- Homologes Organ
-
 Homologie der Handknochen bei verschiedenen Säugetieren. I Mensch, II Hund, III Schwein, IV Kuh, V Tapir, VI Pferd (aus Gagenbauer 1870)
Homologie der Handknochen bei verschiedenen Säugetieren. I Mensch, II Hund, III Schwein, IV Kuh, V Tapir, VI Pferd (aus Gagenbauer 1870)Unter Homologie (griech. ὁμολογέω, homologeo, übereinstimmen) versteht man in der biologischen Systematik und der vergleichenden Anatomie die grundsätzlichen Übereinstimmungen von Organen, Organsystemen, Körperstrukturen, physiologischen Prozessen oder Verhaltensweisen aufgrund eines gemeinsamen evolutionären Ursprungs bei unterschiedlichen systematischen Taxa.
Die Cytogenetik kennt einen speziellen Homologie-Begriff, der von dieser Definition nicht erfasst wird: Auch innerhalb einer Art versteht man unter homologen Chromosomen jene Chromosomen, die die gleichen Gene enthalten. Bei zwei homologen Chromosomen in Organismen, die sich geschlechtlich fortpflanzen, wird dabei je eine Kopie vom Vater und eine von der Mutter vererbt. Bei Chromosomenvergleichen zwischen Arten wird 'homolog' im allgemeinen Sinn verwendet. Zum Beispiel ist das Chromosom 1 des Menschen zum Chromosom 1 des Schimpansen homolog, beide haben einen sehr ähnlichen Inhalt.
Die Ähnlichkeiten von homologen Merkmalen gehen evolutionär auf ein und dieselbe Struktur bei einem gemeinsamem Vorfahren zurück, können sich aber auseinander entwickelt haben und müssen nicht (mehr) die gleichen Funktionen erfüllen, das heißt sie sind bezüglich ihrer Herkunft äquivalent.
„Ein Merkmal zweier oder mehrerer Taxa ist homolog, wenn es sich von demselben (oder einem entsprechenden) Merkmal ihres nächsten gemeinsamen Vorfahren ableitet.“ (Ernst Mayr, 1997)[1]
Diese Merkmale spielen bei der Erstellung von Stammbäumen in der klassischen Systematik eine wichtige Rolle.
In der Genetik und Evolutionsbiologie wird der Begriff der Homologie auch auf Gene angewandt, die in unterschiedlichen Spezies ähnliche oder identische Funktionen haben und in ihrer Sequenz auf einen gemeinsamen Vorläufer zurückzuführen sind. In diesem Fall kann die Homologie zusätzlich in Paralogie und Orthologie unterteilt werden.
Erste "Ahnungen" von Homologie finden sich beim französischen Arzt Félix Vicq d'Azyr (um 1770), etwas später (wohl unabhängig) deutlicher bei Goethe (1790).[2] 1843 führte Richard Owen (1804-1892) den Begriff Homologie ein. Er war noch ein dem Schöpfungsgedanken verpflichteter britischer Paläontologe und Morphologe. Aufgrund der von ihm angenommenen Unveränderlichkeit der Arten könnten sich auch ihre kennzeichnenden Merkmale nicht verändern. Die Organismen seien nach dem Muster verschiedener Typen erschaffen worden; Merkmalsähnlichkeiten beruhten demnach auf „gewollten“ Variationen im göttlichen Baumuster dieser Typen. Mit der Erkenntnis der Veränderlichkeit der Arten, von Darwin 1859 veröffentlicht, wurde es notwendig, Owens ursprüngliche „Erklärung“ der Homologien zu korrigieren. In Unkenntnis der Vererbungsregeln wurde zunächst ein Konzept von abstrakten „Bauplänen“ postuliert, bis das Auftreten von Homologien schließlich durch die Erkenntnisse der Genetiker tatsächlich aufgeklärt wurde.
Inhaltsverzeichnis
Methode der Homologisierung
Grundannahme: Je ähnlicher verschiedene Organismen sind, umso größer ist ihre Verwandtschaft.
