- Purkinjezelle
-
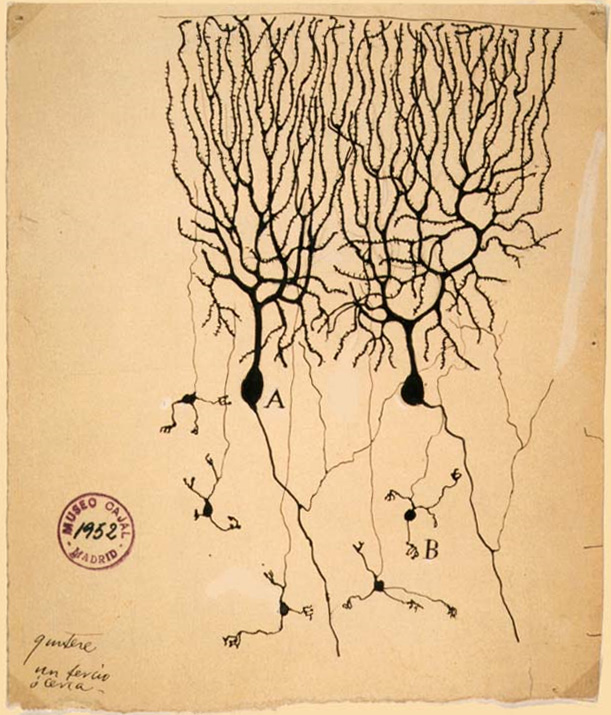
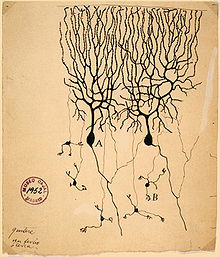 Historische Darstellung von zwei Purkinjezellen aus dem Kleinhirn der Taube
Historische Darstellung von zwei Purkinjezellen aus dem Kleinhirn der Taube
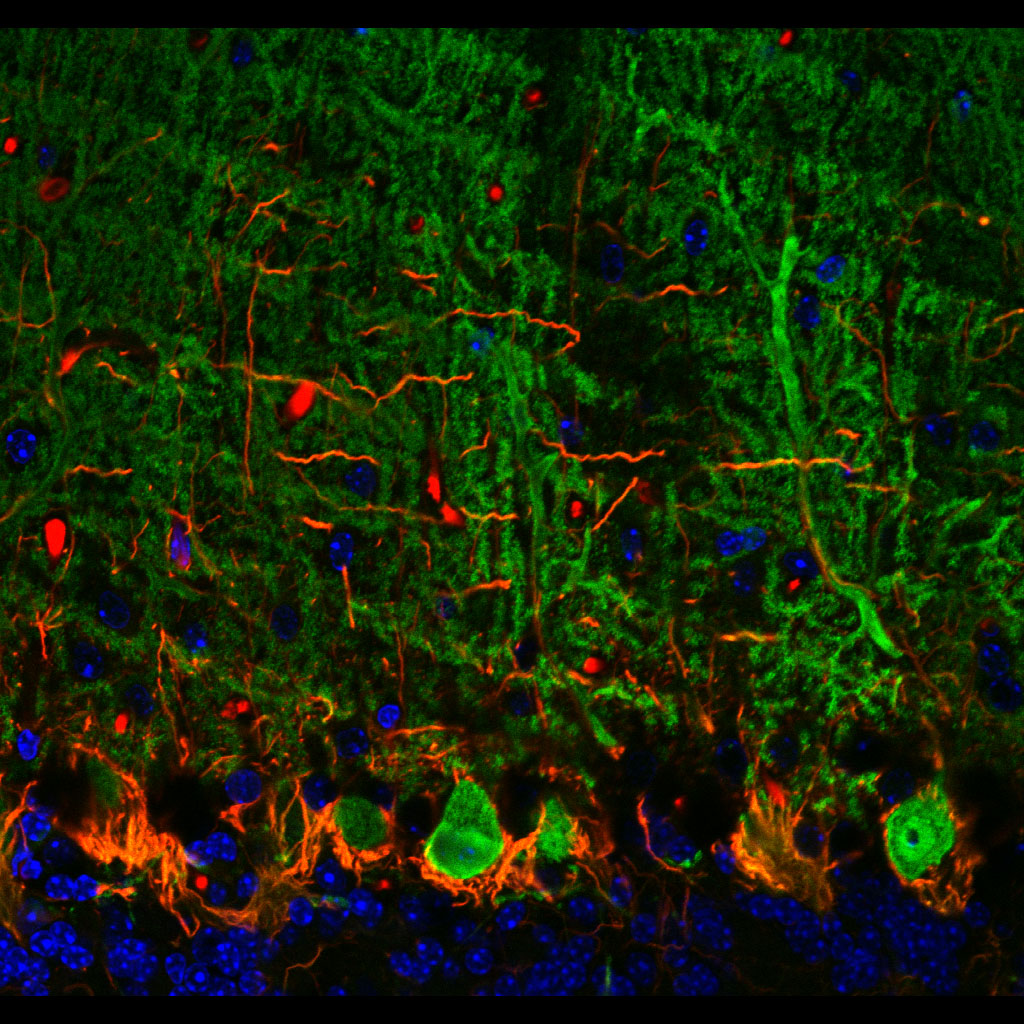
(Santiago Ramón y Cajal, 1899)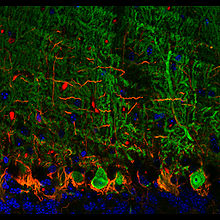 Purkinjezellen in grün (Calbindin) und Korbzellen in rot (Neurofilament) Kerne in blau (DAPI Kernfärbung). Kleinhirn der Maus, Vibratomschnitt, Fluoreszenzmikroskopie (Konfokales Laser Scanning Mikroskop Zeiss 510 META)
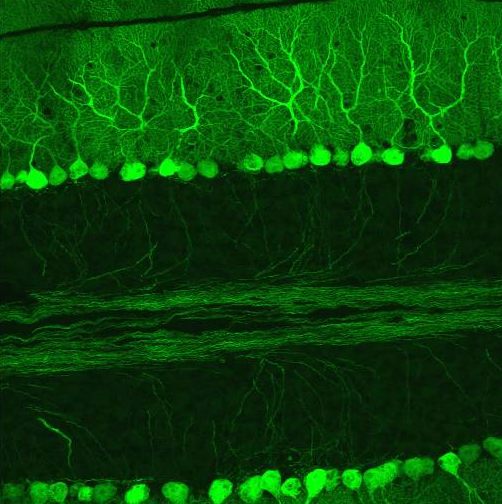
Purkinjezellen in grün (Calbindin) und Korbzellen in rot (Neurofilament) Kerne in blau (DAPI Kernfärbung). Kleinhirn der Maus, Vibratomschnitt, Fluoreszenzmikroskopie (Konfokales Laser Scanning Mikroskop Zeiss 510 META) Purkinjezellen in einem sagittalen Kleinhirnschnitt. Sie exprimieren den GFP-Abkömmling EGFP unter Kontrolle des Purkinjezell-spezifischen Promotors L7 und fluoreszieren deswegen bei Anregung mit blauem Licht.
Purkinjezellen in einem sagittalen Kleinhirnschnitt. Sie exprimieren den GFP-Abkömmling EGFP unter Kontrolle des Purkinjezell-spezifischen Promotors L7 und fluoreszieren deswegen bei Anregung mit blauem Licht.Die Purkinjezellen in der Kleinhirnrinde stellen die größten Neurone im Kleinhirn dar. Sie wurden nach ihrem Entdecker, dem tschechischen Physiologen Jan Evangelista Purkyně (1787-1869), der sie 1837 erstmals beschrieb, benannt. Ihre Axone stellen den einzigen Ausgang der Kleinhirnrinde dar. Purkinjezellen zeichnen sich durch einen verhältnismäßig großen Zellkörper und einen weit verzweigten Dendritenbaum aus. Dieser liegt praktisch vollständig in einer Ebene, und zwar in der sagittalen Schnittebene des Kleinhirns (s. Abb.). Der Dendritenbaum der Purkinjezellen liegt in der Molekularschicht und ihre Axone in der Körnerzellschicht der Kleinhirnrinde. Die Somata der Purkinjezellen bilden die Purkinjezellschicht.
