- Kallose
-
Callose (auch Kallose) ist ein Polysaccharid, das pflanzlichen Zellen als universelles Abdichtungsmaterial in verschiedensten Situationen dient.
 Calloseablagerungen in Schwammparenchymzellen von Tabak
Calloseablagerungen in Schwammparenchymzellen von TabakInhaltsverzeichnis
Aufbau
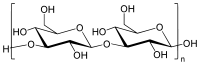 Ein Callosedimer
Ein CallosedimerBei Callose handelt es sich um ein pflanzliches Polysaccharid. β-D-Glucose-Monomere sind hierbei über β-1→3-glycosidische Bindungen zu einem Polymer bzw. Glucan verknüpft, welches teilweise auch β-1,6-Verzweigungen aufweisen kann. Durch diese Form der Verknüpfung ergibt sich eine lange, schraubenförmige Kette des Glucans, das stets in kompakter Form vorliegt.
Ein Nachweis von Callose im pflanzlichen Gewebe kann über Anfärben mit dem Farbstoff Anilinblau erfolgen. Dieser lagert sich in die helikale Struktur der Callose ein und ist unter UV-Licht durch Fluoreszenz nachweisbar.
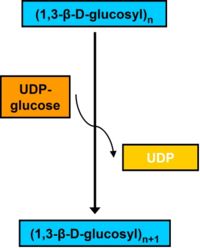 Callosesynthese durch die CalS
Callosesynthese durch die CalSSynthese
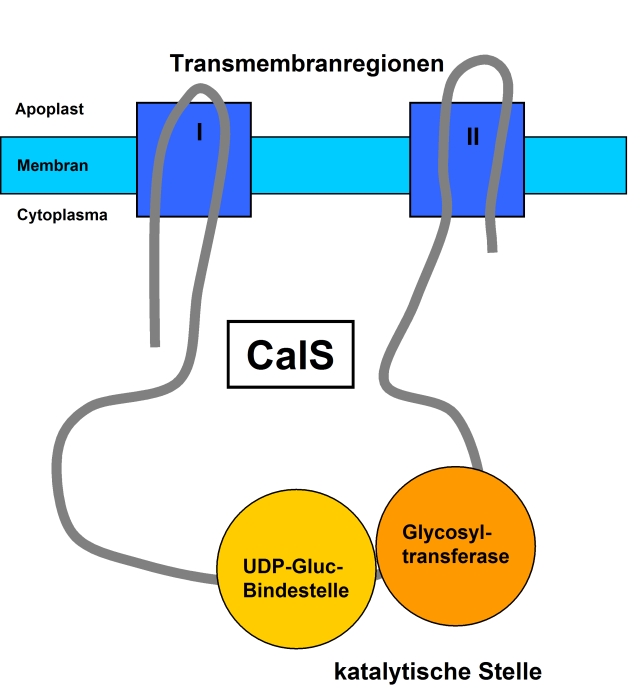
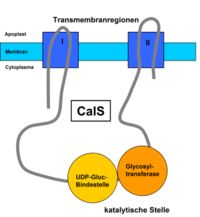 vereinfachter Aufbau der CalS
vereinfachter Aufbau der CalSDie Synthese der Callose erfolgt entgegen der ursprünglichen Vermutung nicht durch die rosettenförmige Cellulosesynthase, sondern durch eine ebenfalls membranständige Callosesynthase (CalS) (EC 2.4.1.34). Die CalS katalysiert eine sogenannte vektorielle Reaktion, bei der das Substrat (UDP-Glucose) an der cytoplasmatischen Seite der Plasmamembran gebunden und nach Verknüpfung an der Außenseite der Zellmembran in den Apoplasten, also zwischen Membran und Zellwand abgelagert wird.
CalS wird durch eine Familie Glucansynthase-ähnlicher Gene (engl. gluccan-synthase like: GSL) codiert, die eine starke Homologität zu in die Glucanbiosynthese von Hefen involvierten Genen aufweist. Die primäre Sequenz der katalytischen Untereinheiten von CalS (2000 Aminosäuren) und Cellulosesynthase (1000 Aminosäuren) sind ein eindeutiger Hinweis auf die separat stattfindende Synthese der Callose. Bei Arabidopsis thaliana konnten bisher zwölf solcher, die Untereinheiten verschlüsselnde GSL-Gene identifiziert werden, die CalS1 bis CalS12 benannt wurden. Bei Arabidopsis liegen diese zwölf Gene über die gesamten fünf vorhandenen Chromosomen verteilt vor.
Die CalS liegt als Protein mit mehreren Transmembrandomänen vor, welche in zwei Regionen zusammengefasst bestehen. Diese beiden Regionen sind zum Cytoplasma hin über eine hydrophile Schleife verbunden, die die katalytische Untereinheit, z. B. CalS1 enthält. Die katalytische Stelle lässt sich in UDP-Glucose-bindende- und Glycosyltransferase-Domäne unterteilen. Mit dieser hydrophilen Schleife assoziiert sind außerdem noch verschiedene weitere Proteine, die der Unterstützung der Synthese und der Regulation des Enzymkomplexes dienen. Eine entscheidende Rolle spielt die Saccharose-Synthase, die durch Spaltung von Saccharose UDP-Glucose generiert. Des Weiteren sind laut einer Hypothese hier auch ein Annexin-ähnliches Membranprotein, ein Rop-Protein, eine Prolin-reiche Domäne, eine Glykosylierungsstelle und zwei Phosphorylierungsstellen zu finden.