Da aber viele entfernt verwandte Organismen auf Grund einer ähnlichen Lebensweise auch sehr ähnlich aussehen (Konvergenz, siehe Analogie), sehr nah verwandte Arten aber in unterschiedlichen Lebensräumen sehr unterschiedlich aussehen können (Divergenz), ist es bei einem Vergleich von Organismen notwendig, von diesen Einflüssen zu abstrahieren: Ihre individuellen Baupläne werden durch "Homologisieren" auf abstrakte Baupläne zurückgeführt. Dabei werden gemeinsame Merkmalsausformungen, die lediglich Anpassungen an eine gemeinsame Lebensweise darstellen, eliminiert, die von der Lebensweise „verformten“ Merkmale (z.B Änderungen in der Größenrelation, Reduktion, Verschmelzung, Funktionsänderung, Ortsänderung im Gefügesystem) „entzerrt“ (siehe Homologiekriterien) und zu einem abstrakten „Bauplan“ vereinheitlicht, der als charakteristisch für die jeweilige Verwandtschaftsgruppe gilt.
Je näher diese betrachteten Organismen verwandt sind, umso detaillierter und komplexer ist ihr gemeinsamer Bauplan. Weit entfernt verwandte Organismen können nur auf einen einfachen, mit wenigen generellen Merkmalen ausgestatteten Bauplan zurückgeführt werden.
Auf der Suche nach dem fehlenden Glied (missing link) gemeinsamer Vorfahren können diese abstrakten Baupläne als Vorlage dienen.
Auf Grund der Komplexität der Bau- und Leistungsmerkmale von Organismen, konzentriert sich die Homologisierung in der Regel auf einzelne Organsysteme. Durch Heranziehen mehrerer Kriterien ergibt sich ein multidimensionaler Ähnlichkeitsraum. (In dem unten angeführten hypothetischen Beispiel wird nur ein Merkmal herangezogen!)
Ein Vergleich von Bau- und Leistungsmerkmalen heute lebender Arten und von Fossilfunden erlaubt ein Ordnen nach Ähnlichkeiten. Je weiter die Vergleichsobjekte voneinander entfernt sind, desto weniger gemeinsame Merkmale haben sie. Um später größere Sicherheit bei der Beurteilung des Verwandtschaftsgrades zu haben, müssen möglichst viele morphologische, anatomische, cytologische, physiologische, chemische, ontogenetische, ethologische Merkmale herangezogen werden. Dadurch ergibt sich ein multidimensionaler Ähnlichkeitsraum. (In dem unten angeführten hypothetischen Beispiel wird nur ein Merkmal herangezogen!)
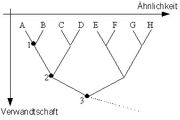 Modellstammbaum zur Methode der Homologisierung
Modellstammbaum zur Methode der HomologisierungAn den „Knoten“ dieses Stammbaumes sind die gemeinsamen Baupläne zu finden, die mit abnehmender Verwandtschaft immer abstrakter und merkmalsärmer werden. Homologiekriterien
Die drei Hauptkriterien (Lage, Struktur und Kontinuität) wurden 1952 von dem Zoologen Adolf Remane (1898-1976) vorgeschlagen. Anhand der solcherart festgestellten Homologien lasse sich auf die Verwandtschaft der Organismen rückzuschließen.
1. Kriterium der Lage
Strukturen sind dann homolog, wenn sie trotz unterschiedlicher Ausprägung in Gestalt und Anzahl in einem vergleichbaren Gefügesystem stets die gleiche Lagebeziehung aufweisen.
Beispiele:
- Innerhalb der Klasse der Insekten erfüllen die verschieden gestalteten Insektenbeine das Kriterium der Lage, denn sie sind alle am Thorax des Tieres angesetzt.
- Innerhalb der Klasse der Wirbeltiere gilt ähnliches für die Verdauungsorgane, die sich in Mund - Speiseröhre - Magen - Darm - After gliedern lassen.
- Ebenso der Aufbau des Herzens bei Säugetieren (linke Kammer, rechte Kammer, linker und rechter Vorhof sowie die ihre Anbindung an die Blutgefäße).
