- Proteinkinase C
-
Proteinkinase C —
Kofaktor DAG (konv./nov.); Ca2+ (konv.) Isoformen α/βI/βII/γ (konventionell); δ/ε/η/θ (novel); Mζ/ι/λ (atypisch) Enzymklassifikation EC, Kategorie 2.7.11.13 Kinase Reaktionsart Phosphorylierung Substrat ATP + Protein Produkte ADP + Phosphoprotein Vorkommen Übergeordnetes Taxon Chordatiere Die Proteinkinase C (kurz PKC) ist ein Enzym der Familie der Proteinkinasen. Durch eine Übertragung von Phosphat auf Serin- oder Threoningruppen steuert sie die Aktivität nachgeordneter Enzyme oder Faktoren. Auf Grund dieser regulatorischen Funktion besitzt die Proteinkinase C eine zentrale Rolle bei der zellulären Signalweiterleitung (Signaltransduktion). Calciumionen (Ca2+), Phospholipide und Diacylglycerin sind für die Aktivität der PKC nötig. Sphingosin hemmt dagegen die PKC.
Entdeckt wurde die PKC durch die Arbeiten von Yasutomi Nishizuka und Mitarbeitern als Phospholipid-abhängige Kinase, deren Aktivität zusätzlich durch Diacylglycerin gesteigert werden konnte.[1]
Wichtige weitere Mitglieder der Enzymklasse der Proteinkinasen sind Proteinkinase A, Proteinkinase B und Proteinkinase G.
Inhaltsverzeichnis
Struktur
Die Proteinkinase C besteht aus einer regulatorischen N-terminalen (R) und einer katalytischen C-terminalen Domäne (C). Wie auch für die Proteinkinase A beschrieben, weist die regulatorische Domäne eine Pseudosubstratsequenz auf, die das aktive Zentrum der katalytischen Domäne im Ruhezustand blockiert: Anstelle eines phosphorylierbaren Serinrestes ist hier ein Alaninrest vorhanden.
Derzeit sind zehn Isoenzyme der Proteinkinase C bekannt. Sie wurde inzwischen aus Drosophila und zahlreichen Säugern kloniert. Man unterscheidet drei Gruppen der PKC-Isoenzyme: die klassische PKC (cPKC), die neue PKC (nPKC) und atypische PKC (aPKC). Ihre Molekülmassen betragen 61 bis 154 kDa. Zu den cPKC-Isoformen zählen α, β1, β2 und γ, zu den nPKC-Isoformen zählen ε, δ, η und θ und die aPKC-Isoformen bestehen aus ζ und λ/τ. In der Literatur ist auch eine PKCμ bekannt, sie wird jedoch als Proteinkinase D1 bezeichnet.[2]
Die cPKC-Isoformen werden durch die Sekundärsignale Ca2+ und Diacylglycerin (DAG) aktiviert, die nPKC-Isoformen werden nur durch DAG aktiviert, und aPKC sind dagegen Ca2+- und DAG-unabhängig. Zusätzlich existieren weitere, zell- und isoformabhängige Aktivierungs- und Inaktivierungswege
Bei PKCλ und PKCτ handelt es sich um die sogenannten orthotopen Enzyme, in den Mäusen wird PKCλ exprimiert, während beim Menschen die gleichen Funktionen von PKCτ wahrgenommen werden.
Funktion und Regulation
Signaltransduktion
 Schematische Übersicht über die Aktivierung der Proteinkinase C. Für Einzelheiten bitte Text beachten.
Schematische Übersicht über die Aktivierung der Proteinkinase C. Für Einzelheiten bitte Text beachten.
Die Proteinkinase C besitzt eine zentrale Bedeutung bei der Signaltransduktion. Ihre Aktivität wird über Hormone und Neurotransmitter gesteuert, deren Signal über sekundäre Botenstoffe, sogenannte Sekundärsignal, weitergeleitet wird. Enzyme der PKC-Familie sind nicht von Beginn an aktiv. Stattdessen unterliegen sie – je nach Isoenzym – einer komplexen Aktivierungsfolge und werden zum gewünschten Wirkort gebracht, bevor sie vollständig katalytisch aktiv sind.
Bindung einer Reihe von Neurotransmittern, Wachstumsfaktoren und Hormonen an ihre G-Protein-gekoppelten Rezeptoren (GPCR) vermittelt über die Aktivierung der Phospholipase C (PLC) die Freisetzung der second messenger Inosit-1,4,5-trisphosphat (IP3) und Diacylglycerin (DAG) aus der Membrankomponente Phosphatidylinositol-4,5-bisphosphat (PIP2) (vgl. Abbildung). IP3 bindet an Rezeptoren in der Membran intrazellulärer Calciumspeicher (endoplasmatisches Retikulum und/oder Mitochondrien) und verursacht die Freisetzung von Ca2+. Dieser Ausstrom von Calciumionen ins Cytosol führt zu einer Aktivierung von calciumabhängigen Proteinkinasen.
Für die Funktion der Proteinkinase C wird Ca2+ benötigt. Darüber hinaus ist die Bindung an Phosphatidylserin, eine Lipidkomponente der inneren Seite der Zellmembran, erforderlich. An das bei der Hydrolyse von PIP2 entstandene Diacylglycerin, ein Sekundärsignal, bindet die peripher-membrangebundenen Form von PKC, die mit der cytosolischen Form in Gleichgewicht steht.[3] Dadurch wird PKC aktiviert und katalysiert die Phosphorylierung vieler Zielproteine.
