- Mitotisch
-
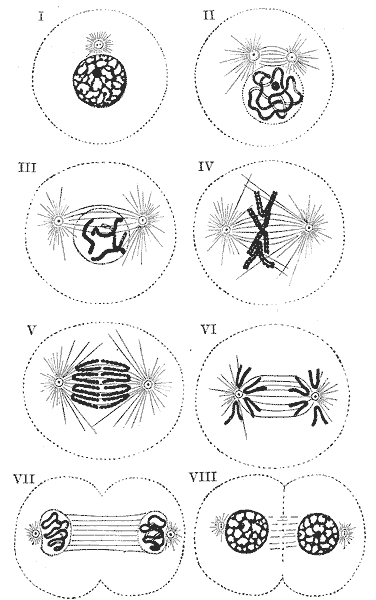
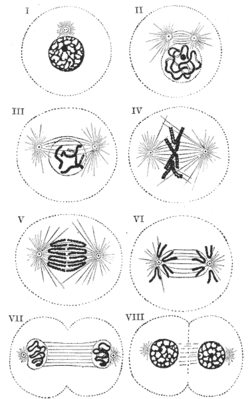 Schematische Darstellung der Mitose - I: Interphase; II: Prophase; III: Prometaphase; IV: Metaphase; V und VI: Anaphase; VII: Telophase; VIII: Zytokinese
Schematische Darstellung der Mitose - I: Interphase; II: Prophase; III: Prometaphase; IV: Metaphase; V und VI: Anaphase; VII: Telophase; VIII: ZytokineseAls Mitose (zu griechisch μίτος = Faden) (auch Karyokinese) bezeichnet man den Vorgang der Zellkernteilung bei Zellen eines eukaryotischen Lebewesens. Im Anschluss an die Kernteilung erfolgt meistens die Teilung des Zellleibs (Zytokinese), so dass aus einer Zelle zwei Tochterzellen entstehen. Mitose und Zytokinese werden auch als M-Phase (Mitose-Phase) zusammengefasst. Mit den jeweils zwischen zwei M-Phasen liegenden Interphasen bilden sie den Zellzyklus.
Während der Interphase werden die Chromosomen und die darin enthaltene DNA verdoppelt, so dass bei der Mitose identische Chromosomen auf die Tochterkerne verteilt werden können. Im Unterschied zur Meiose ändert sich der Ploidiegrad einer Zelle durch die Mitose nicht.
Inhaltsverzeichnis
Geschichte
Im Jahre 1835 beobachtete der Tübinger Botaniker Hugo von Mohl zum ersten Mal die Teilung einer Pflanzenzelle unter dem Mikroskop. In den folgenden Jahren sahen andere Zellforscher den gleichen Vorgang an den Zellen vieler Pflanzen und Tiere. Hugo von Mohl hatte im Nachhinein eine für das Verständnis der Lebensvorgänge wichtige Entdeckung gemacht. Der Berliner Arzt Rudolf Virchow drückte sie 1855 in einem Lehrsatz aus:
„Omnis cellula e cellula: Zellen entstehen nur aus Zellen.“
– Rudolf Virchow
Noch aber herrschten unklare Vorstellungen über den Feinbau der damals bekannten Zellbausteine und ihre Funktion. Dies betraf insbesondere den Zellkern und seine Rolle bei der Teilung. Erst mit der Weiterentwicklung der Mikroskope und der Färbetechniken in der zweiten Hälfte des 19. Jahrhunderts konnten die Forscher neue Erkenntnisse gewinnen. So sah der Bonner Botaniker Eduard Strasburger 1874 in einem Präparat sich teilender Zellen bisher unbekannte Teilungsstadien, in denen statt eines normalen Zellkerns längliche, gekrümmte oder abgewickelte Gebilde sichtbar waren. Wegen ihrer starken Anfärbbarkeit bezeichnete sie der Berliner Arzt Heinrich Wilhelm Waldeyer im Jahre 1888 als Chromosomen. Bei genauerer mikroskopischer Untersuchung stellte man fest, dass jedes Chromosom aus zwei gleichen Hälften besteht, den Chromatiden. Diese liegen eng aneinander, sind aber nur an einer Stelle, dem Centromer, miteinander verbunden.