- Aufbau der Vordergliedmaßen von Landwirbeltieren. Der Grundbauplan besteht aus Knochen, die in der Abfolge von einem Oberarmknochen, zwei Unterarmknochen, mehreren Handwurzelknochen, fünf Mittelhandknochen und fünf Fingern wobei der Daumen zwei Fingerknochen hat und die restlichen vier jeweils aus drei Fingerknochen angelegt werden.
- Trotz identischer innerer Struktur kann das Aussehen und die Funktion der Vordergliedmaßen bei verschiedenen Wirbeltiereklassen sehr verschieden sein, denn sie dienen dem Laufen, Graben, Fliegen, Greifen oder Schwimmen. Im Verlauf der stammesgeschichtlichen Entwicklung haben sich Spezialisierungen der Vorderextremität herausgebildet. Erst der gleiche Grundbauplan weist auf gemeinsame Vorfahren hin, von denen aus sich die Entwicklung zu den heutigen Formen vollzogen hat.
2. Kriterium der spezifischen Qualität und Struktur
Ähnliche Strukturen können auch ohne Rücksicht auf die gleiche Lage homologisiert werden, wenn sie in zahlreichen Sondermerkmalen übereinstimmen. Die Sicherheit wächst mit dem Grad der Komplexität der verglichenen Struktur. Maßgeblich für das Homologiekriterium der spezifischen Qualität ist also der "innere Aufbau" eines Organs oder einer Struktur.
Beispiele:
- Nach dem Homologiekriterium der spezifischen Qualität sind der menschliche Zahn und die Hautschuppe eines Hais homolog, weil die äußerste Schicht aus Zahnschmelz und die darunterliegende Schicht aus Dentin besteht. Da sich Zähne wie Placoidschuppen als Spezifikationen eines urtümlich knöchernen Außenskeletts verstehen lassen, sind beide Strukturen tatsächlich homolog.
3. Kriterium der Kontinuität (Stetigkeit)
Organe sind dann homolog, wenn sie trotz unterschiedlicher Lage durch Zwischenformen, die sich homologisieren lassen, in Verbindung gebracht werden können.
Beispiele:
- Bestimmte Teile des Blutgefäßsystems (Halsschlagader) von Säugetieren lassen sich aufgrund von Embryonalstadien („Kiemenspalte“) mit den Kiemenbogenarterien der Fische homologisieren.
- die Schwimmblase von Knochenfischen und die Lunge von Landwirbeltieren sind homolog, da sich die Entwicklung von der Schwimmblase zur Lunge mit Hilfe der Amphibien und Sauropsiden nachvollziehen lässt.
Rezente Zwischenformen
Beispiel: Die Homologie des Articulare und Quadratum der Amphibien, Reptilien und Vögel mit Amboss und Hammer im Mittelohr der Säugetiere, sowie der hyomandibularen Columella der Vögel, Reptilien und Amphibien mit dem Steigbügel der Säugetiere wurde bereits 1837 von Carl Bogislaus Reichert (1811–1883) entdeckt.
Rudimente und Atavismen
Ein weiterer anatomischer Beleg für die Evolution sind Rudimente. Darunter versteht man rückgebildete Reste von Organen, die ihre Funktion verloren haben und dadurch überflüssig geworden sind. Zum Beispiel ist der Beckengürtel beim Bartenwal ein Rudiment. Erlangen diese Organe im Laufe der Zeit keine neuen Funktionen, können sie ganz verloren gehen: So ist die Blindschleiche keine Schlange, sondern eine Eidechse, deren Extremitäten vollständig zurückgebildet sind. In seltenen Fällen können rückgebildete Organe wieder auftreten, etwa beim Menschen der Fortsatz des Steißbeins als Schwanz. Man spricht in diesen Fällen von Atavismen.
