- Leishmanien
-
Leishmania 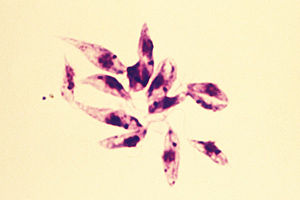
Leishmania tropica
Systematik Klassifikation: Lebewesen Domäne: Eukaryoten (Eucaryota) Stamm: Euglenozoa Ordnung: Kinetoplastida Familie: Trypanosomatidae Gattung: Leishmania Wissenschaftlicher Name Leishmania Ross 1903 Arten - Leishmania aethiopica
- Leishmania donovani
- Leishmania infantum
- Leishmania major
- Leishmania mexicana
- Leishmania amazonensis
- Leishmania tropica
Als Leishmanien (Leishmania) bezeichnet man eine Gattung von geißeltragenden Protozoen, die sich im Blut in Makrophagen vermehren (Hämoflagellaten). Leishmanien leben obligat als intrazelluläre Parasiten mit einem Wirtswechsel zwischen Insekten (Sandmücken, Schmetterlingsmücken) und Wirbeltieren (Schafe, Hunde, Menschen).
Das durch Leishmanien hervorgerufene Krankheitsbild bezeichnet man als Leishmaniose.
Inhaltsverzeichnis
Verbreitung
Sie sind bis auf Australien auf der ganzen Welt verbreitet und gelten als Auslöser vieler Tierseuchen. Der Mensch wird von weniger Arten befallen. Nach Schätzungen der WHO gibt es jährlich weltweit etwa 1,5 Millionen Neuerkrankungen an kutaner Leishmaniose und ca. 500.000 an viszeraler Leishmaniose. Etwa 12 Millionen Menschen sind zur Zeit mit Leishmanianen infiziert[1]
