- Lineweaver-Burk Diagramm
-
Die Enzymkinetik ist ein Teilgebiet der biophysikalischen Chemie. Sie beschreibt, wie schnell enzymkatalysierte chemische Reaktionen verlaufen. Die Enzymkinetik findet breite Anwendung in Biologie und Medizin, da auch biologische Substrate (Reaktionspartner) – darunter solche, die im Menschen auftreten – untersucht werden. Ein Hauptziel der Enzymkinetik ist die Beschreibung der Konzentrationsabhängigkeit der Reaktionsgeschwindigkeit mit geeigneten Formeln, sowie die Bestimmung der dazugehörigen Parameter für ein bestimmtes Protein (Enzymaktivität und katalytische Effizienz). Da Enzyme dazu dienen, Reaktionen zu beschleunigen und zu lenken, ist die enzymkinetische Analyse zum Verständnis von Enzymfunktionen unerlässlich.
Der erste, der den Zusammenhang zwischen Substrat-Konzentration [S] und Umsatzgeschwindigkeit eines Enzymes v beschrieb, war der französische Physikochemiker Victor Henri 1902. Allerdings war die Bedeutung der Wasserstoffionenkonzentration für enzymatische Reaktionen damals noch nicht bekannt, erst nachdem Sørensen 1909 den pH-Wert definiert und die Pufferung eingeführt hatte konnten der Deutsche Leonor Michaelis und seine kanadische Post-Doktorandin Maud Menten 1913 die Ergebnisse Henris experimentell bestätigen. Die Henri-Michaelis-Menten-Gleichung wurde 1925 von G. E. Briggs und J. B. S. Haldane verallgemeinert (Michaelis-Menten-Theorie).
Henris Schlüsselidee war es, die enzymatische Reaktion in zwei Phasen zu zerlegen, die Bindung des Substrates an das Enzym und die Umsetzung des Enzym-Substrat-Komplexes in Enzym und Produkt.
Die Bindungsreaktion kann nach dem Massenwirkungsgesetz beschrieben werden, sie wird charakterisiert durch Dissoziationskonstante Kd = k − 1 / k + 1. Dabei ist Kd diejenige Konzentration des Substrates, bei der (in Abwesenheit der Umsetzung zu Produkt) die Hälfte aller Enzymmoleküle Substrat gebunden haben. Sie ist umgekehrt proportional zur Affinität des Enzymes für das Substrat.
Die Umsetzung des Enzym-Substrat-Komplexes zu Enzym und Produkt wird bestimmt durch eine Geschwindigkeitskonstante 1. Ordnung, kcat. Die tatsächliche Geschwindigkeit der enzymatischen Reaktion ergibt sich zu v = kcat * [ES]. Allerdings vermindert die Zerfallsreaktion die Konzentration von [ES], es ist deshalb erforderlich, diese statt mit Kd mit
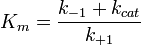 zu berechnen. Dann ergibt sich[1]
zu berechnen. Dann ergibt sich[1]Dies ist die Gleichung einer Hyperbel, sie nähert sich für unendlich hohe Substratkonzentrationen, wenn alle Enzymmoleküle Substrat gebunden haben, einem Grenzwert
![V_{max} = k_{cat} \times [E]_t](/pictures/dewiki/99/cd5e2778cc4fbd799d9a33d61c1ddcc7.png) an. Die Konzentration bei der die Reaktionsgeschwindigkeit gerade die Hälfte des Maximalwertes erreicht, ist die Michaelis-Konstante Km. Zur Bestimmung von Vmax und Km aus Meßreihen von v und [S] dienen computergestützte Verfahren wie die nichtlineare Regressionsanalyse (Simplex- oder Levenberg-Marquardt-Verfahren). Graphische Extrapolationsverfahren (Linearisierungen) wie etwa die doppelt-reziproke Auftragung nach Lineweaver und Burk sollten dafür nicht verwendet werden, da sie zu ungenau sind. Sie eignet sich jedoch sehr gut zur Präsentation der Ergebnisse enzymkinetischer Versuche, weil das menschliche Auge Abweichungen von einer Gerade leichter erkennen kann als die von einer Kurve.
