- Endosymbionten-Theorie
-
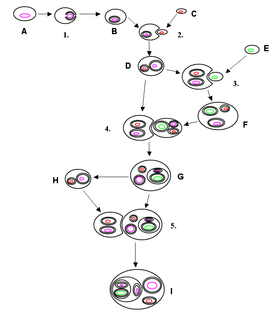 Darstellung der Endosymbiontentheorie
Darstellung der EndosymbiontentheorieDie Endosymbiontentheorie (griech. ἔνδον endo = innen, συμβίωσις symbiōsis = das Zusammenleben) besagt, dass in einer frühen Phase der Evolution der Eukaryoten (also aller höheren Organismen) prokaryotische Zellen (d.h. Bakterien und mikroskopische Algen) durch Endozytose in voreukaryotische Urzellen aufgenommen wurden und in diesen den Status von Organellen erlangt haben.
Inhaltsverzeichnis
Geschichte
Der Gedanke der Endosymbiontentheorie wurde erstmals von dem Botaniker Andreas Franz Wilhelm Schimper im Jahr 1883 veröffentlicht, der damit die Entstehung der Chloroplasten zu erklären versuchte. Die Hypothese wurde erneut um 1905 von dem russischen Evolutionsbiologen Konstantin Sergejewitsch Mereschkowski aufgegriffen. Doch erst 1967 mit der Veröffentlichung von Lynn Margulis wurde sie bekannter.[1]
Kernaussage
Vereinfacht gesprochen besagt die Theorie, dass im Laufe der Entwicklung des Lebens die Zelle eines einzelligen Lebewesens durch die Zelle eines anderen einzelligen Lebewesens „geschluckt“ wurde und zu einem Bestandteil der Zelle eines so entstandenen höheren Lebewesens wurde. Im Laufe der Evolution entstanden so immer komplexere Lebewesen. Auch Bestandteile menschlicher Zellen gehen ursprünglich auf einzellige Lebewesen zurück, die zu einem Bestandteil der Zellen wurden.
Erläuterung
Die Endosymbiontentheorie geht davon aus, dass Mitochondrien und Plastiden sich aus eigenständigen prokaryotischen Lebewesen entwickelt haben. Im Zuge des Evolutionsprozesses sind diese Einzeller eine Endosymbiose mit einer eukaryotischen Zelle eingegangen, das heißt, sie leben in ihrer Wirtszelle zum gegenseitigen Vorteil. Auch heute noch kann man beobachten, dass amöboide Einzeller (also solche mit einer „weichen“ Membran) Cyanobakterien aufnehmen, ohne sie zu verdauen.
Das Zusammenspiel der beiden zellulären Organismen hat sich dann im Verlauf der Evolution zu einer gegenseitigen Abhängigkeit entwickelt, in der keiner der beiden Partner mehr ohne den anderen überleben konnte, das heißt, es entstand eine Symbiose. Diese wird Endosymbiose genannt. Die Abhängigkeit geht so weit, dass die Organellen Teile ihres (nicht mehr benötigten) genetischen Materials verloren oder die entsprechenden Gene teilweise in das Kern-Genom integriert wurden. Einzelne Protein-Komplexe in den Organellen, wie z.B. die ATP-Synthase, werden so zum Teil aus kernkodierten, zum Teil aus mitochondrial kodierten Untereinheiten zusammengesetzt.
Analysen der Genome deuten darauf hin, dass Plastiden von Cyanobakterien und Mitochondrien von aeroben Proteobakterien abstammen. Diese Form der Endosymbiose zwischen einem Eukaryoten und einem Prokaryoten wird als primäre Endosymbiose bezeichnet. Entstand das Zellorganell durch die Aufnahme eines Eukaryoten, der bereits ein primäres Endosymbioseereignis erfahren hat, wird dies als sekundäre Endosymbiose bezeichnet.
Primäre Plastiden sind von zwei Hüllmembranen umgeben, die beide prokaryotischen Ursprungs sind. So gibt es drei Typen von primären Plastiden und somit drei Linien von autotrophen Organismen:
- die einzelligen Algen der Glaucocystaceae besitzen Plastiden, die dem Cyanobakterium in vielerlei Hinsicht noch sehr ähnlich sind und aus diesem Grund oft als "Cyanellen" bezeichnet werden,
- Rotalgen besitzen "Rhodoplasten" genannte Plastiden, die noch den Antennenaufbau der Cyanobakterien tragen (Phycobilisomen).
- Die Plastiden der Grünalgen und höheren Pflanzen stellen die am stärksten entwickelten Plastiden dar und tragen eine große Vielfalt an Antennenkomplexen. Die grünen Plastiden der Algen und höheren Pflanzen werden Chloroplasten genannt.