Merkmale
Wie bei allen Flagellaten, ändern sie ihre Form und die Position der Geißel je nach Wirt und Entwicklungsstadium (siehe: Trypanosomen). Jedoch sind die Leishmanien im Durchschnitt etwas kleiner (Promastigot: 20 Mikrometer, Amastigot: 2 µm).
Lebenszyklus
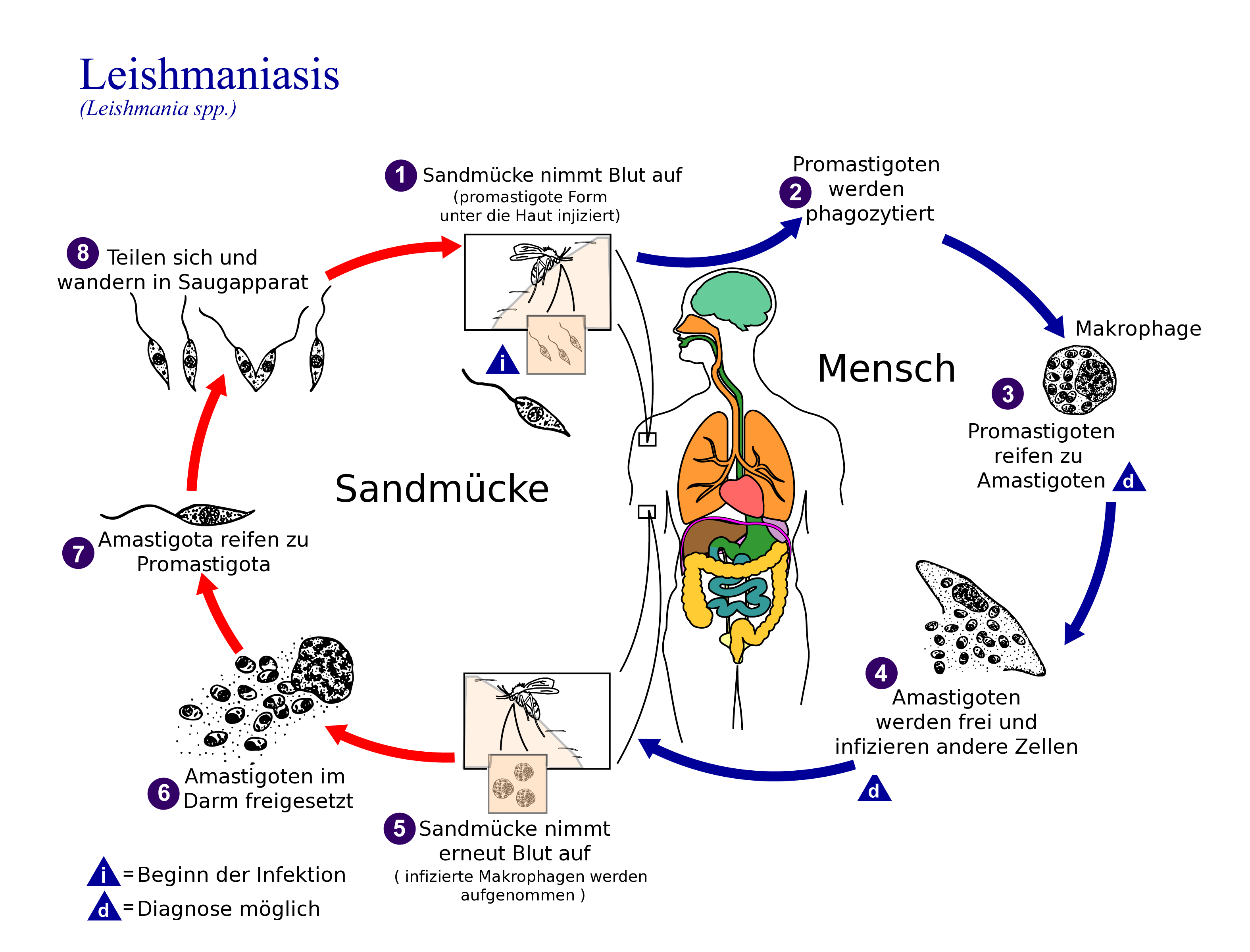
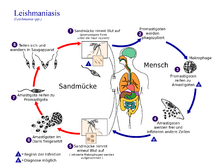 Lebenszyklus
Lebenszyklus
Die Vermehrung der Leishmanien läuft in zwei Wirten ab. Von der Sandmücke werden mit dem Speichel beim Stich promastigote (geißeltragende) Formen abgegeben, die im Wirbeltier von einem Makrophagen (Fresszelle) phagozytiert werden (passive Invasion) und sich dort in eine amastigote (unbegeißelte) Form umwandeln. In dem Makrophagen findet eine Vermehrung durch Teilung statt. Nach Zerstörung der Wirtszelle werden die amastigoten Formen frei und sind in der Lage, in andere Makrophagen einzudringen. Nimmt eine Sandmücke den Erreger aus dem Gewebe oder dem Blut auf, ist der Kreislauf geschlossen. Im Mückendarm wandeln sich die Leishmanien wieder zur promastigoten Form um. Diese wandert über eine abermals amastigote Form im Darmepithel nun in die Speicheldrüse der Mücke und eine Infektion kann erneut erfolgen.