Inhaltsverzeichnis
Synapsen der Purkinjezellen
Erregende Eingänge: Die Purkinjezelle erhält zwei erregende Eingänge, und zwar von der Kletterfaser und den Parallelfasern. Pukinjezellen sind glutamaterg, das heißt sie werden vom Transmitter Glutamat erregt. Die Kletterfaser entspringt der unteren Olive, einem Kerngebiet in der Medulla oblongata. Ihren Namen bezieht sie daher, dass sie den Dendritenbaum der Purkinjezelle aufsteigend umschlängelt: Besonders zu primären Dendriten im proximalen, also näher zum Soma gelegenen Teil, bildet sie besonders starke Synapsen mit der Purkinjezelle aus. An den präsynaptischen Endigungen ist die Wahrscheinlichkeit der Transmitterfreisetzung bei Eintreffen eines Aktionspotentials sehr hoch.
Kurz nach der Geburt sind die meisten Purkinjezellen zunächst von mehreren Kletterfasern innerviert. Durch Elimination überzähliger Synapsen bildet sich während der Entwicklung die typische Monoinnervation der Purkinjezellen durch jeweils eine Kletterfaser heraus.
Die Parallelfasern sind die Axone der Körnerzellen in der Körnerzellschicht. Diese steigen in die Molekularschicht auf und gabeln sich dort im rechten Winkel. Der Teil des Körnerzellaxons nach der Gabelung wird als Parallelfaser bezeichnet. Diese Fasern durchziehen in paralleler Anordnung (daher der Name) die Molekularschicht und treffen im rechten Winkel auf den Dendritenbaum der Purkinjezellen. Jede Parallelfaser innerviert viele Purkinjezellen, bildet dabei aber selten mehr als eine, nie mehr als zwei Synapsen. Das heißt, das Axon endet nicht am Ort der Synapse. Diese Art der Innervation wird auch als en-passant-Synapse bezeichnet. Jede Purkinjezelle besitzt ca. 150 000 - 200 000 Parallelfasersynapsen. Die einzelne Parallelfasersynapse ist "schwach" im Vergleich zur Kletterfasersynapse. Die Wahrscheinlichkeit für Transmitterfreisetzung im präsynaptischen Teil ist gering.- Hemmende Eingänge
Die Purkinjezelle wird in der Molekularschicht von zwei Interneurontypen hemmend innerviert. Im proximalen Teil des Dendritenbaums der Purkinjezellen sind das besonders die Korbzellen, mehr distal vor allem die Sternzellen. In der Körnerzellschicht finden sich die Golgizellen als weiterer Interneurontyp. Die Synapsen dieser drei Interneurone sind GABAerg, das heißt, sie benutzen GABA als Transmitter. Vor kurzem wurde entdeckt, dass entgegen bisherigen Annahmen auch die Lugarozellen in der Körnerzellschicht Synapsen mit der Purkinjezelle hat. Diese sind allerdings nur unter bestimmten Voraussetzungen aktiv. Zum Beispiel brauchen sie die Anwesenheit von Serotonin.
- Ausgang der Purkinjezelle
Die Axone der Purkinjezelle innervieren die Kleinhirnkerne. Sie stellen den einzigen Ausgang der Kleinhirnrinde dar. Da sie selbst GABA ausschütten, wirken sie hemmend. Im Axon der Purkinjezelle werden die Aktionspotentiale generiert, und zwar am ersten Ranvierschen Schnürring, ca. 75 µm vom Soma entfernt.
Spines
Der Dendritenbaum der Purkinjezellen ist, ähnlich wie der vieler zentraler Neurone, dicht mit Dornfortsätzen – den sogenannten spines – besetzt. Auf den spines der Purkinjezellen befinden sich die erregenden Synapsen. Die GABAergen hemmenden Synapsen besitzen keine spines.
Die spines der Purkinjezelle unterscheiden sich darin, ob sie von der Kletterfaser oder den Parallelfasern innerviert werden. Kletterfaser-spines befinden sich vor allem im proximalen (näher zum Soma gelegenen) Teil des Dendritenbaumes, während Parallelfaser-spines vor allem die dünnen distalen (weiter vom Soma entfernten) Bereiche des Dendritenbaumes besetzen.
Kletter- und Parallelfaser-spines entwickeln sich unterschiedlich. Die Anlage der Parallelfaser-spines braucht keine synaptische Aktivität. Sie ist wahrscheinlich in der Entwicklung des Neurons intrinsisch vorgegeben. Das Wachstum von Kletterfaser-spines dagegen benötigt die Anwesenheit und Aktivität funktioneller Parallelfasersynapsen und wird andererseits durch die Aktivität der Kletterfaser gehemmt.
