- Translation (Biologie)
-
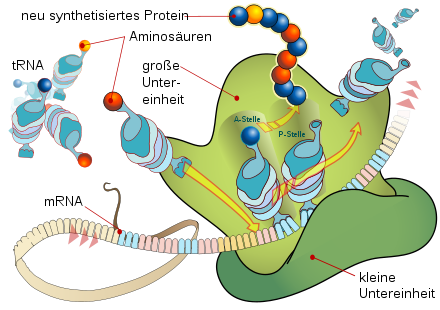 Schematische Darstellung der Translation am Ribosom
Schematische Darstellung der Translation am Ribosom
Als Translation wird die Synthese von Proteinen in den Zellen lebender Organismen (siehe auch Proteinbiosynthese) anhand der auf mRNA-Moleküle kopierten genetischen Informationen bezeichnet. Die Translation, als ein wesentlicher Teilprozess der Genexpression, ist der Transkription (komplementäre Kopie der DNA-Informationen auf einzelne mRNA-Stränge) nachgelagert, und erfolgt in lebenden Zellen an besonderen Strukturen, den Ribosomen.
Inhaltsverzeichnis
Allgemeiner Ablauf
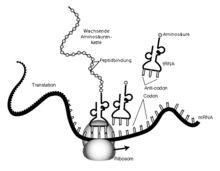 Translation im Cytoplasma; tRNA trägt Aminosäuren diese werden am wachsenden Peptidstrang im Ribosom angehängt.
Translation im Cytoplasma; tRNA trägt Aminosäuren diese werden am wachsenden Peptidstrang im Ribosom angehängt.Die nach dem genetischen Code eines Abschnitts der DNA gebildete mRNA ("m" für engl. "messenger", Bote) enthält Informationen zum Aufbau eines Proteins. Diese Information wird nun im Verlauf der Translation genutzt, um das entsprechende Protein zu synthetisieren. Es codieren jeweils drei aufeinanderfolgende Nukleotide der mRNA, die auch als Basentripletts oder Codons bezeichnet werden, eine bestimmte Aminosäure (siehe: genetischer Code). Das Protein wird sequentiell aus den hintereinander codierten Aminosäuren aufgebaut.
Für diesen Vorgang ist als Aminosäuren-„Transporter“ die tRNA ("t" für engl. "transfer", Übertragung) notwendig. Diese kann mit ihrem einen Ende, dem Anticodon, an dem passenden Codon auf der mRNA andocken und ist an ihrem anderen Ende durch die Aminoacyl-tRNA Synthetasen mit der genau zu diesem Codon passenden Aminosäure beladen.
Das Ribosom bringt die mRNA und eine tRNA, die eine Aminosäure aufgenommen hat, so zusammen, dass sich an ein bestimmtes Codon auf der mRNA – als passendes Gegenstück – ein komplementäres Anti-Codon der tRNA anlagert. Der Vorgang beginnt an der Stelle der mRNA, die das Startcodon mit der Sequenz AUG (Adenin, Uracil, Guanin) trägt. Eine zweite tRNA, die ebenfalls eine Aminosäure trägt, setzt sich neben der ersten tRNA an die mRNA. Die beiden an den tRNAs hängenden Aminosäuren werden mit einer Peptidbindung verknüpft, und die erste tRNA verlässt ohne Aminosäure das Ribosom. Die auf das nächste Codon passende tRNA lagert sich nun an die mRNA an. Ihre Aminosäure wird an die bereits bestehende Aminosäurekette geknüpft und erweitert so diese um ein weiteres Glied. Dieser Prozess setzt sich fort, so dass sich eine immer länger werdende Kette aus Aminosäuren bildet. Das Ribosom, das diesen Prozess katalysiert, wandert dabei immer um ein Codon auf der mRNA weiter, und zwar so lange, bis die Information der mRNA vollständig abgearbeitet ist. An dieser Stelle befindet sich ein Stopp-Codon (z.B.UGA) in der mRNA. An dieses kann keine der vorhandenen tRNA-Molekülarten andocken.
Dieses durch die Verkettung der Aminosäuren neu gebildete Eiweiß löst sich vom Ribosom und faltet sich dann meistens so, dass eine komplexe räumliche Struktur entsteht (Sekundärstruktur und Tertiärstruktur). Eventuell verbindet es sich noch mit anderen Proteinen zu übergeordneten Quartärstrukturen.