an. Die Konzentration bei der die Reaktionsgeschwindigkeit gerade die Hälfte des Maximalwertes erreicht, ist die Michaelis-Konstante Km. Zur Bestimmung von Vmax und Km aus Meßreihen von v und [S] dienen computergestützte Verfahren wie die nichtlineare Regressionsanalyse (Simplex- oder Levenberg-Marquardt-Verfahren). Graphische Extrapolationsverfahren (Linearisierungen) wie etwa die doppelt-reziproke Auftragung nach Lineweaver und Burk sollten dafür nicht verwendet werden, da sie zu ungenau sind. Sie eignet sich jedoch sehr gut zur Präsentation der Ergebnisse enzymkinetischer Versuche, weil das menschliche Auge Abweichungen von einer Gerade leichter erkennen kann als die von einer Kurve.Inhaltsverzeichnis
Direkt-lineare Auftragung

Wenig bekannt ist die Tatsache, dass sich enzymkinetische Parameter bequem und präzise direkt aus einer Sättigungshyperbel gemäß der Abbildung herleiten lassen („direkt-lineare Auftragung“ auch „Cornish-Bowden-Diagramm“ genannt). In dieser Hyperbel ist die enzymatische Umsatzgeschwindigkeit v (Y-Achse) als Funktion der Substratkonzentration [S] dargestellt.
Für die direkt-lineare Auftragung überträgt man die Anfangsgeschwindigkeiten des enzymatischen Umsatzes direkt in das v-[S]-Diagramm. Die [S]-Werte sind vor Versuchsbeginn bekannt (eingestellte Substratkonzentrationen); während der Versuchsreihe ist dann der Ordinatenwert für v (die Anfangsgeschwindigkeit) nachzutragen. Im Gegensatz zu normalen Auftragungen werden die Messwerte also nicht als Punkte dargestellt, sondern in Form von Linien, die sich im günstigsten Fall in einem gemeinsamen Punkt schneiden, aus dessen Koordinatenwerten Km und Vmax folgen. Die katalytische Effizienz folgt übrigens aus der Steigung der Tangente an den Ursprung: Vmax/Km; daraus ergibt sich kcat/Km.
Die Fehlerbehandlung wird im direkt-linearen Plot weitgehend vereinfacht: die Linien im zweiten Quadranten werden mehr oder weniger um einen Punkt streuen. Mittelwertsbildung gibt dann die wahrscheinlichen Werte für die Parameter Km und Vmax. Bei Inspektion der Streubreite der Messpunkte (nicht identisch mit deren Standardabweichung) können Ausreißer leicht identifiziert und sogenannte Mediane abgelesen werden.
An dieser Stelle sei erwähnt, dass alle (auch die nachfolgenden) Auswertungverfahren nicht nur für Enzyme, sondern auch für die Bindungsvorgänge von Carriern oder Rezeptoren Gültigkeit haben. Historisch gesehen wurden all diese Methoden (Hanes und Eadie-Hofstee-Auftragung für Enzyme, Scatchard und Hill-Auftragungen für Carrier) ursprünglich von Woolf entwickelt, der aber seiner Zeit wohl etwas voraus war.
Linearisierungsverfahren
Linearisierungsverfahren wurden in der Vergangenheit sehr häufig für die schnelle grafische Bestimmung der wichtigen Kinetikparameter Km und Vmax verwendet. Sie sind zwar einprägsam und verbreitet, führen jedoch zu einer teils erheblichen Verfälschung des Ergebnisses durch Messfehler und sind zur Fehlerbetrachtung mehr oder weniger ungeeignet. Mittlerweile hat die Ermittlung der Michaelis-Menten-Parameter durch nichtlineare Regression stark an Bedeutung gewonnen, die zu deutlich genaueren Ergebnissen führt. Deshalb sollen die Linearisierungsverfahren hier nur gestreift werden.
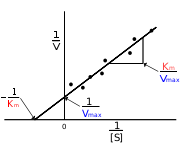 Lineweaver-Burke-Diagramm
Lineweaver-Burke-DiagrammLineweaver-Burk-Diagramm (doppelt-reziproke Auftragung)
- 1/v als Funktion von 1/[S].
zur Datenrepräsentation meist verwendet, zur Auswertung jedoch am wenigsten verlässlich. Kleine Fehler in v ergeben bei kleinen [S]-Werten eine große Abweichung in 1/v, bei großen [S]-Werten ist diese eher zu vernachlässigen. Die Autoren der Methode haben die Unsicherheit großer 1/v Werte betont und darauf hingewiesen, dass diese grundsätzlich geringer zu gewichten sind. Spätere Anwender haben dies zumeist ignoriert. Wo immer möglich sollte dieses Computerverfahren zur Bestimmung enzymkinetischer Parameter ersetzt werden.