Schadwirkung
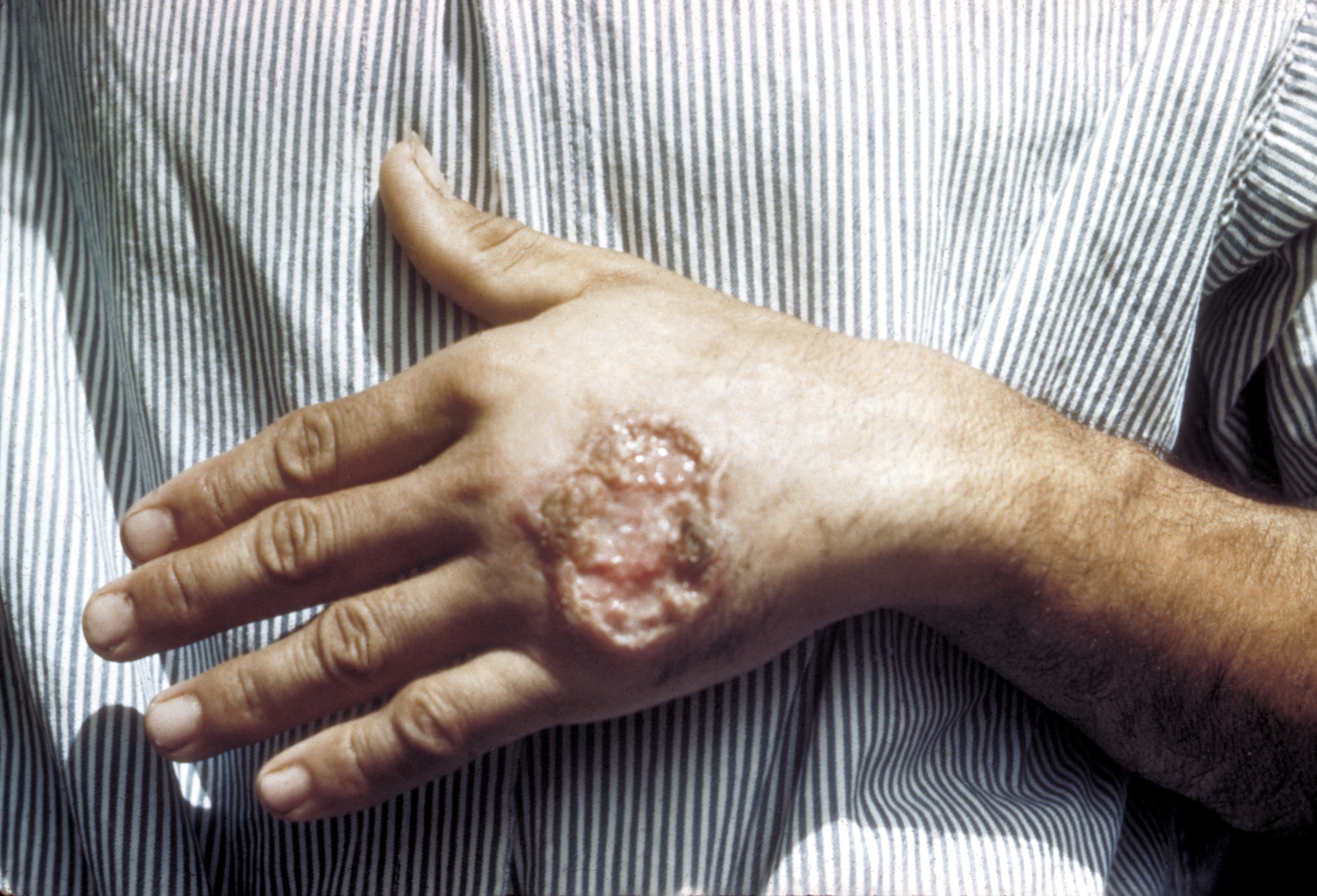
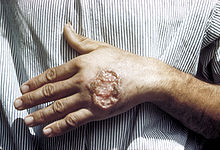 Kutane Leishmaniose
Kutane LeishmanioseBei den Krankheiten, die von Leishmania hervorgerufen werden (Leishmaniosen), unterscheidet man zwischen cutanen Formen („Orientbeule“) und viszeralen Formen („Kala Azar“). Bei der Orientbeule siedeln sich die Leishmanien bevorzugt in Makrophagen unter und im Hautgewebe an. Diese verläuft meist nicht tödlich. Wenn sie jedoch in innere Organe (Milz, Leber, Darm, Lymphknoten und Knochenmark) eindringen und dort Makrophagen befallen, kann es zu Hepatosplenomegalie-Symptomen (Krankhafte Vergrößerung von Milz und Leber) kommen, die unbehandelt in 90 % der Fälle durch Störungen der Blutgerinnung und zusätzliche Sekundärinfektionen tödlich verlaufen. Eine überstandene Leishmaniose führt zu einer lang andauernden oder lebenslangen Immunität.
