- Embden-Meyerhof-Weg
-
Glykolyse (aus dem Griechischen glykys = süß und lysis = auflösen) ist der erste Teil des Glukose-Abbaus (Katabolismus) in den Zellen der meisten Lebewesen. In diesem biochemischen Abbauweg wird ein Molekül Glukose in zehn enzymatisch katalysierten Reaktionen in zwei Pyruvat-Moleküle umgewandelt.
Die sehr gut bekannte und untersuchte Form der Glykolyse ist der Embden-Meyerhof-Weg (nach Gustav Embden und Otto Meyerhof). Ein alternativer Weg ist der Entner-Doudoroff-Weg. Die Bezeichnung Glykolyse kann verwendet werden, um alle alternativen Abbauwege mit einzuschließen. Glykolyse wird jedoch hier als Synonym für den Embden-Meyerhof-Weg verwendet.
Inhaltsverzeichnis
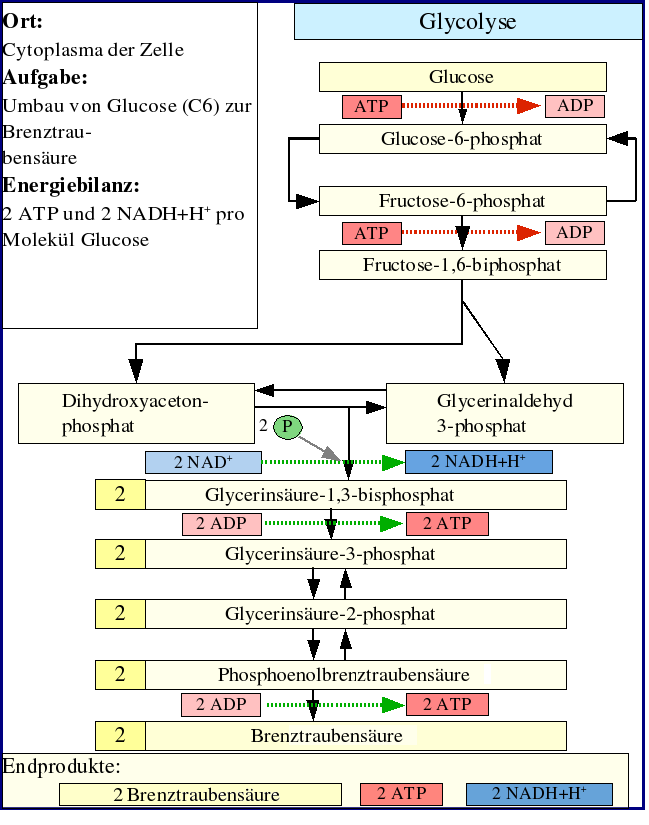
Ort der Glykolyse
Die Glykolyse ist der wichtigste Abbauweg der Kohlenhydrate im Stoffwechsel und findet im Cytoplasma jeder Zelle statt. Verstand man früher darunter lediglich den Abbau von Kohlenhydraten unter Sauerstoffmangelbedingungen über Pyruvat zu Lactat, so weiß man heute, dass der Abbau von Glukose bis zu Pyruvat auch bei ausreichendem Sauerstoffangebot gleichartig abläuft: sie findet in beiden Fällen ohne "Verbrauch" von Sauerstoff statt.
In Prokaryoten und in überwiegend anaerob arbeitenden Zellen oder Geweben (Skelettmuskel) von Eukaryoten wird Pyruvat anaerob zu Milchsäure oder wie bei vielen Hefen zu Ethanol und Kohlenstoffdioxid (CO2) verstoffwechselt.
Aerob arbeitende Gewebe (Prototyp: Herzmuskel) bauen den „C3-Körper“ Pyruvat zu Acetyl-CoA und Kohlenstoffdioxid ab und das Acetyl-CoA weiter im Citratzyklus zu CO2 und Wasserstoff. Der Wasserstoff wird dabei an die Wasserstoffüberträger Nicotinamidadenindinukleotid (NAD) und Flavinadenindinukleotid (FAD) gebunden (NADH bzw. FADH2). Die Energie aus der Oxidation des Wasserstoffs in der Atmungskette der Mitochondrien wird zur Synthese von ATP genutzt (oxidative Phosphorylierung).