Eadie-Hofstee-Diagramm
Das Eadie-Hofstee-Diagramm nimmt eine Mittelstellung ein. Hierbei wird v über (v/[S]) aufgetragen.
Eine Umformung der Michaelis-Menten-Gleichung ergibt die folgende Gleichung:
Aus dem Diagramm lassen sich vmax als Y-Achsenabschnitt und Km als negative Steigung der Regressionsgeraden ableiten.
Der Fehler wächst mit v/[S]. Da v bei beiden Koordinaten eingeht, konvergieren alle Abweichungen zum Ursprung.
Scatchard-Diagramm
Das Scatchard-Diagramm
- v/[S] als Funktion von v.
gleicht dem vorigen (Achsen sind vertauscht) und wird zumeist zur Repräsentation von Bindungsmessungen (anstelle enzymkinetischer Daten) angewendet. Scatchard- und Eadie-Hofstee-Diagramme gelten als die besten Werkzeuge zur Diagnose kooperativer Phänomene. Im Falle negativer Kooperativität oder nicht-identischer, isolierter Bindungsplätze entsteht ein konkaver Verlauf mit linearem Endast. Die Steigungen entsprechen hier den Affinitäten (Kd beziehungsweise Km) und die Gesamtzahl der Bindungsplätze (aktiven Zentren) ist aus dem Schnittpunkt mit der x-Achse abzulesen.
Hanes(-Wilkinson)-Diagramm (Hanes-Woolf-Diagramm)
- [S]/v als Funktion von [S].
Bestmögliche Auftragung dieses Typs. Fehler in [S]/v sind eine weit bessere Annäherung der Fehler in v. Aufgrund einer unverfälschten Spreizung der Messpunkte entlang der [S]-Achse wird das Ergebnis durch einzelne Ausreißer prinzipiell weniger verfälscht. Da aber abhängige und unabhängige Variable vermischt werden ist auch hier eine Datenoptimierung durch lineare Regression nicht sinnvoll. Der Anstieg der Regressionsgeraden beträgt 1/vmax, der Schnittpunkt mit der Y-Achse entspricht Km/vmax.
Hill-Diagramm
Graphische Methode, vorrangig für die Bestimmung eines kooperativen Bindungsvorganges für ein Protein (Enzym). Gebunden werden Substrate S (bzw. Liganden L) Die Graphik setzt Kenntnis von Vmax (beziehungsweise n) voraus. Die Auftragung von
- log v/(Vmax-v) als Funktion von log [S] beziehungsweise
- log r/(n-r) als Funktion von log [L]
ist eine Gerade der Steigung 1 (nH = 1), wenn die Bindungsplätze voneinander unabhängig sind. Bei kooperativen Systemen kann die Steigung am Nullpunkt (der Hill-Koeffizient nH) theoretisch der Zahl der Untereinheiten (n) gleichen, wird aber praktisch darunter bleiben. Falls die individuellen Km-(Kd-)Werte nicht durch Tangenten an die 45°- Ausläufer der Kurve extrapolierbar sind, ist nH ein Ersatzmaß der Kooperativität einzusetzen; nur für nH = 1 gleicht der Nulldurchgang ln Km (beziehungsweise ln Kd). Für Hämoglobin (n = 4), das als klassisches, hochkooperatives Protein gilt, wurde nH zu 2,8-2,9 bestimmt.

Inhibitoren
- Hauptartikel: Enzymhemmung
Viele Therapeutika und Gifte sind Hemmstoffe (Inhibitoren) von Enzymen. Aus diesem Grunde ist der Aufklärung des Wirkungsmechanismus immer eine besondere Bedeutung zugekommen. Die Nomenklatur der Hemmtypen wurde von W.W. Cleland 1963 auf eine systematische Grundlage gestellt, leider werden in vielen Lehrbüchern immer noch Begriffe abweichend verwendet.