Der Verlauf einer Leishmaniose ist sehr vom Immunstatus des Patienten abhängig. Bei HIV-Patienten gibt es häufig symptomarm verlaufende Infektionen (ohne Splenomegalie), die trotz Therapie und Rezidivprophylaxe eine mittlere Überlebensdauer von etwa einem Jahr zeigen. Die Häufigkeit von tödlich verlaufenden Infektionen ist besonders bei zusätzlicher Unterernährung sehr hoch.
Siehe auch: Parasiten des Menschen
Neutrophile Granulozyten – Trojanische Pferde für Leishmanien
Die Strategie des „Trojanischen Pferdes“ als Pathogenitätsfaktor von intrazellulären Mikroorganismen ist es, durch Makrophagen Clearance von neutrophilen Granulozyten, die durch Apoptose ein „keine Gefahr“-Signal auf ihrer Oberfläche tragen, die Immunabwehr und deren Gedächtnisfunktion geschickt zu umgehen.
Die von der Sandmücke übertragenen Parasiten der Art Leishmania major korrumpieren die Wirkung der ersten Abwehrreaktionen ihres Wirtsorganismus zu ihrem eigenen Nutzen und kehren sie von Fressen/Entzündung/Töten nach Fressen/keine Entzündung/kein Töten um. Sie machen sich die phagozytosefreudigen Polymorphonukleären Neutrophilen Granulozyten (PMN) entscheidend zu Nutzen für ein trickreiches Versteckspiel, in dem sie unerkannt vom Immunsystem die langlebigen Makrophagen befallen, um sich dort zu vermehren.
Aufnahme und Überleben
Bei einer Infektion mit Mikroorganismen im Gewebe treten PMN aus der Blutbahn aus und wandern, angelockt von Chemokinen, in das betroffene Gebiet (Haut nach Insektenstich) ein. Dort beginnen sie unverzüglich mit ihrer Arbeit als „erste Immunabwehrfront“ und phagozytieren die Eindringlinge aufgrund ihrer fremden, aktivierenden Oberflächenstrukturen. Dabei entsteht ein lokaler Entzündungsprozess. Aktivierte PMN sezernieren nun Chemokine, vor allem IL-8, die weitere Granulozyten anlocken und zur Phagozytose anregen. Zusätzlich fördern bei einer Leishmanieninfektion, bereits phagozytierte Leishmanien die Bildung von IL-8 durch PMN. Im Sinne der Parasiten klingt dies zuerst unlogisch. Dieser Mechanismus findet sich jedoch auch bei anderen obligat und fakultativ intrazellulären Mikroorganismen. Für solche Erreger gibt es mehrere Wege, innerhalb einer Zelle zu überleben. Überraschenderweise verursacht die Coinjektion von apoptotischen und lebenden Erregern einen weitaus fulminanteren Krankheitsverlauf als die alleinige Injektion mit lebenden Parasiten. Das normalerweise auf der Oberfläche von apoptotischen Zellen gefundene antiinflammatorische Signal Phosphatidylserin schaltet, bei Leishmania major vorkommend, den oxidativen Burst der Granulozyten aus, weshalb kein Töten und kein Abbau der lebenden Erreger erfolgt. Eine Vermehrung in den Granulozyten findet nicht statt, jedoch können die Erreger so unerkannt und daher unbekämpft im primär infizierten Gewebe persistieren. Weiterhin sind die promastigoten Formen der Leishmanien fähig, ein bestimmtes Chemokin zu produzieren, den Leishmania chemotactic factor (LCF), der die Chemotaxis für PMN steigert, nicht aber für andere Leukozyten (NK-Zellen, Monozyten). Dazu wird die Bildung des interferoninduzierbaren Chemokins CXCL 10 in den infizierten PMN gestoppt, was die Aktivierung von NK- und Th1-Zellen zusätzlich verhindert. Während der Phagozytose bleiben die Parasiten am Leben, da ihre primären Wirtszellen (PMN) durch die Ausprägung von apoptotic cell associated molecular patterns (ACAMP) „kein Erreger“ signalisieren.
