in den letzten beiden Jahrzehnten konnte ihre Bedeutung für epigenetische Mechanismen beschrieben werden.[1]
Histonklassen
Es sind fünf Haupt-Histon-Proteine bekannt:
Die Histonproteine H2A und H2B lagern sich zu Dimeren zusammen. Dasselbe gilt für H3 und H4. Zwei H3/H4-Dimere lagern sich zu einem Tetramer zusammen, an das wiederum zwei H2A/H2B-Dimere angelagert werden. Dadurch entsteht der oktamere Nukleosomenkern (´core particle´), um den sich die DNA in ca. zwei großen linksgängigen Windungen legen kann. Das fünfte Histon, H1, wird möglicherweise benötigt, um eine 30 nm-Faser zu bilden - eine übergeordnete Struktur, die einer Helix aus Nukleosomen entspricht. Dadurch wird die DNA Packung weiter verstärkt. Die Komprimierung des DNA-Moleküls beträgt ohne H1 Faktor 7 und wird mit H1 auf Faktor 40-50 erhöht, d. h. ein unkomprimierter DNA-Strang enthält 3 Mio. Nukleotide/mm, komprimiert ohne H1 20 Mio. Nukleotide/mm und mit H1 120 Mio. Nukleotide/mm.
Neben den oben genannten Histonproteinen gibt es außerdem Varianten, die sehr spezifische Funktionen bei der Regulation der Genexpression und der Strukturierung der Chromosomen übernehmen. Ein Beispiel ist Makro-H2A, welches das Histon H2A partiell auf dem inaktivierten X-Chromosom von weiblichen Säugern ersetzt. Ein anderes Beispiel ist CENP-A, eine Variante des Histons H3, die nur im Bereich des Zentromers zu finden ist und für die spezifische Struktur dieser Chromosomenregion essentiell ist. Im Großen und Ganzen sind die Core-Histone H2A, H2B, H3 und H4 in der Evolution streng konserviert worden. Lediglich das Histon H1 ist in seiner Struktur sehr variabel, wie der Vergleich unterschiedlicher Organismen zeigt. Bei der Bäckerhefe (Saccharomyces cerevisiae) fehlt dieses Histon sogar völlig. Die Bindung der DNA an die Histone kann die Transkription positiv oder negativ beeinflussen. Für die Vorgänge der Transkription, Replikation und DNA-Reparatur müssen die Histone von der DNA gelöst oder auf dem DNA-Strang verschoben verden - ein Vorgang, den man als Nukleosomenremodelling bezeichnet.[2]
Histonmodifikationen
Das N-terminale Ende eines Histons kann von Enzymen modifiziert werden. Diese Histonmodifikationen können Methylierung, Phosphorylierung, Sumoylierung, Ubiquitinylierung und Acetylierung sowie deren Rückreaktionen umfassen. Hieraus ergibt sich der spezifische Histon-Code einer Zelle. Diese Modifikationen haben Einfluss auf das Chromatingerüst des Zellkerns und somit auf die Genregulation.[3][4][5][6][7][8]
Die Funktion der Methylierung von Histonen wird derzeit intensiv erforscht und steht überwiegend in Beziehung zur epigenetischen Inaktivierung von Genen. So kann eine regionale Trimethylierung des Lysinseitenrestes (K9) am Histon 3 eines Promoters, zu einer Kondensierung der Chromatinstruktur in diesem Bereich führen, dies hat dann eine Inaktivierung der Genexpression des auf diesem Abschnitt liegenden Gens zur Folge. Darüber hinaus existieren Verbindungen zum inaktivierenden Prozess der DNA-Methylierung. Die Phosphorylierung von Histonproteinen erhöht in den meisten Fällen die Zugänglichkeit der DNA und spielt unter anderem eine wichtige Rolle bei der Regulation der Transkription während der Mitose und Meiose. Die Acetylierung setzt, wie die Phosphorylierung, in den meisten Fällen die Bindefähigkeit der Histone für die DNA herab, indem die Ladung der Histone negativ wird und sich die negative DNA abstößt. Sie ist deshalb Voraussetzung für die Transkription der mit Histonen assoziierten DNA.[9][10]
Histongene
Histone sind offensichtlich evolutionär sehr alte und sehr bedeutende Moleküle, sodass sie hoch konserviert sind. Histongene sind S-Phase-abhängig, werden also nur exprimiert, wenn neue DNA gebildet wird, und zwar dann extrem stark.
Die Histongene liegen ähnlich wie die rDNA häufig in Clustern wiederholt vor, sind also gekoppelt, sie besitzen jedoch jeweils einen eigenen Promotor. Die Zahl der Wiederholungen kann recht stark schwanken (Saccharomyces cerevisiae hat 2 Cluster, der Grüne Wassermolch hat 700, beim Menschen sind es 10-24).
Die Reihenfolge der einzelnen Histongene im Cluster ist recht unterschiedlich, teilweise sind die Gene zudem gegenläufig, teilweise auch in gleicher Richtung angeordnet. In jedem Cluster jedoch sind alle 5 Histone vorhanden.