Eine mRNA wird in der Regel mehrfach abgelesen, bis sie durch Nucleaseaktivität in ihre Bausteine, die Ribonucleotide, zerlegt wird. Bei Eukaryoten ist die Haltbarkeit durch posttranskriptionelle Modifikationen im Kern erhöht.
Biochemischer Ablauf
Es existieren (beim Menschen) nur 41 verschiedene tRNA-Arten, was nicht äquivalent zu den 64 Tripletts ist. Die dritte Position des Codons ist bei vielen Aminosäuren variabel. Hierfür wird die Bezeichnung Wobble-Hypothese verwendet. Die tRNA hat in der Sekundärstruktur die bekannte Kleeblattstruktur, bedingt durch die intramolekulare Basenpaarung komplementärer Nucleotide. Dies entspricht aber nicht der dreidimensionalen Tertiärstruktur, welche eher dem Buchstaben L ähnelt.
Am Akzeptorarm sind die 5'- und 3'-Enden verknüpft. Am 3'-Ende, dem posttranskriptionalen CCA, bindet die entsprechende Aminosäure.
Die Anticodonschleife liegt in der Sekundärstruktur gegenüber, in der Tertiärstruktur hat diese den größtmöglichen Abstand zum Akzeptorarm. Drei zentrale Basen, in der Regel Nummer 34, 35 und 36 bilden das Anticodon. Wobei Position 34 die 1. Base des Anticodons ausmacht, welche mit der 3. Base des Codons paart.
Der D-Arm enthält das ungewöhnliche Dihydrouridin.
Der T-Arm enthält typischerweise Pseudouridin und Cytosin.
Die V-Schleife ist variabel, also bei den tRNAs unterschiedlich zusammengesetzt.
Die Beladung der tRNA mit einer Aminosäure erfolgt durch eine Aminoacyl-tRNA-Synthetase. Für jede Aminosäure gibt es eine spezifische Synthetase. Die genannten Basen gelten im Normalfall, sind aber auch variabel.
Ribosomen und Protein-Synthese
An den Ribosomen erfolgt die Bindung der Aminoacyl-tRNA an die Codons der mRNA und die Synthese der Proteine.
Bei den Ribosomen lassen sich zwei Untereinheiten (jeweils aus RNA und Polypeptiden bestehend) unterscheiden, die zunächst separat existieren. Bei der Translation vereinigen sie sich und bilden zwei funktionell bedeutsame Regionen, an denen sich die tRNAs binden können: an der Peptidyl-Stelle (P-Stelle) sitzt die tRNA mit der wachsenden Proteinkette, an der Aminoacyl-Stelle (A-Stelle) bindet die tRNA mit der nächsten anzufügenden Aminosäure. Eine weitere Region wird als Exit-Stelle (E-Stelle) bezeichnet, an der die entladenen tRNAs das Ribosom verlassen.
Initiation der Translation bei Prokaryoten
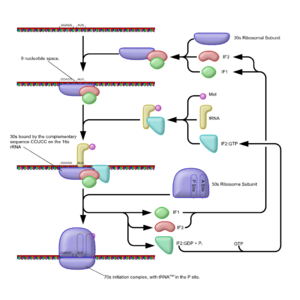 Schematische Darstellung der Initiation der Proteintranslation bei Prokaryoten
Schematische Darstellung der Initiation der Proteintranslation bei ProkaryotenBei der Initiation (Kettenstart) der Translation (bei Prokaryoten) spielen drei Initiationsfaktoren (IF 1, IF 2, IF 3) eine Rolle.