Die Glykolyse ist der einzige metabolische Weg, den praktisch alle modernen Organismen gemeinsam haben, was auf eine sehr frühe Entstehung hinweist; die Glykolyse entstand möglicherweise in den ersten Prokaryoten vor rund 3,5 Milliarden Jahren.
Die Reaktionsschritte der Glykolyse
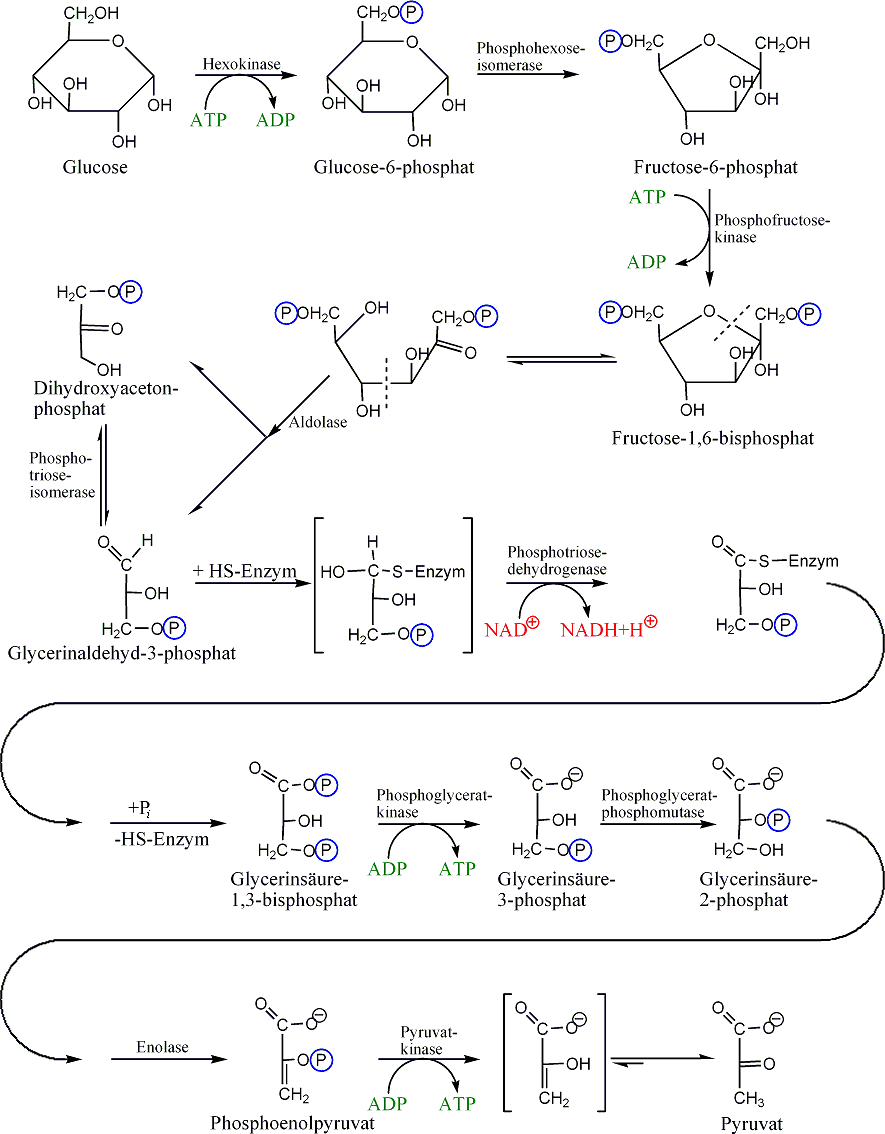
Der erste Schritt der Glykolyse ist die Phosphorylierung von Glukose zu Glucose-6-phosphat (G6P). In Abhängigkeit vom Zelltyp wird diese Reaktion durch das Enzym Hexokinase (in allen Körperzellen, wie zum Beispiel im Gehirn) oder Glucokinase (nur in Leber und Pankreas) katalysiert. Bei der Phosphorylierung wird 1 ATP verbraucht, was allerdings eine gute Investition darstellt. Die Zellmembran ist nämlich durchlässig für Glukose, aber nicht für das durch die Phosphorylierung entstehende Glukose-6-phosphat, welches sich dadurch in der Zelle anreichert und durch Verschiebung des Gleichgewichts an der Membran die Aufnahme von Glukose begünstigt. Glucose-6-phosphat wird dann von der Glucose-6-phosphat-Isomerase in Fructose-6-phosphat (F6P) umgebaut. (An diesem Punkt kann auch Fruktose durch Phosphorylierung in den glykolytischen Weg eintreten.)