Chromosomen entdeckte man nicht nur in Pflanzen-, sondern auch in Tierzellen. Im Laufe der Zeit fand man heraus, dass jede Pflanzen- und Tierart in allen Körperzellen eine arttypische Anzahl von Chromosomen besitzt. Die Anzahl liegt zwischen zwei Chromosomen beim Pferdespulwurm (Ascaris megalocephala univalens) und einigen hundert bei manchen Pflanzen.
Funktion der Mitose
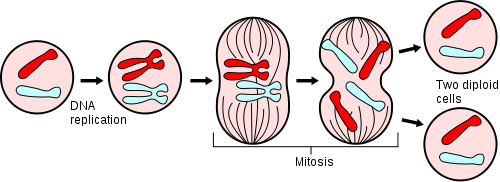 Schematische Darstellung des Zellzyklus. Zur besseren Veranschaulichung sind Chromosomen hier auch in Interphase so gezeichnet, wie sie in der Mitose aussehen. Dies entspricht jedoch nicht der Wirklichkeit.
Schematische Darstellung des Zellzyklus. Zur besseren Veranschaulichung sind Chromosomen hier auch in Interphase so gezeichnet, wie sie in der Mitose aussehen. Dies entspricht jedoch nicht der Wirklichkeit.Die Mitose ermöglicht, dass beide Tochterzellkerne die gleiche Anzahl an Chromosomen und damit die gleiche Erbinformation erhalten. Um das Erbgut einer Mutterzelle auf zwei Tochterzellen zu verteilen, werden die Chromosomen in der Interphase, einer weiteren Phase des Zellzyklus, verdoppelt. Ein Chromosom, das nach der Kernteilung zunächst aus einem Chromatid besteht, hat nach der Verdopplung zwei Chromatiden, die am Centromer zusammenhängen. Die Mitose wird durch sogenannte Mitogene ausgelöst.
Bei einzelligen Eukaryonten (Protisten) ist die Mitose zusammen mit der Zytokinese Grundlage der Vermehrung. Bei vielen Protisten verläuft die Mitose wie bei den mehrzelligen Eukaryonten als offene Mitose, d. h. die Kernhülle wird vorübergehend aufgelöst. Eine Ausnahme bilden die Dinoflagellaten, bei denen eine geschlossene Mitose, ohne Auflösung der Kernmembran stattfindet. Bei mehrzelligen Eukaryonten ist die Mitose die Voraussetzung für die Bildung eines neuen Zellkerns und somit üblicherweise für die Bildung neuer Zellen. Einige Ausnahmen von dieser Regel sind im Artikel über Zellkerne beschrieben. In mehrzelligen Organismen wie den Menschen findet die Zellteilung nicht mehr bei allen Zellen statt. So vermehren sich die Nervenzellen und Muskelzellen nach abgeschlossener Differenzierung nicht mehr. Hier verbleibt die Zelle in der sogenannten G0-Phase, so dass die DNA gar nicht erst repliziert wird (siehe Zellzyklus). Die roten Blutkörperchen können sich nicht mehr teilen, da ihr Zellkern fehlt und damit keine Mitose eingeleitet werden kann. Epidermalzellen hingegen vermehren sich wesentlich häufiger als der Durchschnitt.
Eine Mitose dauert bei menschlichen Zellen in der Regel etwa eine Stunde (im Vergleich dauert die Interphase von sich fortlaufend teilenden Zellen insgesamt durchschnittlich 24 Stunden). Bei Fliegen ist die Mitose teilweise nur 8 Minuten lang.
Eine Sonderform der Kernteilung vollziehen die Keimzellen: Sie entstehen durch eine in zwei Teilungsschritten ablaufende Teilung, die man Meiose oder Reifeteilung nennt und bei der aus einer diploiden Ausgangszelle vier haploide Zellen entstehen. Hier finden zwei Teilungsschritte statt, wobei die zweite Teilung ziemlich genau dem einer normalen Mitose entspricht.
Phasen der Mitose
 Einzelne Phasen der Mitose
Einzelne Phasen der MitoseDie Mitose wird in 4 oder 5 fließend ineinander übergehende Phasen eingeteilt. Die Prometaphase wird besonders in der englischsprachigen Literatur als eigenständige Phase gesehen, während in der klassischen deutschen Literatur die Metaphase direkt an die Prophase anschließt.
