- Proteinkinase
-
Proteinkinase —
Enzymklassifikation EC, Kategorie 2.7.-.- Kinase Reaktionsart Phosphorylierung Substrat Proteine Produkte Phospho-Proteine Proteinkinasen sind Enzyme, die den Transfer einer Phosphatgruppe von einem Donor (meist ATP) auf die Seitenketten-Hydroxyl-(OH-)Gruppe einer Aminosäure katalysieren. Kinasen sind daher Phosphoryltransferasen.
Inhaltsverzeichnis
Evolution der Kinasen
Laut Manning codieren rund 2% aller Gene des eukaryotischen Genoms für Kinasen. Die rund 518 Kinasen können 30% aller Proteine einer Zelle phosphorylieren.[1] Die Summe aller Kinasen einer Zelle werden auch Kinom genannt (in Anlehnung an -omik). Eine Klassifizierung, basierend auf Sequenzähnlichkeiten der Kinasedomänen, bekannter biologischer Funktionen, zeigt 10 Gruppen, 143 Familien und 212 Subfamilien in den Kinomen von Menschen, Fliegen, Würmern und der Hefe.
Funktionen der Kinasen
Die Proteinphosphorylierung ist ein sehr wichtiger post-translationaler Kontrollmechanismus in der Signaltransduktion der Zelle. Hierzu gehört zum Beispiel die Regulation der Aktivität von Enzymen oder Transkriptionsfaktoren. Die Phosphorylierung kann durch Ladungs- oder Konformationsänderungen das Zielprotein aktivieren/inhibieren oder seine subzelluläre Lokalisierung bestimmen. Die physiologischen Auswirkungen dieser Proteinphosphorylierungen hängen vom jeweiligen Substrat der Kinase ab.
Dysfunktionen von Proteinkinasen sind die Ursache von zahlreichen Erkrankungen. Damit sind Proteinkinasen attraktive molekulare Ziele medikamentöser Intervention und werden von der pharmazeutischen Industrie intensiv erforscht. "Spezifische" Inhibitoren von Proteinkinasen werden erfolgreich bei der Krebstherapie eingesetzt (siehe z. B. Imatinib in der Behandlung der Chronisch myeloischen Leukämie).
Regulation der Kinaseaktivität
Auf Grund der weitreichenden Einflüsse der Proteinkinasen müssen diese selbst streng reguliert werden. Regulierende Faktoren sind unter anderem:
- Kofaktoren/Sekundäre Botenstoffe (Ca2+, IP3, cAMP et cetera)
- Aktivator- und Inhibitorproteine
- Pseudosubstrate
- Autoinhibition (ein Teil der Peptidkette der Proteinkinase agiert als Pseudosubstrat)
- Ligandenbindung an regulatorische Untereinheiten
- Phosphorylierung im aktiven Zentrum durch
- andere Proteinkinasen (Trans-phosphorylierung)
- sich selbst (Cis-phosphorylierung/Auto-phosphorylierung)
- Subzellulärer Lokalisierung innerhalb der Zelle
Typen von Proteinkinasen
Die meisten Proteinkinasen phosphorylieren entweder die Aminosäuren Serin/Threonin oder aromatische Tyrosine. Es gibt jedoch auch bispezifische Proteinkinasen (EC 2.7.12; z.B. MEK oder MAP2K), die sowohl Serin/Threonin als auch Tyrosin-Reste phosphorylieren können.
Serin/Threonin-Kinasen
Diese Proteinkinasen (EC 2.7.11) phosphorylieren die Hydroxylgruppen (OH-Gruppen) der Aminosäuren Serin und Threonin. Diese Kinasen werden reguliert durch:
- cAMP oder cGMP
- 1,2-Diacylglycerine (DAG)
- Ca2+ bzw. Calmodulin
- PIP3 und anderen Phospholipid-Derivaten
Dass Serin/Threonin-Kinasen nicht alle Serine und Threonine anderer Proteine phosphorylieren, wird durch die Interaktion mit der umgebenden Peptidsequenz bestimmt. Diese Sequenzen werden Konsensussequenzen genannt. Durch die geringe Spezifität werden durch diese Kinasen keine einzelnen Proteine phosphoryliert, sondern ganze Proteinfamilien. Inhibiert werden diese Enzyme, indem ein Pseudosubstrat an das aktive Zentrum bindet, indem es die Zielsequenz der entsprechenden Kinase nachahmt, jedoch über kein Serin oder Threonin verfügt.