Die Histongene selbst haben zudem eine regelrecht altertümliche Struktur. Sie besitzen weder Introns, noch erhalten sie nach der Transkription einen poly(A)-Schwanz. Der 3'-UTR ist sehr kurz und enthält zwei gegenläufig gerichtete Wiederholungseinheiten (inverted repeats). Diese bilden eine Haarnadelstruktur aus, was man ansonsten nur von Genen von Prokaryoten kennt. Die Haarnadelstruktur ist für die koordinierte Reifung der Histon-mRNA und für die Regelung ihrer Lebenszeit in der S-Phase wichtig.Bei Säugetieren und Vögeln sind die Cluster nicht repetitiv angeordnet, sondern etwas verteilt. Einige Histongene sind S-Phase-korreliert und liegen recht nahe beieinander. Sie entsprechen vom Aufbau her den "normalen" Histongenen. Andere hingegen sind sogenannte Ersatzhistongene, die defekte Histonproteine ersetzen sollen, auch außerhalb der S-Phase. Diese liegen abseits der Histoncluster und besitzen lange 3'-UTRs und einen poly(A)-Schwanz.
Siehe auch
Zur Rolle von Histonmodifikationen bei der Regulation des Genoms siehe auch Epigenetik.
Zum Komplex, den die Histone mit der DNA bilden, siehe auch Nukleosom.
Weitere verwandte Themen:Literatur
- Histone modification and replacement in chromatin activation. Genes Dev 16(14):1739-42 (2002) PMID 12130533
- A tale of histone modifications. Genome Biol 2(4):REVIEWS0003 (2001) PMID 11305943
- Protamines, in the footsteps of linker histone evolution. J Biol Chem 281(1):1-4 (2006) PMID 16243843
- Monitoring gene expression in Chinese hamster ovary cells using secreted apoaequorin. Anal Biochem 201(1):114-8 (1992) PMID 1621948
- Functional reorganization of the brain in recovery from striatocapsular infarction in man. Ann Neurol 31(5):463-72 (1992) PMID 1596081
- Tails of intrigue: phosphorylation of RNA polymerase II mediates histone methylation. Cell 113(4):429-32 (2003) PMID 12757703
- Cellular memory and the histone code. Cell 111(3):285-91 (2002) PMID 12419240
- Histone methylation: dynamic or static? Cell 109(7):801-6 (2002) PMID 12110177
- Signaling to chromatin through histone modifications. Cell 103(2):263-71 (2000) PMID 11057899
- Histones and histone modifications. Curr Biol 14(14):R546-51 (2004) PMID 15268870
- Linker histones: paradigm lost but questions remain. FEBS Lett 441(2):161-4 (1998) PMID 9883876
- More histone structures. FEBS Lett 100(2):201-18 (1979) PMID 378692
- Dynamic regulation of histone lysine methylation by demethylases. Mol Cell 25(1):1-14 (2007) PMID 17218267
Einzelnachweise
- ↑ Informationen der Nobelstiftung zur Preisverleihung 1910 an Albrecht Kossel (englisch)
- ↑ Kamakaka RT, Biggins S: Histone variants: deviants?. In: Genes Dev.. 19, Nr. 3, Februar 2005, S. 295–310. doi:10.1101/gad.1272805. PMID 15687254.
- ↑ Bártová E, Krejcí J, Harnicarová A, Galiová G, Kozubek S: Histone modifications and nuclear architecture: a review. In: J. Histochem. Cytochem.. 56, Nr. 8, August 2008, S. 711–21. doi:10.1369/jhc.2008.951251. PMID 18474937. Volltext bei PMC: 2443610.
- ↑ Eberharter A, Becker PB: Histone acetylation: a switch between repressive and permissive chromatin. Second in review series on chromatin dynamics. In: EMBO Rep.. 3, Nr. 3, März 2002, S. 224–9. doi:10.1093/embo-reports/kvf053. PMID 11882541. Volltext bei PMC: 1084017.
- ↑ Zhang Y: Transcriptional regulation by histone ubiquitination and deubiquitination. In: Genes Dev.. 17, Nr. 22, November 2003, S. 2733–40. doi:10.1101/gad.1156403. PMID 14630937.
- ↑ Mersfelder EL, Parthun MR: The tale beyond the tail: histone core domain modifications and the regulation of chromatin structure. In: Nucleic Acids Res.. 34, Nr. 9, 2006, S. 2653–62. doi:10.1093/nar/gkl338. PMID 16714444. Volltext bei PMC: 1464108.
- ↑ Suganuma T, Workman JL: Crosstalk among Histone Modifications. In: Cell. 135, Nr. 4, November 2008, S. 604–7. doi:10.1016/j.cell.2008.10.036. PMID 19013272.
- ↑ Weake VM, Workman JL: Histone ubiquitination: triggering gene activity. In: Mol. Cell. 29, Nr. 6, März 2008, S. 653–63. doi:10.1016/j.molcel.2008.02.014. PMID 18374642.
- ↑ Cloos PA, Christensen J, Agger K, Helin K: Erasing the methyl mark: histone demethylases at the center of cellular differentiation and disease. In: Genes Dev.. 22, Nr. 9, Mai 2008, S. 1115–40. doi:10.1101/gad.1652908. PMID 18451103. Volltext bei PMC: 2732404.
- ↑ Zhang Y, Reinberg D: Transcription regulation by histone methylation: interplay between different covalent modifications of the core histone tails. In: Genes Dev.. 15, Nr. 18, September 2001, S. 2343–60. doi:10.1101/gad.927301. PMID 11562345.
Wikimedia Foundation.