Funktionen
Callose dient pflanzlichen Zellen als universelles Abdichtungsmaterial und kommt sowohl in regulären Entwicklungs- und Wachstumsprozessen als auch als Reaktion auf biotische und abiotische Stressfaktoren zum Einsatz. Besteht für die Zelle Bedarf, kann Callose schnell in großen Mengen synthetisiert werden, auf der anderen Seite ist auch ein rascher Abbau möglich.
Typische Funktionen der Callose „im regulären Betrieb“ der Pflanze sind z. B. das Auftreten in der Zellplatte der sich teilenden Zelle während der Cytokinese. In Pollenmutterzellen stellt die Callose die Hauptkomponente dar. Auch in wachsenden Pollenschläuchen versiegelt der vorrückende Protoplast den verlassenen Abschnitt des Schlauchs mit Callosepfropfen. Callose verschließt auch die Poren der Siebplatten von ruhendem Phloem (Winterruhe) bzw. von defektem Phloem und trennt so effizient vom übrigen Gewebe ab. In Plasmodesmata (Zell-Zell-Verbindung) stellt die Callose einen strukturellen Bestandteil dar, der durch einen Mantel um die Halsregion des Kanals dessen Durchmesser regulieren und so den interzellulären Transport einschränken kann.
Der Verschluss von Siebporen und daneben auch die Regulation der Größe von Plasmodesmata sind Funktionen, die auch in Stresssituationen zum Tragen kommen. So verhindert Callose u.a. die weitere Ausbreitung von Viren über die Plasmodesmata. Bei Attacken durch Pilzhyphen oder bei Verwundungen wird Callose äußerst schnell als erste physische Barriere (Papillen) bzw. zum Wundverschluss durch die Pflanze synthetisiert. Solche Papillen, die neben Callose auch aus Lignin, weiteren Polysacchariden, phenolischen Komponenten, RO-Intermediaten und Proteinen bestehen, werden von resistenten Pflanzen schon unmittelbar nach Erkennen des Pathogens hergestellt und an den Penetrationsstellen abgelagert. Sie sollen so den drohenden Pathogeneintritt verhindern oder zumindest für eine Verlangsamung bzw. Demobilisierung des Pathogens sorgen. Der attackierten Zelle wird auf diese Weise in jedem Fall zu einem Aufschub verholfen, der ihr Zeit zur Einleitung weiterer Abwehrmaßnahmen wie dem oxidativen Burst oder für den Einsatz von antipathogenen Substanzen wie PR-Proteine (engl. pathogen-related protein, Pathogenzellwand-abbauende Enzyme) oder Phytoalexinen bietet. Kommt es doch zum Pathogeneintritt bzw. zur Infektion der Zelle, schottet sich diese mit Hilfe weiterer Calloseablagerungen an den Plasmodesmata ab. Diese Abschottung führt zur Akkumulation von Kohlenhydraten und über das sogenannte sugar sensing zur Aktivierung des sink-Stoffwechsels (Scharte, Schön & Weis, 2005; Schön 2005). Der nun auf Abwehr eingestellte Zellstoffwechsel startet eine ganze Palette weiterer Maßnahmen, an deren Ende der programmierte Zelltod (PCD: engl. programmed cell death) stehen kann.
Regulation
Die bei der Regulation der CalS durchlaufenen Signalwege sind noch weitgehend unbekannt. Allerdings wird Ca2+-Ionen eine wichtige Rolle zugeschrieben. Z. B. kommt es bei resistenten Pflanzen unmittelbar nach Beginn einer Pathogenattacke in den Zellen zur Öffnung Elicitor-abhängiger Membrankanäle, wodurch ein Einstrom von Ca2+-Ionen und parallel dazu auch von H+-Ionen ausgelöst wird. Die Ca2+-Konzentrationserhöhung stimuliert vermutlich über das oben erwähnte Annexin-ähnliche Membranprotein die Callosesynthese.
Eine Ca2+-unabhängige Isoform der Callosesynthase scheint während der Ausbildung der Zellplatte beteiligt zu sein. Die Aktivierung verläuft hier voraussichtlich über eine Phragmoplastin-Rop-Interaktion (Rop-Protein an der hydrophilen Schleife).
Daneben werden z. B. auch G-Proteine, Glykolysierung (Glykolysierungsstelle), Phosphorylierung (Phosphorylierungsstellen), der Einfluss von Phytohormonen (z. B. Abscisinsäure) und das Erreichen eines pH-Optimums für die CalS als Regulationsmechanismen diskutiert.
Siehe auch
Literatur
- McCurdy, DW. et al. (2008): Wall ingrowth formation in transfer cells: novel examples of localized wall deposition in plant cells. In: Curr Opin Plant Biol 11(6); 653–661; PMID 18849189; doi:10.1016/j.pbi.2008.08.005
- Will, T. und van Bel, AJ. (2006): Physical and chemical interactions between aphids and plants. In: J Exp Bot 57(4); 729–737; PMID 16473888; PDF (freier Volltextzugriff, engl.)
- Maor, R. und Shirasu, K. (2005): The arms race continues: battle strategies between plants and fungal pathogens. In: Curr Opin Microbiol 8(4); 399–404; PMID 15996507; doi:10.1016/j.mib.2005.06.008
- Verma, DP. und Hong, Z. (2001): Plant callose synthase complexes. In: Plant Molecular Biology 47(6): 693–701; PMID 11785931; doi:10.1023/A:1013679111111
- Hans W. Heldt und Birgit Piechulla: Pflanzenbiochemie. Spektrum Akademischer Verlag; 4. Auflage 2008; ISBN 978-3-8274-1961-3; S. 258f.
- Sitte P. et al. (2002): Strasburger – Lehrbuch der Botanik. Spektrum Akademischer Verlag. Heidelberg. Berlin. Oxford. 35. Auflage.
Weblinks


Wikimedia Foundation.