- In der Prophase der tierischen Zelle trennen sich die beiden Zentrosomen und wandern an entgegengesetzte Pole der Zelle. Zentrosomen wirken als Mikrotubuli-organisierende Zentren (engl. MTOC: Microtubule organising center) und sind der Ausgangspunkt der Mitosespindel. Die Zellen der höheren Pflanzen besitzen keine Zentrosomen, hier fungieren andere Zellbestandteile als MTOCs. Die Chromosomen (nur hier sind sie als das meist dargestellte "X" zu erkennen, in der Interphase sind sie mit Lichtmikroskopen gar nicht zu sehen) kondensieren und werden sichtbar. Sie bestehen aus je zwei Chromatiden, die nur am Zentromer zusammenhängen. Das Ende der Prophase ist erreicht, wenn die Kernhülle fragmentiert (englischsprachige Literatur) oder wenn die Kondensation der Chromosomen abgeschlossen ist (klassische deutsche Literatur)
- In der Prometaphase zerfällt die Kernhülle und die Spindelfasern dringen in den Bereich des jetzt aufgelösten Kerns ein. Die Chromosomen sammeln sich im Zentrum der Zelle. An den Zentromeren setzen die dreischichtigen Kinetochor-/Chromosomenmikrotubuli an, durch die die Chromosomen in der Metaphase ausgerichtet und in der Anaphase mit Hilfe der Polfasern auseinander gezogen werden können.
- In der Metaphase werden die kondensierten Metaphasechromosomen durch die Mikrotubuli der Spindel zwischen den Spindelpolen in der Äquatorialebene ausgerichtet. Die Metaphase ist abgeschlossen, wenn alle Chromosomen in dieser Metaphaseplatte angekommen sind.
- In der Anaphase werden die beiden Chromatiden eines Chromosoms durch die Spindelfasern (mit dem Zentromer voran) in Richtung Spindelpole auseinander gezogen. So erhält jedes Spindelfaserkörperchen einen vollständigen Chromatidensatz auf seiner Seite. Damit ist die Basis für die beiden zukünftigen Tochterzellen geschaffen. Die Anaphase gilt als beendet, wenn sich die Chromosomen der beiden zukünftigen Tochterzellen nicht mehr weiter auseinander bewegen.
- Als Telophase wird die letzte Phase der Mitose bezeichnet. Sie folgt übergangslos auf die vorausgegangene Anaphase. Die Kinetochorfasern depolymerisieren, die Kernhülle wird wieder gebildet und die Chromosomen dekondensieren. Nach Abschluss der Dekondensation befindet sich der Kern wieder in der Interphase.
- Auf die Telophase folgt in den meisten Fällen die Zytokinese, die Zellteilung. Diese ist jedoch nicht mehr Bestandteil der Mitose.
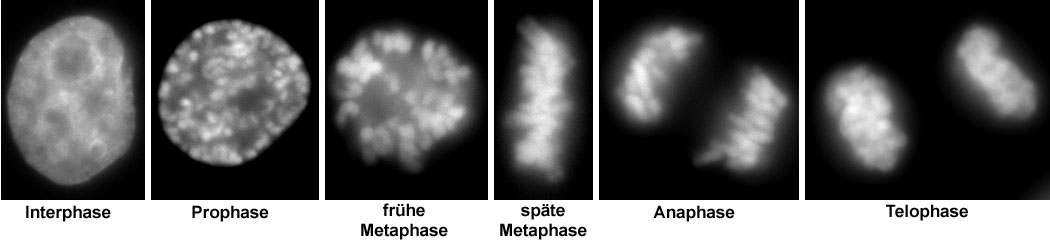
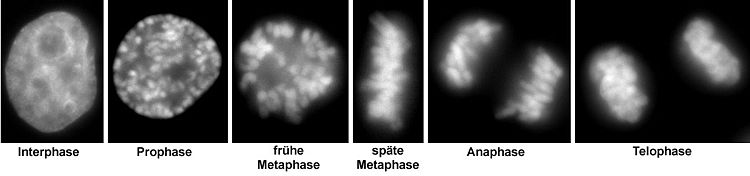 Neben einem zum Vergleich dargestellten Zellkern in der Interphase sind verschiedene Stadien der Mitose gezeigt (entsprechend der deutschen Literatur, daher ohne Prometaphase).