Zellproliferation
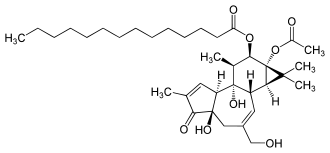 Struktur von TPA, ein karzinogener Aktivator der PKC.
Struktur von TPA, ein karzinogener Aktivator der PKC.PKC hat Bedeutung bei der Regulierung des zellulären Wachstums. Eine Fehlsteuerung kann an der Auslösung von Krebs und an der Entstehung diabetischer Spätkomplikationen[4] beteiligt sein.
Die Bedeutung der PKC für Zellteilung und Proliferation wurde offensichtlich, als man die Wirkungsweise der Phorbolester erkannte. Phorbolester, polyzyklische Alkoholderivate wie beispielsweise 12-O-Tetradecanoylphorbol-13-acetat (TPA), sind wichtige Karzinogene, die zwar nicht selbst eine Tumorbildung initiieren, aber dafür die von kanzerogenen Stoffe fördern.[5] Phorbolester aktivieren das Enzym aufgrund ihrer Ähnlichkeit zum natürlichen Aktivator DAG. Die dadurch vermittelte Aktivität hat lange Bestand, da Phorbolester im Gegensatz zu DAG nur langsam abgebaut werden.
Bei Patienten mit Diabetes mellitus führen erhöhte Blutzuckerspiegel zu einem Anstieg der Diacylglycerol-Konzentration in der Zelle und so zu einer Aktivierung der Proteinkinase C. Dies fördert die Produktion von extrazellulärer Matrix und Zytokinen, erhöht die Kontraktilität und Permeabilität (Durchlässigkeit) von Blutgefäßen, steigert des Zellwachstum in Blutgefäßen, aktiviert die Phospholipase A2 und hemmt die Na+/K+-ATPase. Die Folge sind Gefäßschäden der Netzhaut des Auges, der Niere und des Herzens. Ruboxistaurin, ein Hemmstoff der Proteinkinase C, ist möglicherweise in der Lage, bei Patienten mit Diabetes Schäden an den kleinen Blutgefäßen (Mikroangiopathie) günstig zu beeinflussen.[6]
Signalprozesse in Immunzellen
Ob PKC-Isoenzyme eine Funktion bei Signalprozessen in Immunzellen haben, ist Gegenstand der Forschung.[7][2] Möglicherweise spielen im Signalweg von B- oder T-Zellen die Isoenzyme α, β und θ eine wichtige Rolle.[8] So soll nPKC-θ für die Aktivierung von T-Zellen wichtig sein. In Mäusen wurde gezeigt, dass ihre T-Zellen schwere Störungen bei der T-Zellen vermittelten Aktivierung aufweisen, wenn man bei ihnen keine nPKC-θ gebildet wird. Dagegen ist ihre B-Zell-Funktion normal.[2]
Neben dem θ-Isoenzym könnte auch das α-Isoenzym für T-Zellen bedeutsam sein. Bei einer Aktivierung der T-Zellen wird die Ausschüttung von PKC-α kurzfristig erhöht. PKC-β ist für die Aktivierung von B-Zellen wichtig.[9] Wenn man in Mäusen die Gene für die β1- und β2-Isoform entfernt, bildet sich eine Immundefizienz aus. Auch die aPKC-ζ ist für Funktion der B-Zellen wichtig.[2]
Einzelnachweise
- ↑ Nakamura, S. und Yamamura, H. (2010): Yasutomi Nishizuka: father of protein kinase C. In: J Biochem. 148(2); 125–130; PMID 20668066; PDF (freier Volltextzugriff, engl.)
- ↑ a b c d Daisuke Kitamura: How the Immune System Recognizes Self and Nonself: Immunoreceptors and Their Signaling. Springer-Verlag GmbH 2007; ISBN 978-4431738831; S. 107f.
- ↑ H. Robert Horton, Laurence A. Moran, K. Gray Scrimgeour, Marc D. Perry, J. David Rawn und Carsten Biele (Übersetzer): Biochemie. Pearson Studium; 4. aktualisierte Auflage 2008; ISBN 978-3-8273-7312-0; S. 386ff.
- ↑ D. Koya, G. L. King: Protein kinase C activation and the development of diabetic complications. In: Diabetes 47: S. 859–866.
- ↑ Gerhard Krauss: Biochemistry of Signal Transduction and Regulation. Wiley-VCH; 2., veränd. Auflage 2001; ISBN 978-3527303779; S. 259
- ↑ V. J. Scott u.a.: Ruboxistaurin, a Protein Kinase C ß Inhibitor, as an Emerging Treatment for Diabetes Microvascular Complications In: Ann Pharmacother. 39: S. 1693–1699.
- ↑ Baier, G. (2003): The PKC gene module: molecular biosystematics to resolve its T cell functions. In: Immunol Rev. 192; 64–79; PMID 12670396; doi:10.1034/j.1600-065X.2003.00018.x
- ↑ Baier, G. und Wagner, J. (2009): PKC inhibitors: potential in T cell-dependent immune diseases. In: Curr Opin Cell Biol. 21(2); 262–267; PMID 19195860; doi:10.1016/j.ceb.2008.12.008
- ↑ Guo, B., Su, TT. und Rawlings, DJ. (2004): Protein kinase C family functions in B-cell activation. In: Curr Opin Immunol. 16(3); 367–373; PMID 15134787; doi:10.1016/j.coi.2004.03.012
Weblinks
Kategorien:- Proteinkinase
- Proteingruppe


Wikimedia Foundation.