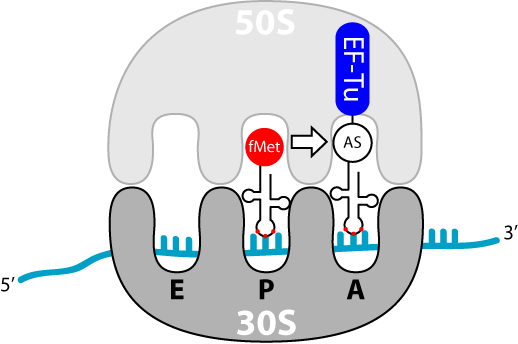
Die kleine Untereinheit (30S) bildet zu Beginn einen Komplex mit den Initiationsfaktoren 1 und 3[1]. Die Aufgabe des IF1 ist es, eine frühzeitige Bindung der beiden ribosomalen Untereinheiten zu verhindern während IF3 die Dissoziation der (in einem dynamischen Gleichgewicht liegenden) Nichtinitiator-tRNA bezweckt. Der IF2, ein G-Protein, bindet GTP, durchläuft eine Konformationsänderung und erhält dadurch die Möglichkeit die Initiator-fmet-tRNA zu binden. Dieser Komplex hat die Möglichkeit die mRNA und die kleine Untereinheit zu binden. Die Bindung erfolgt durch eine Interaktion der anti-Shine-Dalgarno-Sequenz der 16S-rRNA (ribosomale RNA und Teil der 30S Einheit) mit der Shine-Dalgarno-Sequenz auf der mRNA. Bei dieser handelt es sich um eine upstream (5' vom AUG-Codon gelegen), gelegene, nicht-codierende Sequenz. Dadurch erfolgt auch die Erkennung des Startcodons, durch die Initiator tRNA. Der Abschluss der Initiation wird durch GTP-Hydrolyse am IF2 eingeleitet. Es kommt zum Entlassen der Initiationsfaktoren und zur Bindung der 50S Untereinheit, wodurch der 70S-Initiator-Komplex entsteht. Die fmet-tRNA befindet sich zu Beginn der Translation bereits in der P-Stelle der 50S Untereinheit. Die beiden anderen Stellen, A und E, sind leer.
Elongation der Polypeptidkette
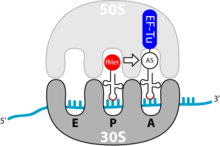 Elongationsschritte der Translation. (A=Aminoacyl- bzw. Erkennungsort, P=Peptidyl- bzw. Bindungsort). Die E-Stelle (E=Exit, Ausgang) des Ribosoms, dient der Positionierung entladener tRNA.
Elongationsschritte der Translation. (A=Aminoacyl- bzw. Erkennungsort, P=Peptidyl- bzw. Bindungsort). Die E-Stelle (E=Exit, Ausgang) des Ribosoms, dient der Positionierung entladener tRNA.Die Elongation ist der Prozess der Verlängerung der Aminosäurenkette; sie findet am Erkennungs- und am Bindungsort des Ribosoms statt. Ein einzelner Elongationsschritt enthält drei Schritte: Bindung der beladenen tRNA, Ausbildung der Peptidbindung und Vorbereitung auf den nächsten Elongationsschritt. Dies wiederholt sich so lange, bis ein terminierendes Codon erreicht ist.
Termination
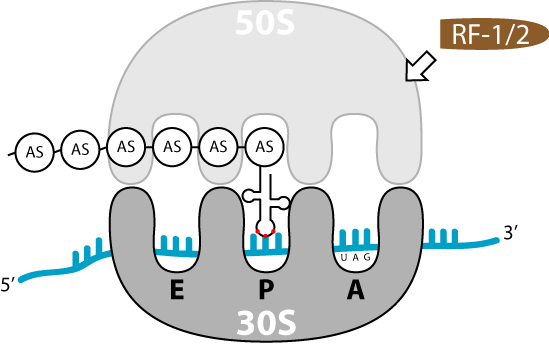
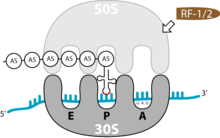 Terminationsschritte der Translation
Terminationsschritte der TranslationDas Ende der Translation ist erreicht, wenn auf der mRNA eines der Stopp-Tripletts UAG, UAA oder UGA auftaucht. Da es in der Zelle keine passende tRNA für diese Codons gibt, hält die Translation an.