Sekundäre Plastiden verfügen über drei oder sogar vier Hüllmembranen. Es ist kein Fall bekannt, in dem eine Aufnahme eines Glaucophyten zu einer sekundären Endosymbiose geführt hätte. Dagegen existiert eine Fülle von Organismengruppen, die eine Rotalge aufgenommen haben und sie in unterschiedlichem Maße reduziert haben. Einige Autoren gehen davon aus, dass diese Ereignis nur einmal in der Evolution stattgefunden hat und definieren so das Monophylum der Chromalveolata. In diese Gruppe gehören die Braunalgen, Gelbgrünalgen, Goldalgen, Cryptophyceen, Chlorarachniophyceen, Haptophyceen, Euglenozoa und die Apicomplexa (z. B. Malaria-Erreger Plasmodium).
Auch die sekundäre Endosymbiose zwischen Grünalgen und Eukaryoten ist bekannt. So wird angenommen, dass die Euglenozoa und die Chlorarachniophyta unabhängig voneinander primäre Endosymbionten in sich aufgenommen haben.
 Die Endosymbiontentheorie - bezogen auf autotrophe Organismen und die Entstehung der unterschiedlichen Pigmentsysteme.
Die Endosymbiontentheorie - bezogen auf autotrophe Organismen und die Entstehung der unterschiedlichen Pigmentsysteme.Es gibt einige Protozoen, die keine Mitochondrien (und keine Plastiden) besitzen ("Archezoa"). Zunächst wurde angenommen, sie seien primitiv und unmittelbar aus der urtümlichen Wirtszelle der Endosymbionten hervorgegangen. Dies ist vermutlich falsch, da ihre DNA Sequenzen enthält, die eindeutig mitochondrialen Ursprungs sind. Wahrscheinlich haben alle amitochondriaten Eukaryoten ihre Mitochondrien sekundär verloren.
Indizien
- Man kann heute bei unterschiedlichen Lebewesen verschiedene Stadien zwischen Symbiose und Endosymbiose beobachten:
- Korallen, einige Muscheln, der Wurm Convoluta roscoffensis aber zum Beispiel auch Blattläuse leben in Symbiose mit Algen oder Bakterien, die im Zellinneren ihrer Wirte leben. Bei den endosymbiotischen Bakterien der Blattläuse werden Beschleunigungen der Evolutionsraten einhergehend mit Genverlusten und einem Anstieg des AT-Gehaltes der DNA beobachtet, wie sie auch bei Zellorganellen zu finden ist.
- Die Wurzeln einiger Pflanzen leben in Symbiose mit stickstofffixierenden Bakterien (Rhizobien).
- Der Süßwasserpolyp (Hydra viridissima) kann durch Endocytose Zoochlorellen aufnehmen und mit deren Hilfe Photosynthese betreiben.
- Bei Dinoflagellaten sind verschiedene Stadien zu finden: Kleptoplastiden, komplexe Rhodoplasten und tertiäre Endosymbiosen, die auf die Aufnahme von Haptophyceen oder Cryptophyceen zurückzuführen sind.
- Plastiden und Mitochondrien sind von ihrem Aufbau her Prokaryonten: kein Zellkern, ringförmige DNA, die DNA ist nicht durch Histone assoziiert, Größe entspricht kleinen Bakterien. Sie stellen ihre eigenen Proteine her, wobei sie einen prokaryotischen Proteinbiosyntheseapparat besitzen. Ihre Ribosomen ähneln denen der Bakterien, nicht denen der Wirtszelle (70-S- anstatt 80-S-Ribosomen). Die mRNA der beiden Organellen besitzt nicht die für Eukaryoten typische 5'-Cap-Sequenz und auch die Polyadenylierung fehlt. Die Cyanellen der Glaucocystophyceen sind sogar noch von einer dünnen bakteriellen Zellwand umgeben. Rotalgen und Glaucocystophyceen setzen wie Cyanobakterien Phycobiline zum Auffangen von Photonen in der Photosynthese ein.
- Die DNA-Sequenzen der Mitochondrien ähneln denen der α-Proteobakterien, während Plastiden-DNA-Sequenzen mitten im Cyanobakterien-Stammbaum platziert werden. Ein Vergleich mit der Wirts-DNA weist auf keine Abstammung der Organellen vom Wirt hin.
- Primäre Plastiden und Mitochondrien sind von Doppelmembranen umgeben, wobei, der Hypothese entsprechend, die äußere beim "Verschlucken" des Bakteriums hinzugekommen ist. Die innere entspricht der von Bakterien (Vorkommen von Cardiolipin, kein Cholesterin; außerdem Vorkommen von Transmembranproteinen (β-barrel-Proteine), die nur in den Membranen von Bakterien und Zellorganellen vorkommen), die äußere der von Eukaryoten.