Embryonale Zwischenformen
Während des embryonalen Entwicklungsprozesses werden Homologien zu spezifischen Arten sichtbar, die allerdings nur von ephemerer (vorübergehender) Dauer sind und beim endgültig entwickelten Lebewesen nicht mehr feststellbar sind. So hatte insbesondere Ernst Haeckel erkannt, dass jeder Mensch während seiner embryonalen Entwicklung eine vollständige, erdgeschichtliche „Mikroevolution“ durchläuft.
Beispiele:
- Der menschliche Embryo weist in einem bestimmten Entwicklungsstadium eine Kiemenspalte (Kiemenanlage) und damit eine unverkennbare Ähnlichkeit mit den Fischen auf, die später wieder verschwindet.
- Der Vogelflügel ist mit der Hand des Menschen homologisierbar, da der Vogelembryo noch eine Hand mit fünf Fingern aufweist, die im Laufe seiner Entwicklung durch Verschmelzung und Reduktion umgebaut werden.
Fossile Zwischenformen (auch fossile Brückentiere)
Anhand von gefundenen und restaurierten Fossilien lässt sich die schrittweise Entwicklung der Evolution belegen. Dies geschieht entweder durch Progressionsreihen oder Regressionsreihen, also eine nachvollziehbare Entwicklung von weniger komplexen Strukturen zu sehr komplexen Strukturen bzw. umgekehrt. Beispiel: Archaeopteryx. Der sogenannte „Urvogel“ zeigt den Übergang zwischen Landbewohnern und Vögeln. Es lassen sich sowohl landspezifische Entwicklungsreihen finden, als auch Entwicklungsreihen, die sich unverkennbar am Lebensraum der Luft orientierten.
Ethologische Übergänge
Auch ähnliche Verhaltensweisen können homolog sein, wie speziell die klassische ethologische Forschung herausgearbeitet hat.
Beispiele: das (angeborene) Balzverhalten innerhalb verwandter Vogelgruppen, etwa von verschiedenen Entenarten oder Hühnerartigen.
Homologie vs. Analogie
- Homolog sind Strukturen, die sich auf einen gemeinsamen Bauplan zurückführen lassen. Ihre unterschiedliche Ausprägung wird durch Divergenz erklärt. Ein Beispiel: Die Flosse eines Delfins und das Grabbein eines Maulwurfs sind bezüglich des Armskelettes homolog, da die Reihenfolge der Knochen, also Oberarmknochen, Elle und Speiche, Handwurzelknochen, Mittelhandknochen und Fingerknochen, gleich geblieben ist. Kurz: Gleicher Bauplan aber andere Funktion.
- Analog sind Strukturen, die sich nicht auf einen gemeinsamen Bauplan zurückführen lassen. Ihre ähnliche Ausprägung wird durch Konvergenz erklärt. Ein Beispiel: Die Flügel der Vögel und die Flügel der Fledermäuse sind bezüglich der Tragfläche – Federn bzw. Flughaut – zueinander analog (allerdings sie sind vom Skelett her homolog als Tetrapodenvorderextremitäten). Ebenso sind die Flügel von Insekten und Vögeln analoge bzw. konvergente Strukturen. Kurz: Anderer Bauplan durch anderen Ursprung, aber gleiche Funktion.
Die analogen Strukturen oder Verhaltensweisen erfüllen in den einzelnen Organismen den gleichen Zweck, sind also bezüglich ihrer Funktion äquivalent, jedoch nicht auf gemeinsame Vorfahren zurückzuführen. Vereinfacht ausgedrückt sind homologe Merkmale immer ursprungsgleich, analoge Merkmale lediglich funktionsgleich.
Problematik
Die Hauptproblematik des typologischen Homologiekonzepts besteht darin, dass die Richtung der auseinander strebenden Entwicklung homologer Organe erst aus ihrer Phylogenie verstanden werden kann. Es müsste also vor dem Erkennen der Homologie die phylogenetische Verwandtschaft bekannt sein.
- Ähnlichkeiten sind nicht immer auf enge phylogenetische Verwandtschaft zurückzuführen, es können auch Anpassungsähnlichkeiten (Analogien) sein (aus dem fehlenden Haarkleid bei Walen und Seekühen kann man nicht auf eine enge Verwandtschaft schließen, beide haben parallel eine thermische Isolation durch Fettschichten entwickelt).