Terminationsfaktoren (release factors) binden an das Stopp-Codon: RF1 an UAG und UAA oder RF2 an UAA und UGA. Ein weiterer RF 3-Faktor vermittelt die Interaktion zwischen RF1/ RF2 und dem Ribosom. Die Spaltung der Bindung zwischen der letzten Aminosäure und der letzten tRNA ist die interessanteste Funktion des Ribosoms. Der Ester kann nicht durch Hydrolyse aufgebrochen werden, da der Bereich der Peptidyl-Transferase vollkommen wasserfrei sein muss, um eine spontane Hydrolyse während der Elongation zu verhindern. Der Mechanismus dieser Reaktion lässt sich durch die Struktur des eukaryotischen Terminationsfaktor erklären. Die Struktur des prokaryotischen RFs ist noch nicht aufgeklärt. Man geht deshalb von einer hypothetischen Homologie aus. Der eukaryotische RF transportiert über eine bestimmte AS-Sequenz (Glycin-Glycin-Glutamin) genau ein Molekül Wasser in das Peptidyl-Transferase-Zentrum. Dieses kann dann, mit Hilfe katalytischer Aktivität des Ribosoms die Esterbindung spalten. Diese Sequenz befindet sich auch im prokaryotischen RF.
Nun fällt das Protein und die mRNA vom Ribosom ab, das wieder in seine beiden Untereinheiten zerfällt. Der Initiationsfaktor IF 3 erhält den dissoziierten Zustand aufrecht. Somit kann der Kreislauf von Neuem beginnen.
Translation in Eukaryoten
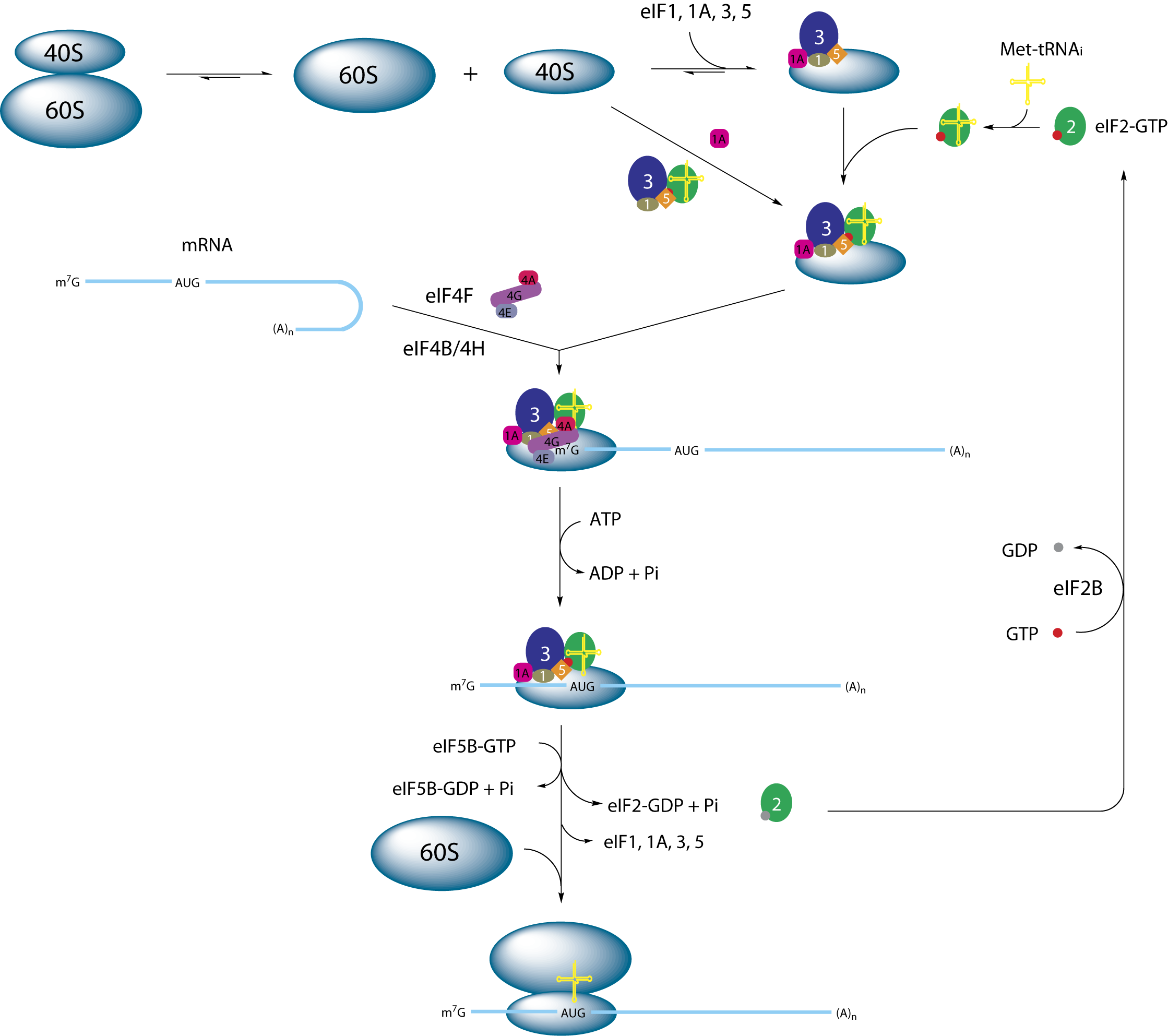
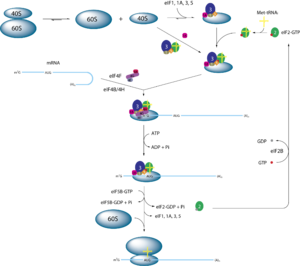 Schematische Darstellung der Initiation der Proteintranslation bei Eukaryoten
Schematische Darstellung der Initiation der Proteintranslation bei EukaryotenDie Translation in Eukaryoten unterscheidet sich besonders in der Initiation von der prokaryotischen Translation. Beteiligt sind daher spezielle eukaryotische Initiationsfaktoren. Es gibt keine fMet-tRNA, sondern eine Initiator-Met-tRNA, welche in vivo nicht formyliert ist. Außerdem gibt es keine Shine-Dalgarno-Sequenz auf der mRNA. Es wird gewöhnlich das erste AUG (5'-seitigste) der mRNA als Startcodon gewählt. Die Bindung der 40S Untereinheit erfolgt an der 5'-Cap-Struktur der mRNA. Daraufhin läuft das Ribosom in 3'-Richtung auf der mRNA entlang und sucht ein AUG. Den Erfolg der Suche "meldet" ein erfolgreiches Paaren der Initiator-Met-tRNA mit der mRNA. Des Weiteren hat die eukaryotische mRNA aufgrund der Prozessierung und des Transports aus dem Kern, genug Zeit komplexe Sekundärstrukturen zu bilden. Diese müssen durch Helicasen wieder aufgebrochen werden.