Persistieren und Anlocken
Die natürliche Lebensdauer von Granulozyten ist relativ kurz und beträgt durchschnittlich 6 bis 10 Stunden im peripheren Blut, wonach sie sich einer spontanen Apoptose unterziehen. Bei infizierten PMN mit der Gattung Leishmania major konnte beobachtet werden, dass die Apoptoseanfälligkeit deutlich, durch eine Hemmung der Caspase-3-Aktivierung, um etwa 2 bis 3 Tage, verlängernd beeinflusst wird. Diese Tatsache ist besonders günstig für den Infektionsverlauf, da die eigentlichen Wirtszellen der Leishmanien, die Makrophagen, in welchen auch die Vermehrung und Entwicklung stattfindet, erst 2 bis 3 Tage nach den PMN in das Entzündungsgewebe einwandern. Die Parasiten lauern nicht nur tatenlos an ihrem Eintrittsort, sondern veranlassen die Granulozyten dazu, die Chemokine MIP-1α / CCL3 und MIP-1ß / CCL4 (macrophage inflammatory protein) auszuschütten, um damit die Makrophagen anzulocken.
„Silent Phagocytosis“
Um die Integrität des umliegenden Gewebes vor den Zellgiften und proteolytischen Enzymen aus den Granulozyten zu schützen, werden die apoptotischen PMN von Makrophagen abgeräumt. Das „iss mich“-Signal wird hier von Phosphatidylserin dargestellt, welches sich im Prozess der Apoptose auf die Außenseite der Zytoplasmamembran umlagert. Durch die verzögerte Apoptose werden die in den PMN persistierenden Leishmanien letztendlich über ein völlig physiologisches Abräumgeschehen von den Makrophagen aufgenommen. Der Weg über diese „leise Phagozytose“ hat nun folgende Vorteile für die Infektion:
- Die Aufnahme von apoptotischem Material dämpft die Makrophagenaktivität, wodurch auch keine Abwehrmechanismen gegen den intrazellulären Parasiten aktiviert werden und die Erreger überleben.
- Intrazellulär in PMN liegende Erreger haben keinen direkten Kontakt zu den Makrophagen-Oberflächenrezeptoren, da diese den Feind innerhalb der apoptotischen Zelle nicht „sehen“. Somit erfolgt keine Aktivierung der Fresszelle und des Immunsystems.
Quellen
- N. Suttorp, M. Mielke et al.: Infektionskrankheiten, Stuttgart (Thieme) 2004
- H. Hahn, D. Falke et al.: Medizinische Mikrobiologie und Infektiologie, 5. Aufl., Berlin, Heidelberg (Springer) 2004
Literatur
- Ger van Zandbergen u. a.: „Leishmania“ disease development depends on the presence of apoptotic promastigotes in the virulent inoculum. In: Proceedings of the National Academy of Sciences of the United States of America. PNAS. Vol. 103, No. 37, September 2006, ISSN 0027-8424, S. 13837–13842, doi:10.1073/pnas.0600843103.
- T. Laskay, G. van Zandbergen, W. Solbach: Neutrophil granulocytes – Trojan horses for „Leishmania major“ and other intracellular microbes? In: Trends in microbiology. Vol. 11, No. 5, Mai 2003, ISSN 0966-842X, S. 210–214.
Einzelnachweise
Weblinks
 Commons: Leishmanien – Sammlung von Bildern, Videos und AudiodateienKategorien:
Commons: Leishmanien – Sammlung von Bildern, Videos und AudiodateienKategorien:- Parasit
- Euglenozoen
- Veterinärmedizinische Parasitologie
- Medizinische Parasitologie

Wikimedia Foundation.