Hier sollte allerdings beachtet werden, dass sich klassische Analysen auf reversibel bindende Stoffe beschränken. Irreversible Bindung einer Substanz an ein Enzym führt zur Inaktivierung, nicht zur Hemmung.
Abgeleitet aus der Michaelis-Menten-Gleichung (v = Vmax x [S] / (Km + [S]) stellt sich die allgemeine Inhibitionsgleichung wie folgt dar:
Danach kann das Verhältnis des Ki-Wertes (Dissoziationskonstante des Komplexes EI) und des Kii-Wertes (Dissoziationskonstante des Komplexes EIS) zur Ableitung des Inhibitionstyps dienen:
Kompetitiv
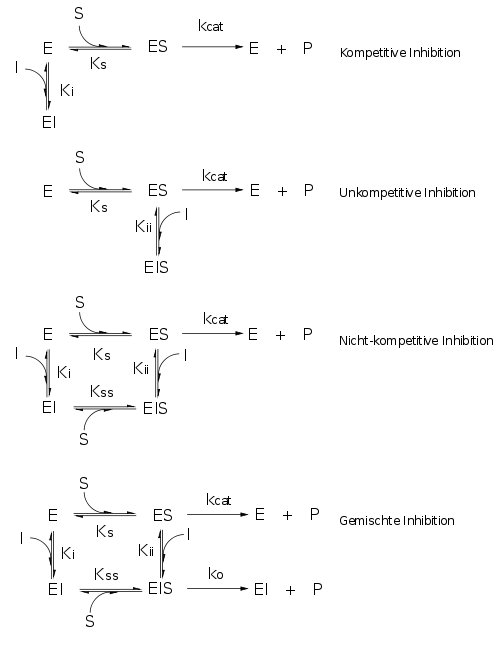
Inhibitor und Substrat schließen sich gegenseitig von der Bindung an das Enzym aus. Dies bedeutet jedoch nicht notwendigerweise, dass der Inhibitor an der gleichen Bindungsstelle bindet wie das Substrat. Auch wenn die Bindung von Substrat bzw. Inhibitor zu Konformationsänderung im Enzym führen, welche die Bindungsstelle für den jeweils anderen blockieren, ist die Hemmung kompetitiv. Wenn Substrat und Inhibitor allerdings die gleiche Bindungsstelle haben, dann ist der Hemmtyp notwendig kompetitiv.
Bei der Kompetitiven Hemmung kann der Inhibitor durch Substrat aus dem Enzym verdrängt werden, Vmax ändert sich also nicht. Allerdings wird für jede gewünschte Geschwindigkeit eine höhere [S] benötigt, die scheinbare Km wird also mit steigender [I] höher. Im Lineweaver-Burk-Diagramm führt dies zu einer Schar von Geraden, die einen gemeinsamen Schnittpunkt auf der y-Achse (1 / Vmax) haben.
Unkompetitiv
Der Inhibitor bindet nicht an das freie Enzym, sondern an den ES-Komplex. Höhere Konzentrationen des Substrates können daher den Hemmstoff nicht vom Enzym verdrängen, sondern führen zu vermehrter Bindung. Umgekehrt vermindert Bindung des Hemmstoffes die Konzentration von ES, nach dem Prinzip von Le Chatelier muss sich also zusätzliches ES aus E und S bilden: Die scheinbare Km vermindert sich, die Affinität des Enzymes für das Substrat steigt mit steigender [I]. Gleichzeitig nimmt natürlich Vmax ab. Im Lineweaver-Burk-Diagramm finden wir eine Schar paralleler Geraden.
Nicht-kompetitiv
Der Inhibitor kann sowohl an E als auch an ES binden. Im einfachsten Fall ist dabei Ki = Kii, d.h. das die Substratbindung die Affinität des Enzymes für den Inhibitor nicht verändert, etwa durch Konformationsänderung. Dann folgt natürlich auch, dass die Bindung des Inhibitors die Affinität des Enzymes für das Substrat nicht ändert und Ks = Kss. Wegen des Zusammenhangs zwischen Ks und Km ändert die Bindung von Inhibitor also auch nicht Km.