- Die besten Belege für sekundäre Endosymbiosen finden sich bei den Chlorarachniophyceen, zu den Cercozoa gehörende Amöben, und den Cryptophyceen, einer eigenständigen Algenklasse. Beide Algengruppen enthalten komplexe Plastiden mit vier Hüllmembranen. Zwischen den beiden äußeren und den beiden inneren Hüllmembranen befindet sich der periplastidäre Raum mit einem Nucleomorph, einem stark reduzierten eukaryotischen Zellkern mit je drei linearen kleinen Chromosomen und eukaryotischen 80-S-Ribosomen. Genomsequenzierungen und phylogenetische Analysen zeigten, dass Nucleomorph und Plastid der Chlorarachniophyceen auf eine sekundäre Endosymbiose mit einer Grünalge, der komplexe Plastid der Cryptophyceen jedoch auf eine sekundäre Endosymbiose mit einer Rotalge zurückzuführen sind. Von der Chlorarachniophycee Bigelowiella natans und von der Cryptophycee Guillardia theta wird/wurde das Nucleomorph-Genom vollständig durchsequenziert. Da bei Rotalgen die Stärkesynthese im Cytoplasma stattfindet und nicht wie bei den Grünalgen und Landpflanzen im Plastiden, spricht das Vorkommen von Stärke im periplastidären Raum der Cryptophyceen ebenfalls für eine sekundäre Endosymbiose.
- Mitochondrien und Plastiden vermehren sich durch Teilung und werden bei Teilung der Wirtszelle auf die Tochterzellen verteilt. Sie entstehen nicht de novo, d.h. sie können von der Zelle bei zufälligem Verlust auch nicht neu gebildet werden.
Quellen
- ↑ Lynn Sagan (1967). On the origin of mitosing cells. In: J. Theoretical Biology. Bd. 14, Nr. 3, S. 255-274. PMID 11541392 doi:10.1016/0022-5193(67)90079-3
Literatur
- Schimper A.F.W. (1883): Über die Entwicklung der Chlorophyllkörner und Farbkörper. In: Bot. Z. 41, 102-113.
- Mereschkowsky C. (1905): Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. In: Biol. Centralb. 25: 593-604
- Margulis L, Sagan D (1997 , engl.1995). Leben: Vom Ursprung zur Vielfalt. Spektrum Verlag
- Lynn Margulis (1999, engl.1998). Die andere Evolution. Spektrum Verlag
- Archibald JM, Rogers MB, Toop M, Ishida K-i, Keeling PJ (2003). Lateral gene transfer and the evolution of plastid-targeted proteins in the secondary plastid-containing alga Bigelowiella natans. Proceedings of the National Academy of Sciences of the USA 100: 7678-7683
- Douglas SE, Zauner S, Fraunholz M, Beaton M, Penny S, Deng LT, Wu X, Reith M, Cavalier-Smith T, Maier U-G (2001). The highly reduced genome of an enslaved algal nucleus. Nature (London) 410: 1040-1041
- Linne von Berg K-H, Hoef-Emden K, Marin B, Melkonian M (2004). Der Kosmos-Algenführer. Die wichtigsten Süßwasseralgen im Mikroskop. Kosmos, Stuttgart. ISBN 3-440-09719-6
- McFadden GI (2001). Primary and secondary endosymbiosis and the origin of plastids. Journal of Phycology 37: 951-959
- Gould SB, Waller RF, McFadden GI (2008) Plastid Evolution. Annu Rev Plant Biol Vol.59: 491-517
- Moran NA (1996). Accelerated evolution and Muller's ratchet in endosymbiotic bacteria. Proceedings of the National Academy of Sciences of the USA 93: 2873-2878
- Turner S, Pryer KM, Miao VPW, Palmer JD (1999). Investigating deep phylogenetic relationships among Cyanobacteria and plastids by small subunit rRNA sequence analysis. Journal of Eukaryotic Microbiology 46: 327-338
- Witzany G (2006). Serial Endosymbiotic Theory (SET): The Biosemiotic Update. Acta Biotheoretica 54: 103-117
Siehe auch
Weblinks
- www.wissenschaft.de: Japanische Wissenschaftler beobachten die Entstehung einer neuen Pflanze Erstmals ist es den japanischen Wissenschaftlern gelungen, den Verlauf der Endosymbiose live zu beobachten.
- FR Biologie Hannover: Die Endosymbiontentheorie inklusive einer Erklärung zur Hydrogenhypothese als Alternative zur Endosymbiontentheorie.

Wikimedia Foundation.