Danach wird Fruktose-6-phosphat unter Einwirkung des Schlüsselenzyms der Glykolyse Phosphofructokinase 1 mit einem Molekül ATP zu Fructose-1,6-bisphosphat (1,6-FBP) phosphoryliert, wobei aus ATP ADP gebildet wird. Die damit verbundene Übertragung von Energie ist auf zweierlei Weise gerechtfertigt: Zum einen macht auch dieser Schritt – neben der Glukokinase sowie der Pyruvatkinase – die Glykolyse irreversibel, zum anderen erlaubt die zweite Phosphatgruppe die Spaltung des Fruktoserings durch Aldolase in Dihydroxyacetonphosphat (DHAP) (phosphorylierte Keto-Triose) und Glycerinaldehyd-3-phosphat (3-GAP) (phosphorylierte Aldo-Triose). Dihydroxyacetonphosphat wird von der Triosephosphatisomerase (TIM) in Glycerinaldehyd-3-phosphat umgewandelt. Jedes der beiden resultierenden Glycerinaldehyd-3-phosphat-Moleküle wird dann durch NAD+, Glycerinaldehyd-3-phosphat-Dehydrogenase (GAPDH) und der Übertragung einer Phosphatgruppe zu 1,3-Bisphosphoglycerat (1,3-BPG) oxidiert. Dabei entsteht in einer Zwischenstufe ein Thioester. In chemischem Sinne findet mit dieser Reaktion die Oxidation der Carbonylgruppe zur Carboxyl-Gruppe, d.h. der Übergang vom Zucker(-Phosphat) zum Carbonsäure(-Phosphat) statt.
Im nächsten Schritt erzeugt die Phosphoglyceratkinase je ein Molekül ATP bei der Umwandlung von 1,3-Bisphosphoglycerat zu 3-Phosphoglycerat durch Übertragung eines Phosphatrests auf ADP. Damit ist die Energiebilanz der bisherigen Glykolyse ausgeglichen: zwei Moleküle ATP wurden verbraucht und zwei wiedergewonnen. Diese ATP-Synthese braucht ADP als Basis. Falls die Zelle bereits viel ATP (und damit wenig ADP) hat, hält die Reaktion an dieser Stelle an, bis wieder genügend ADP zur Verfügung steht. Diese Feedbackregulation ist wichtig, da ATP relativ schnell zerfällt, wenn es nicht genutzt wird. Überproduktion von ATP wird somit verhindert. Phosphoglyceratmutase katalysiert dann die Umwandlung von 3-Phosphoglycerat zu 2-Phosphoglycerat, woraus schließlich mit Hilfe der Enolase Phosphoenolpyruvat wird. Dieses wird schließlich in der Pyruvatkinasereaktion unter Erzeugung eines weiteren ATP zu Pyruvat (= Anion der Brenztraubensäure) umgesetzt. Auch dieser Schritt ist ADP-reguliert.
Einen in Erythrozyten vorhandenen alternativen Nebenweg vom 1,3-Bisphosphoglycerat zum 3-Phosphoglycerat stellt der über das Intermediat 2,3-Bisphosphoglycerat verlaufende Rapoport-Luebering-Zyklus dar, dessen zentrales Enzym die Bisphosphoglyceratmutase ist.