Translation durch Membranen
Da die meisten Bakterien beziehungsweise Prokaryoten von mehreren stabilen Membranen umgeben sind, haben sich hier einige besondere Mechanismen entwickelt, um Proteine durch Membranen nach außen zu synthetisieren. Auch bei Eukaryoten kommen solche Mechanismen vor, da auch die Organellen von einer (Doppel)-Membran umgeben sind. Es gibt zwei Vorgehensweisen:
Posttranslationaler Proteintransport
Bei diesem Verfahren wird das Protein in der Zelle vollständig zusammengebaut und durch Chaperone wird eine vorzeitige Auffaltung verhindert, wobei bei Bakterien durch einen eingebauten „Knick“ im Protein das Durchfädeln durch die Zellmembran erleichtert wird. Bei Eukaryoten konnte der posttranslationale Transport durch die ER-Membran bisher nur bei Hefen gezeigt werden.
Cotranslationaler Proteintransport
Bei diesem Weg wird das Ribosom durch Signalproteine an die Zellmembran (vor allem des Endoplasmatischen Reticulums) gebracht. Das entstehende Protein wird dann durch einen speziellen Tunnel in den dahinterliegenden Bereich geschoben.
Regulation
Jedes von der Zelle zum Überleben benötigte Protein ist in den Genen kodiert. Die benötigte Menge allerdings ist dabei nicht direkt im Gen kodiert und außerdem abhängig von Umgebungsbedingungen, Alter und Zellzyklus.
Unter der Regulation der Translation versteht man dementsprechend verschiedene biochemische Mechanismen, die die Translation, also Proteinbiosynthese steuern. Diese Steuerung erfolgt dabei ähnlich wie bei der Transkription: Ein Repressor bindet an den Translationsstartpunkt und blockiert so die Initiation dieses Prozesses. In einigen Fällen bezieht dies die Erkennung spezieller Strukturen an der mRNA ein. Ein wichtiger Signalweg, der durch Regulation der Translation Zellwachstum und Zellzyklus steuert, ist der mTOR-Signalweg.