Rezeptoren und Ionenkanäle in Purkinjezellen
An den erregenden glutamatergen Kletter- und Parallelfasersynapsen exprimieren Purkinjezellen Glutamatrezeptoren. Bemerkenswerterweise verfügen reife Purkinjezellen nicht über NMDA-Rezeptoren. Die einzigen ionotropen Glutamatrezeptoren in Purkinjezellen sind AMPA-Rezeptoren. Da letztere über die GluR2-Untereinheit verfügen, ist ihre Calcium-Permeabilität gering.
Purkinjezellen exprimieren als einzige Neuronen im ZNS den GluRδ2-Rezeptor. Die Aminosäuresequenz dieses Rezeptors lässt vermuten, dass es sich um einen ionotropen Glutamatrezeptor handelt. Trotzdem wurde bisher weder eine direkte Bindung von Glutamat an diese Untereinheit, noch ihr Einbau in einen bekannten Glutamtarezeptor nachgewiesen. GluRδ2-Rezeptoren befinden sich vor allem an den Parallelfasersynapsen der Purkinjezellen. Bei Fehlen des Rezeptors kommt es zu Störungen der synaptischen Plastizität auf zellulärer Ebene und zu Störungen der motorischen Kontrolle und des motorischen Lernens auf Verhaltensebene. Viele andere postsynaptische Proteine stehen in Wechselwirkung mit der GluRδ2-Untereinheit. Der GluRδ2-Rezeptor hat also eine Schlüsselrolle für die Funktion des Kleinhirns, auch wenn seine Funktion und sein Wirkungsmechanismus noch ungeklärt sind.
Sowohl an Parallelfaser- als auch an Kletterfasersynapsen sind in Purkinjezellen perisynaptisch (also am Rand, nicht im Zentrum der Synapse) metabotrope Glutamatrezeptoren lokalisiert, und zwar überwiegend der Subtyp mGluR1.
Wie in allen Neuronen sind auch in Purkinjezellen spannungsaktivierte Natriumkanäle für die Entstehung und Weiterleitung von Aktionspotentialen zuständig. Sie werden in erster Linie im Soma und Axon der Purkinjezelle exprimiert. Im Dendritenbaum nimmt ihre Dichte mit zunehmendem Abstand vom Soma schnell ab. Aus diesem Grund dringt im Gegensatz zu anderen Neuronentypen wie zum Beispiel Pyramidenzellen im Hippocampus das Aktionspotential in Purkinjezellen nicht stark in den Dendritenbaum ein. In den Purkinjezellen von Säugetieren sind vor allem die Natrium-Kanäle Nav1.1, Nav1.2 und Nav1.6 exprimiert.
Im Dendritenbaum und Soma der Purkinjezellen finden sich spannungsabhängige Calciumkanäle, die zum größten Teil zum P/Q-Typ gehören. Diese bewirken bei starker Depolarisation (wie zum Beispiel einem Aktionspotential oder synaptischer Aktivität von Kletter- und/oder Parallelfasern) einen Einstrom von Calciumionen in die Zelle.
In der Membran des endoplasmatischen Retikulums (ER) von Purkinjezellen befinden sich ligandenaktivierte Calciumkanäle, und zwar sowohl IP3-Rezeptoren als auch Ryanodin-Rezeptoren. Beide setzen bei ihrer Aktivierung Calcium-Ionen aus dem ER-Calciumspeicher in das Cytosol frei und erhöhen dort die Konzentration sogenannter freier Calcium-Ionen. Von den verschiedenen IP3-Rezeptoren wird in Purkinjezellen in erster Linie der Subtyp 1 (IP3R1) exprimiert, und zwar im Vergleich zu anderen Zelltypen in mindestens 10facher Menge.
Spontanaktivität der Purkinjezellen
Purkinjezellen zeichnen sich durch eine hohe Spontanaktivität aus. Darunter versteht man, dass sie Aktionspotentiale unabhängig davon generieren, ob sie von Kletter- oder Parallelfasern erregt werden. Sie feuern mit einer Frequenz von ca. 50-150 Hz. Als Rhythmusgeneratoren dabei dienen vor allem Calcium-aktivierte Kaliumkanäle, die sogenannten BK-Kanäle[1].
Literatur
Weblinks
 Wikibooks: Topographische Anatomie: Neuroanatomie: Kleinhirn – Lern- und Lehrmaterialien
Wikibooks: Topographische Anatomie: Neuroanatomie: Kleinhirn – Lern- und Lehrmaterialien
Wikimedia Foundation.