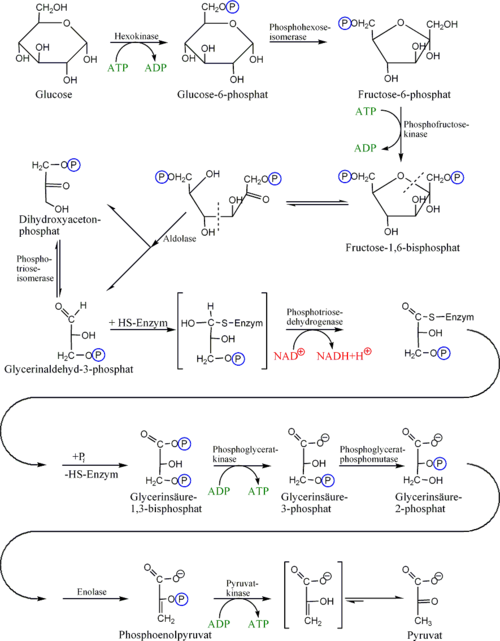
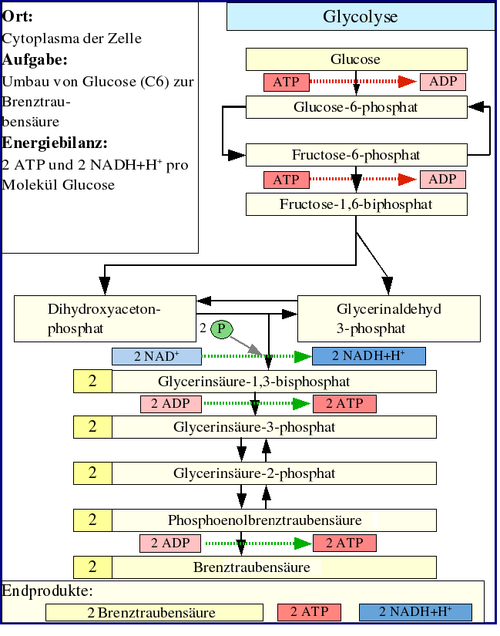
Reaktionsverlauf der Glykolyse Energetische Aspekte der Glykolyse
Gleichgewichtslage
Schritt Reaktion in der Glykolyse ΔG0’ [kJ/mol][1][2] ΔG [kJ/mol][2] 1 Glucose + ATP → Glucose-6-P + ADP −16,7 −34 2 Glucose-6-P ↔ Fructose-6-P +1,7 −2,9 3 Fructose-6-P + ATP → Fructose-1,6-P + ADP −14,2 −19 4 Fructose-1,6-P ↔ DHP + G-3-P +23,8 −0,2 5 DHP ↔ G-3-P +7,5 +2,4 6 G-3-P + Pi + NAD+ ↔ 1,3-Bis-P-glycerat + NADH + H+ +6,3 −1,3 7 1,3-Bis-P-glycerat + ADP ↔ 3-P-glycerat + ATP −18,8 +0,1 8 3-P-glycerat ↔ 2-P-glycerat +4,4 +0,8 9 2-P-glycerat ↔ PEP + H2O +7,5[1] bzw. +1,8[2] +1,1 10 PEP + ADP → Pyruvat + ATP −31,4 −23,0 Bemerkenswert ist, dass die meisten Reaktionen unter Standardbedingungen (ΔG0’) energetisch ungünstig sind. Sie würden kaum ablaufen, wenn sie nicht durch die energetisch günstigen Kinasereaktionen (Phosphofructokinase 1 (PFK 1), Phosphoglyzeratkinase (PGK), Pyruvatkinase (PK)) „hindurchgezogen“ werden würden. Damit liegen unter physiologischen Bedingungen (ΔG) die Konzentrationsverhältnisse der Produkte so vor, dass bis auf drei Reaktionen die gesamte Glykolyse reversibel ablaufen kann.
Die hier zugrundeliegende Strategie ist erwähnenswert, begünstigt diese Gleichgewichtslage doch die Gluconeogenese, das ist die Synthese von Glukose aus Pyruvat bei günstigem Energiestatus. Dies erfordert alle Enzyme, bis auf zwei der erwähnten „Zugpferde“, die allein der Glykolyse zugeordnet und hier mit −19 und –23 kJ/mol stark exergonisch sind.