Muskel Phosphorylase Kinase A
Dieses Enzym (EC 2.7.11.19) ist die erste Ser/Thr-Kinase, die im Jahr 1956 entdeckt wurde[2]. Es handelt sich dabei um ein Schlüsselenzym des Glykogenstoffwechsels.
Proteinkinase A
Dieses Enzym besitzt einige Funktionen in der Zelle. Hierzu zählen die Regulation des Glycogen-, Zucker- und Lipidmetabolismus. Die Protein Kinase A (EC 2.7.11.11) besteht aus zwei Domänen. Die kleinere besteht aus einer großen Zahl β-Faltblättern, wohingegen die große Untereinheit eine große Zahl von α-Helices besitzt. Das katalytische Zentrum liegt zwischen den beiden Untereinheiten. Bindet ATP an ein Substrat, verdrehen sich die Untereinheiten gegeneinander, sodass die γ-Phosphatgruppe des ATP in die Nähe der zu phosphorylierenden Aminosäure kommt und die Transferreaktion stattfinden kann.
Sie selbst werden durch cAMP reguliert. Durch Bindung von cAMP an ein inaktives Tetramer aus zwei regulatorischen und zwei katalytischen Untereinheiten (R2C2) werden die regulatorischen Untereinheiten von den katalytischen abgetrennt, wodurch die Phosphorylierung anderer Proteine ermöglicht wird. Die Proteinkinase A wird wiederum durch Phosphorylierung reguliert.
Außerdem wird durch die Aktivierung einer Phosphodiesterase die verfügbare Menge an cAMP durch Umwandlung in AMP gesenkt. Dies führt dazu, dass die Proteinkinase A ihre eigene Hemmung verursacht und es dadurch zu keiner "Daueraktivierung" der Kinase kommen kann.
Proteinkinase B
Die in der vor allem englischsprachigen Literatur oftmals als Akt bezeichnete Kinase ist ein Enzym des PI3K/Akt-Signalweges, der zahlreiche Auswirkungen auf die Homöostase der Zelle hat, und das Überleben, die Apoptose, die Proliferation und den Stoffwechsel reguliert. Akt kommt in drei eng miteinander verwandten Isoformen vor (Akt1, Akt2, Akt3) deren Sequenzen auf Chromosom 14q32,19q13 respektive 1q43 kodiert werden. Keine Substratspezifitätsunterschiede konnten zwischen den einzelnen Isoformen festgestellt werden.
Die Aktivierung erfolgt in mehreren Schritten. Zunächst werden Phosphatidylinositol-3-Kinasen (PI3K) durch aktivierte Rezeptortyrosinkinasen zur Membran rekrutiert und somit aktiviert. Aktivierte PI3K katalysieren die Phosphorylierung von Phosphatidylinositol zu Phosphatidylinositol-3-Phosphat bzw. von Phosphatidylinositol-4-Phosphat zu Phosphatidylinositol-3,4-Bisphosphat und von Phosphatidylinositol-4,5-Bisphosphat zu Phosphatidylinositol-3,4,5-trisphosphat. Die so entstandenen Substrate rekrutieren nun Proteine mit Pleckstrin Homology(PH)-Domänen zur Membran. Das so angedockte Akt kann nun über ein weiteres PH-Domänen aufweisendes Signalmolekül, das PDK1 (phosphoinositide-dependent kinase 1) phosphoryliert, aktiviert werden.
Wie oben erwähnt reguliert die Proteinkinase B unter anderem das Überleben, die Proliferation und den Sterbezyklus der betroffenen Zelle. Ein Beispiel ist das vermehrte Auftreten vom Glucose-Rezeptor Glut4 bei einem eingehenden Insulinsignal. Dabei wird die Transkription dieses Carriers kaskadenartig angefeuert und der Einbau desselben durch Vesikeltransport und -Abschnürung veranlasst. Selbst reguliert werden kann die Aktivität des Enzym beispielsweise durch das Tumor-Suppressor-Protein PTEN, welches für die Dephosphorylierung von bspw. Phosphatidylinositol-3,4,5-trisphosphat zu Phosphatidylinositol-4,5-bisphosphat (oder anderen 3'-phosphorylierten Phosphoinositiden) zuständig ist. Damit wird das Substrat der Proteinkinase untauglich, da es nun nicht mehr imstande ist, an die PH-Domäne anzudocken.
Die Funktion und Regulationsmechanismen sind mannigfaltig und werden heute noch eingehend untersucht, da angenommen wird, dass eine Mutation in bestimmten Bereichen des Signalweges (Bsp.: PTEN) eine Grundlage für die Ausbildung von Tumoren ist.