Neben der Regulation der Translation hat die Zelle noch einige weitere Möglichkeiten, die exprimierte Proteinmenge durch Beeinflussung vorhergehender Glieder der Informationskette zu steuern:
- Regulation der Transkription
Bei eukaryotischen Zellen außerdem:
- Regulation des Transportes der mRNA aus dem Zellkern
- Alternatives Splicing
Beispiel einer Regulation der Translation ribosomaler Proteine
Die korrekte Expression ribosomaler Proteine stellt ein interessantes regulatorisches Problem für die Zelle dar. Jedes Ribosom enthält circa 50 spezielle Proteine, die alle mit derselben Rate synthetisiert werden müssen. Des Weiteren sind die Syntheserate von Proteinen der Zelle und der Bedarf an Ribosomen eng mit dem Zellwachstum verbunden. Eine Veränderung der Wachstumsbedingungen führt schnell zu einem Anstieg oder Absinken der Syntheserate dieser ribosomalen Komponenten. Dafür wird eine Regulation benötigt.
Die Kontrolle der Gene für die ribosomalen Proteine ist vereinfacht durch die Organisation in verschiedene Operons, die jeweils Gene für bis zu 11 ribosomale Proteine enthalten.
Die primäre Kontrolle geschieht auf der Ebene der Translation. Dies kann etwa durch das folgende Experiment nachgewiesen werden:
Durch Einbringen zusätzlicher Kopien eines solchen Operons in das Erbgut einer Zelle steigert sich dementsprechend die Menge der durch Transkription erzeugten mRNA. Trotzdem bleibt die Syntheserate des Proteins nahezu unverändert. Die Zelle kompensiert also die erhöhte mRNA-Menge. Dabei wirken ribosomale Proteine als Repressoren ihrer eigenen Translation.
Bei jedem Operon kann dabei ein schon synthetisiertes ribosomales Protein an die mRNA des Operons binden. Diese Bindungsstelle liegt in der Nähe eines der ersten Gene des Operons. Dadurch werden Ribosome daran gehindert, an die mRNA zu binden und mit der Translation zu beginnen. Die Repression der Translation der ersten Gene verhindert also die Expression eines Teils oder des gesamten Rests der nachfolgenden Gene.
Dieser Mechanismus ist sehr empfindlich. Schon wenige nicht zur Bildung von Ribosomen verbrauchte Moleküle des Proteins L4 zum Beispiel verhindern sowohl die Synthese dieses Proteins als auch der übrigen 10 ribosomalen Proteine im gleichen Operon. Dadurch wird also sichergestellt, dass die Proteine nicht in zu großen Mengen erzeugt werden, die nicht komplett zur Bildung von Ribosomen verbraucht werden können.
Wie ein Protein sowohl als ribosomale Komponente als auch als Regulator seiner eigenen Translation dienen kann, konnte durch Vergleich der Bindungsstellen des Proteins an der rRNA mit den Bindungsstellen mit seiner eigenen mRNA erforscht werden. Beide Bindungsstellen ähneln sich in ihrer Sequenz und ihrer Sekundärstruktur. Da die Bindung der ribosomalen Proteine an die rRNA stärker ist als die an die mRNA, wird die Translation nur unterdrückt, wenn der Bedarf an Proteinen für die Produktion von Ribosomen gedeckt ist.
Literatur
- Rolf Knippers: Molekulare Genetik. 9. komplett überarbeitete Auflage. Thieme, Stuttgart u. a. 2006, ISBN 3-13-477009-1.
Weblinks
- Lukas Hensel – Animation zur Translation
- Universität Bern – Translation bei Prokaryonten und Eukaryonten
- Zentrale für Unterrichtsmedien im Internet – Translation
- IUBMB: Prokaryotic and eukaryotic translation factors
Einzelnachweise
- ↑ James D. Watson u. a.: Molecular Biology of the Gene. 5. Auflage. Pearson/Cummings u. a., San Francisco CA 2004, ISBN 0-8053-4635-X, S. 435.

Wikimedia Foundation.