Zudem ist erwähnenswert, dass die Phosphofructokinase das Schrittmacher- und wesentliche Schlüsselenzym der Glykolyse ist. Verschiedene Substanzen aktivieren bzw. hemmen, vermittelt über die PFK, den weiteren Ablauf der Glykolyse. So wird das allosterische Enzym Phosphofructokinase durch ADP und AMP aktiviert, während ATP, Citrat und eine erhöhte Protonenkonzentration (bei Übersäuerung durch Lactat) als Hemmung wirken.
Energieausbeute unter anaeroben und aeroben Bedingungen
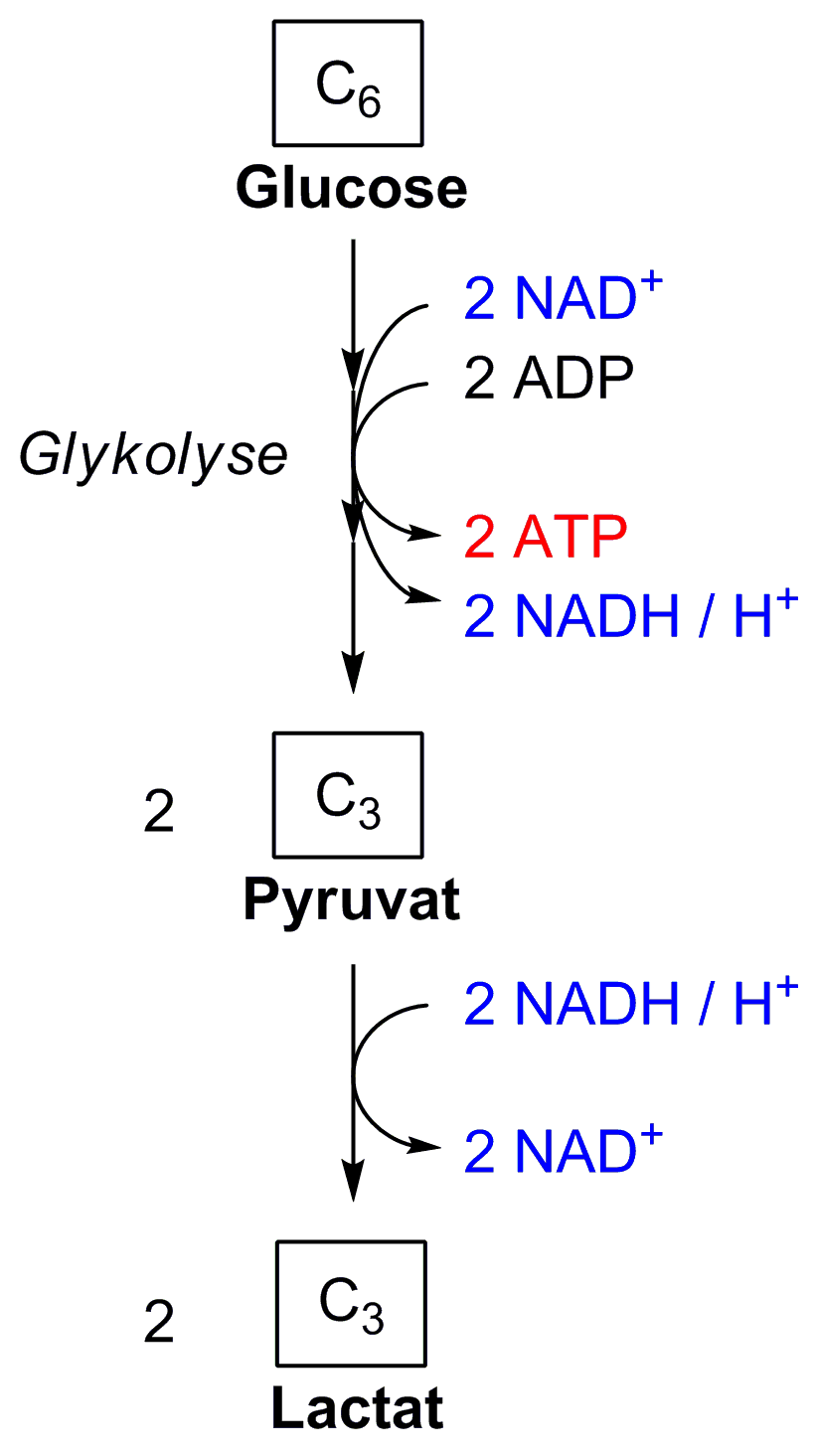
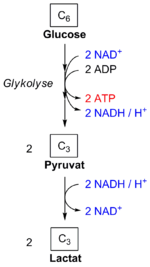 Bei der homofermentativen Milchsäuregärung wird das in der Glykolyse verbrauchte NAD+ in einer Folgereaktion wieder regeneriert.