Proteinkinase C
Die Bezeichnung Proteinkinase C (EC 2.7.11.13) trifft auf eine Familie von Proteinen mit in Säugetieren 12 Mitgliedern zu, die Ca2+, Diacylglycerol, und ein Phospholipid wie z. B. Phosphatidylcholin zur Aktivierung benötigen. In den meisten Fällen wird Proteinkinase Cα gemeint, wenn von diesen Enzymen die Rede ist.
Die Proteinkinasen sind hochkonservierte Proteine, die aus einer N-terminalen regulatorischen Domäne und einer C-terminalen katalytischen Domäne bestehen. Solange das Enzym nicht durch einen Tumorpromotor wie Tetradecanoyl-Phorbolacetat (TPA) oder einen der oben genannten Kofaktoren aktiviert wird, ist es inaktiv. Die allgemeine lineare Struktur:
N - Pseudosubstrat - TPA/DAG-Bindung - Ca2+-Bindung - ATP-Bindung - Substratbindung - C
Während der Aktivierung verlagert sich die Proteinkinase C zur Zellmembran mit Hilfe von Rezeptoren für aktivierte Proteinkinase C (RACK-Proteine). Nach Aktivierung bleiben diese Kinasen lange Zeit aktiv, obwohl die Ca2+-Konzentration wieder gesunken ist. Dies wird auf die Wirkung von Diacylglycerol zurückgeführt, das aus Phosphatidylinositol durch eine Phospholipase gebildet wird. Diese wird durch die gleichen Signale aktiviert, wie die Proteinkinase selbst.
Die Zielsequenz der Proteinkinase C ist der der Proteinkinase A ähnlich, da sie viele basische Aminosäurereste in der Nähe der phosphorylierten Ser/Thr-Reste enthält. Substrate der Proteinkinase C sind MARCKS Proteine, MAP Kinasen, Transkriptionsfaktor-Inhibitor IκB, Vitamin D3 Rezeptor, Raf Kinase, Calpain und der EGF-Rezeptor.
Tyrosin-Kinasen
Unter den Tyrosinkinasen (EC 2.7.10) gibt es so genannte Rezeptor-Tyrosin-Kinasen (RTKs) sowie Tyrosin-Kinasen ohne Rezeptorfunktion. Es werden an die 50 verschiedenen RTKs in 18 Rezeptor-Familien zusammengefasst.[3] Nach den Liganden werden diese RTKs in folgende Familien gruppiert:
Referenzen
- ↑ Manning G. Genomic overview of protein kinases. WormBook. 2005 Dec 13;:1-19. Review. PMID 18050405
- ↑ Krebs EG, Fischer EH. The phosphorylase b to a converting enzyme of rabbit skeletal muscle. Biochim Biophys Acta. 1956 Apr;20(1):150-7. PMID 13315361
- ↑ Aaronson SA. Growth factors and cancer. Science. 1991 Nov 22;254(5035):1146-53. Review. PMID 1659742
Weblinks
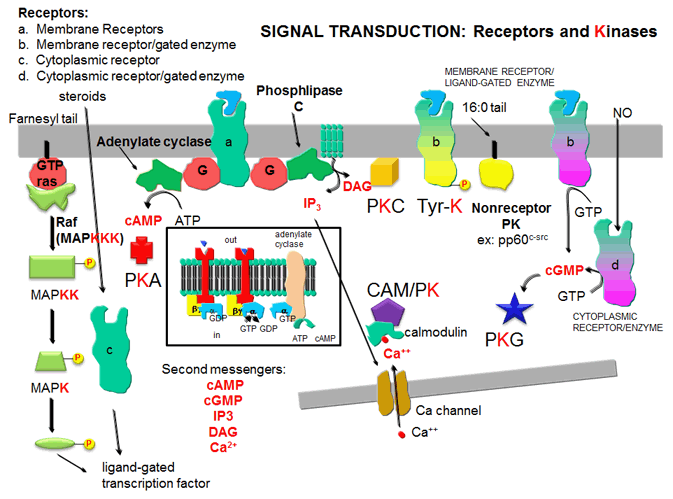
- Receptors and Kinases: five major protein kinases - Darstellung der Rolle wichtiger Proteinkinasen bei der Signaltransduktion (engl.)
- The Protein Kinase Resource - Umfassende Darstellung des Kinoms mit interaktiven Vergleichsmöglichkeiten einzelner Kinasen, bzgl. Funktion und Struktur (engl.)
Kategorien:- Proteinkinase
- Proteingruppe
{kind=link}
Wikimedia Foundation.