- Unterschiede sind nicht immer auf entfernte phylogenetische Verwandtschaft zurückzuführen, sie können auch durch divergente Entwicklung von Schwesterarten in unterschiedlichen Lebensräumen, durch „Gendrift“ oder durch Neuerwerb von Merkmalen entstanden sein.
- Homologe Merkmale können im Laufe der Evolution soweit auseinander entwickelt werden, dass sie allein anhand eines Homologiekriteriums nicht gleich erkannt werden können. Dies tritt vor allem dann ein, wenn ein Merkmal bei der einen Art vollständig reduziert wird. Ohne Fossilfunde, Embryonalstadien, genetische Untersuchungen oder Zwischenstadien bei rezenten Arten ist dann schwer entscheidbar, ob die Abwesenheit dieses Merkmals primär oder sekundär ist. (So sind Nacktschnecken trotz des Fehlens eines Gehäuses mit den gehäusetragenden Schnecken eng verwandt, die gemeinsamen Vorfahren haben also ein Gehäuse getragen, welches die Nacktschnecken dann verloren haben.)
Verständnis der phylogenetischen Systematik
Merkmale sind dann homolog, wenn sie auf einen gemeinsamen Ursprung in der Phylogenese zurückzuführen sind. Homologie kann also erst dann festgestellt werden, wenn die Verwandtschaft bekannt ist.
1950 wird von Willi Hennig der Begriff der Homologie durch die Begriffe der phylogenetischen Systematik (Kladismus) ersetzt:
- Apomorphie liegt bei einem vergleichsweise abgeleiteten (in der Stammesgeschichte später erworbenen) Merkmal vor.
- Plesiomorphie liegt bei einem vergleichsweise ursprünglichen (also stammesgeschichtlich früher erworbenen) Merkmal vor.
- Autapomorphie ist der Neuerwerb von Merkmalen in einer Stammart (Beispiel: die Feder bei der Stammart aller Vögel).
- Synapomorphie ist das Auftreten von abgeleiteten Merkmalen einer Stammart bei ihren beiden Tochterarten und deren Nachfahren (Beispiel: Echoortung bei Zahnwalen und Bartenwalen).
- Symplesiomorphie ist das Auftreten von gemeinsamen Merkmalen bei verschiedenen Arten, das nicht auf deren letzte gemeinsame Stammart zurückzuführen ist, sondern auf eine noch ältere Stammart, die auch der Vorfahre anderer Arten ist (Beispiel: Haare bei Schimpanse und Mensch).
Die Bewertung der Merkmale ist also abhängig von der Lesrichtung (ursprünglich vs. abgeleitet) und Homologie (Rückführung auf einen gemeinsamen Grundplan): Das Merkmal "Neurocranium" ist eine Autapomorphie der Schädeltiere, eine Synapomorphie aller zu den Schädeltieren gehörenden Gruppen und eine Plesiomorphie für einzelne zu den Schädeltieren gehörende Gruppen. Der apomorphe Merkmalszustand ist an den Grundplan geknüpft, in dem er zuerst auftritt, in Grundplänen, die aus der weiteren Aufspaltung der resultierenden Gruppe hervorgehen ist dasselbe Merkmal dann im plesiomorphen Zustand.
Der Begriff des Bauplans wird durch den Begriff Grundplan oder Grundmuster ersetzt. Letzterer ist definiert als die Summe aller Plesiomorphien und Autapomorphien der letzten gemeinsamen Stammart einer monophyletischen Gruppe.
Zur Anwendung dieses Konzeptes siehe Kladistik und phylogenetische Systematik.
Anmerkungen
- ↑ Ernst Mayr: Das ist Leben - die Wissenschaft vom Leben, Spektrum Akademischer Verlag, Heidelberg, S. 189 ISBN 3-8274-1015-0
- ↑ Versuch die Metamorphose der Pflanzen zu erklären, gedruckt bei Carl Wilhelm Ettinger, Gotha, 1790
Weblinks


Wikimedia Foundation.