Bei der homofermentativen Milchsäuregärung wird das in der Glykolyse verbrauchte NAD+ in einer Folgereaktion wieder regeneriert.Die Glykolyse liefert bei vollständigem Ablauf 2 Moleküle ATP pro Molekül Glukose. Aerobe Prokaryoten und die Mitochondrien von Eukaryonten können aus den beiden gleichzeitig entstehenden Molekülen Pyruvat aerob maximal weitere 34 Moleküle ATP gewinnen (also insgesamt 36 Moleküle ATP gewinnen); diese Bilanz hängt bei Eukaryoten vom Weg ab, auf dem im Cytosol gebildetes NADH + H+ die Mitochondrienmembran passiert (Malat-Aspartat-Shuttle bzw. Glycerin-3-Phosphat-Shuttle). Es kann allerdings auch in Abwesenheit von Sauerstoff die komplette Glykolyse ablaufen. Dies ist z.B. bei einigen Tumorzellen der Fall, aber auch bei Erythrozyten, denen die Mitochondrien für die Atmungskette fehlen.
Die Glykolysereaktionen bis zum Pyruvat werden sowohl unter aeroben als auch unter anaeroben Bedingungen durchlaufen. Die Regeneration des Coenzyms NAD+, das zur Oxidation des Glycerinaldehyd-3-phosphats durch die zugeordnete Dehydrogenase GAPDH eingesetzt und dabei zu NADH + H+ wird, erfolgt im ersten Fall in der Atmungskette. Im Falle starker Muskelbeanspruchung kann Sauerstoff nicht mehr schnell genug in die Zellen gelangen. Damit kann die Atmungskette nicht mehr ablaufen, und die Zelle muss ihre Energie ausschließlich aus der Glykolyse beziehen. Um NAD+ zu regenerieren, erfolgt hierbei die stark exergone (ΔG0´ = –25 kJ/mol bzw. ΔG = –14,8 kJ/mol)[2] Lactatdehydrogenase- (LDH-)Reaktion in der sog. Milchsäuregärung. Pyruvat wird mittels NADH + H+ zu Lactat und NAD+ reduziert, so dass die Glykolyse wieder ablaufen kann. Hefen, die Ethanol bilden, wird diese Funktion durch zwei Enzyme, Pyruvatdecarboxylase und Alkoholdehydrogenase übernommen.
Hemmstoffe
Iodacetat hemmt die Glycerinaldehyd-3-phosphat-Dehydrogenase, die Glycerinaldehyd-3-phosphat mit einem anorganischen Phosphat und unter Beteiligung von NAD+ zu 1,3-Bisphosphoglycerat oxidiert. Es modifiziert dabei eine SH-Gruppe des Enzyms, deshalb kann durch Zugabe von Mercaptanen diese Hemmung wieder aufgehoben werden.
Einzelnachweise
- ↑ a b David L. Nelson und Michael M. Cox: Lehninger Principles of Biochemistry. a. a. O. S. 553
- ↑ a b c d Garrett und Grisham: Biochemistry a. a. O. S. 582f.
Literatur
- Reginald Garrett und Charles M. Grisham: Biochemistry. (International Student Edition). Thomsom Learning Inc.; 3. Auflage 2005; ISBN 0-534-41020-0; S. 578–607
- David L. Nelson und Michael M. Cox: Lehninger Principles of Biochemistry. Palgrave Macmillan; 5. Auflage 2008; ISBN 978-0-7167-7108-1; S. 527–568
Weblinks
- Die Glycolyse auf der Homepage von Ulrich Helmich
- Audio-Beitrag zum Thema Glykolyse
- Jennifer McDowall/Interpro: Protein Of The Month: Enzymes of Glycolysis. (engl.)
Siehe auch
Wikimedia Foundation.