Es lässt sich nun zeigen (durch Substitution und Eliminierung aus den Definitionen von Ki,Kii,Ks und Kss), dass Ki / Kii = Ks / Kss. Wenn also Ki < Kii, dann folgt Ks < Kss und die scheinbare Km steigt mit [I]. Falls andererseits Ki > Kii, dann folgt Ks > Kss und die scheinbare Km sinkt mit steigendem [I].
Die nicht-kompetitive Hemmung führt im Lineweaver-Burk-Diagramm zu einer Schar von Geraden mit gemeinsamen Schnittpunkt links von der y-Achse, der Schnittpunkt liegt auf der x-Achse wenn Ki = Kii, er liegt über der x-Achse falls Ki < Kii und unter der x-Achse falls Ki > Kii.
Gemischt-kompetitive Hemmung
Der Mechanismus dieses Hemmtyps (der in der Praxis von geringer Bedeutung ist) ähnelt der nicht-kompetitiven Hemmung, allerdings hat der EIS-Komplex noch eine katalytische Aktivität. Auch das Lineweaver-Burk-Diagramm sieht aus wie bei der nicht-kompetitivben Hemmung (mit allen 3 Möglichkeiten). Im sog. Sekundärdiagramm (Steigung bzw y-Schnittpunkt im Lineweaver-Burk-Diagram als Funktion von [I]) sieht man aber im Falle der nicht-kompetitiven Hemmung Geraden, im Falle der gemischt-kompetitiven jedoch Kurven.
Einzelnachweise
- ↑ für eine Herleitung, siehe hier
Siehe auch
- Allosterie
- Crabtree-Effekt
- Energieladung
- exergon
- Fließgleichgewicht
- Mehrsubstratreaktion
- Multienzymkomplexe
- Pasteur-Effekt
- Substratzyklus
- Wechselzahl
Literatur
- H. Bisswanger: Enzymkinetik - Theorie und Methoden. 3. Auflage, Wiley-VCH, Weinheim 2000, ISBN 978-3527300969
- E. Buxbaum: Fundamentals of protein structure and function. Springer, New York 2007, ISBN 978-0-387-26352-6
- G. E. Briggs und J. B. Haldane: A Note on the Kinetics of Enzyme Action. In: Biochemical Journal. Band 19, Nr. 2, 1925, S. 338–229; PMID 16743508
- W. W. Cleland: The kinetics of enzyme-catalyzed reactions with two or more substrates or products. In: Biochimica et Biophysica Acta. Band 67, 1963, S. 104–137, 173–187, 188–196
- R. Eisenthal und A. Cornish-Bowden: The direct linear plot. A new graphical procedure for estimating enzyme kinetic parameters. In: Biochemical Journal. Band 139, Nr. 3, 1974, S. 715–720; PMID 4854723
- J. B. S. Haldane: Graphical methods in enzyme chemistry. In: Nature. Band 179, 1957, S. 832
- V. Henri: Theorie generale de l'action de quelques diastases. In: Comptes rendues l'Academie des sciences. Band 135, 1902, S. 916–919
- A. V. Hill: The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. In: The journal of physiology (London) Band 40, Supplement, 1910, S. iv–vii
- L. Michaelis und M. L. Menten: Die Kinetik der Invertin-Wirkung. In: Biochemische Zeitschrift. Band 49, 1913, S. 333–369
- I. H. Segel: Enzyme Kinetics. Wiley, New York 1975 (Nachdruck 1993)
- S. P. L. Sørensen: Enzymstudien II. Über die Messung und Bedeutung der Wasserstoffionenkonzentration bei enzymatischen Prozessen. In: Biochemische Zeitschrift. Band 21, 1909, S. 131–304
Weblinks
![\begin{align}v = k_{cat} [E]_t \frac{ [S]}{ K_m + [S]} = V_{max} \frac{ [S]}{K_m + [S]} \end{align}](/pictures/dewiki/49/1154f2bdb340d228c9c34cb47c079384.png)

![v = -K_m { v \over [S] } + V_{max}](/pictures/dewiki/57/9e236dce457252a814bf7544f5573a92.png)
![\nu =
\frac { V_{max} \times [S] }
{ Km (1 + \frac{[I]}{Ki}) + [S] (1 + \frac{[I]}{Kii}) }](/pictures/dewiki/55/75a1d92c0ac01d8bc750a55e591c5901.png)

Wikimedia Foundation.