Neben einem zum Vergleich dargestellten Zellkern in der Interphase sind verschiedene Stadien der Mitose gezeigt (entsprechend der deutschen Literatur, daher ohne Prometaphase).Prophase
 Lichtoptischer Schnitt durch zwei Mauszellkerne in der Prophase. Durch die hohe Auflösung des verwendeten 3D-SIM-Mikroskops sind die kondensierten Chromosomen (rot) ungewöhnlich genau dargestellt. Die Kernhülle (blau) und Mikrotubuli (grün) sind durch Immunfärbung eingefärbt. Oben rechts ist ein Centrosom zu erkennen. Der Maßstabsbalken entspricht 5 µm.
Lichtoptischer Schnitt durch zwei Mauszellkerne in der Prophase. Durch die hohe Auflösung des verwendeten 3D-SIM-Mikroskops sind die kondensierten Chromosomen (rot) ungewöhnlich genau dargestellt. Die Kernhülle (blau) und Mikrotubuli (grün) sind durch Immunfärbung eingefärbt. Oben rechts ist ein Centrosom zu erkennen. Der Maßstabsbalken entspricht 5 µm.Im Anschluss an die Interphase und der damit fast abgeschlossenen Replikation der DNA kondensiert das Chromatin, so dass die Chromosomen sichtbar werden. Dabei falten und verdichten sich die Chromatide soweit, dass der genetische Code nicht mehr exprimierbar ist. Nun erkennt man die meistens dargestellte Chromosomenstruktur, mit dem Chromosomenpaar und dem zentralen Centromer. Zu diesem Zeitpunkt ist die Replikation noch nicht abgeschlossen.
Die Zentriolen organisieren die Teilungsspindel und bauen diese aus den Mikrotubuli auf, die sich zunächst sternförmig um die Zentriolen anordnen. Dabei wandert ein Zentriolenpaar auf die gegenüberliegende Seite und beide bilden so die Pole der Spindel. In dieser Phase löst sich zudem der Nucleolus (Kernkörperchen) auf.
Kondensation
Während der Interphase liegen die Chromosomen im Zellkern dekondensiert vor. Während der Prophase verdichten und verkürzen sich die Chromatinfäden durch Aufschraubung und Faltung. Es entstehen lichtmikroskopisch sichtbare Gebilde, die Kernschleifen oder Chromosomen. Sie sind nicht grundsätzlich neue Strukturen, sondern sie stellen nur eine kompaktere, für den Transport geeignete Form der Chromatinfäden dar. In diesem Zustand ist die DNA nicht mehr exprimierbar. Jedes Chromosom zeigt einen deutlichen Längsspalt. Es besteht nämlich aus zwei Längsstrukturen, den Chromatiden. An einer Einschnürungsstelle, dem Centromer, werden die Chromatiden zusammengehalten.
Polfaserbildung
Neben der Kondensierung der Chromatiden organisieren die Centriolen, welche sich ebenfalls in der S-Phase der Interphase verdoppelt haben, den Spindelapparat. Aus depolymerisierten Mikrotubuli des Cytoskeletts (siehe Weitere Ereignisse), den Tubulin-Untereinheiten, wird nun eine Spindel aufgebaut, die sich zunächst sternförmig um die Zentriolen anordnet. Man spricht hier auch von der Aster. Darauf hin wandert ein Zentriolenpaar auf die gegenüberliegende Seite des Zellkerns und definiert so die Pole. Schließlich erkennt man eine von einem Pol der Zelle zum anderen laufende Faserbildung, die sogenannten Polfasern. Diese spielen bei den späteren Verteilungsvorgängen eine Rolle.
Die Zentriolen sind nicht alleinige Zentren der Mikrotubuliorganisation. Sie bilden mit einer zu ihnen assoziierten amorphen Masse die Zentrosomen, welche auch für die Funktion der Spindel zuständig zu sein scheinen. Nach Zerstörung der Zentriolen durch einen Laser bleibt die Funktionalität der Spindel nämlich erhalten.
Pflanzenzellen benötigen selbst keine Zentriolen für ihre Zellteilung.
Weitere Ereignisse
Neben der Kondensation und der Polfaserbildung löst sich das Nucleolus auf. Außerdem depolymerisieren die Mikrotubuli des Zytoskeletts, woraufhin sich die Zelle abrundet.
Prometaphase
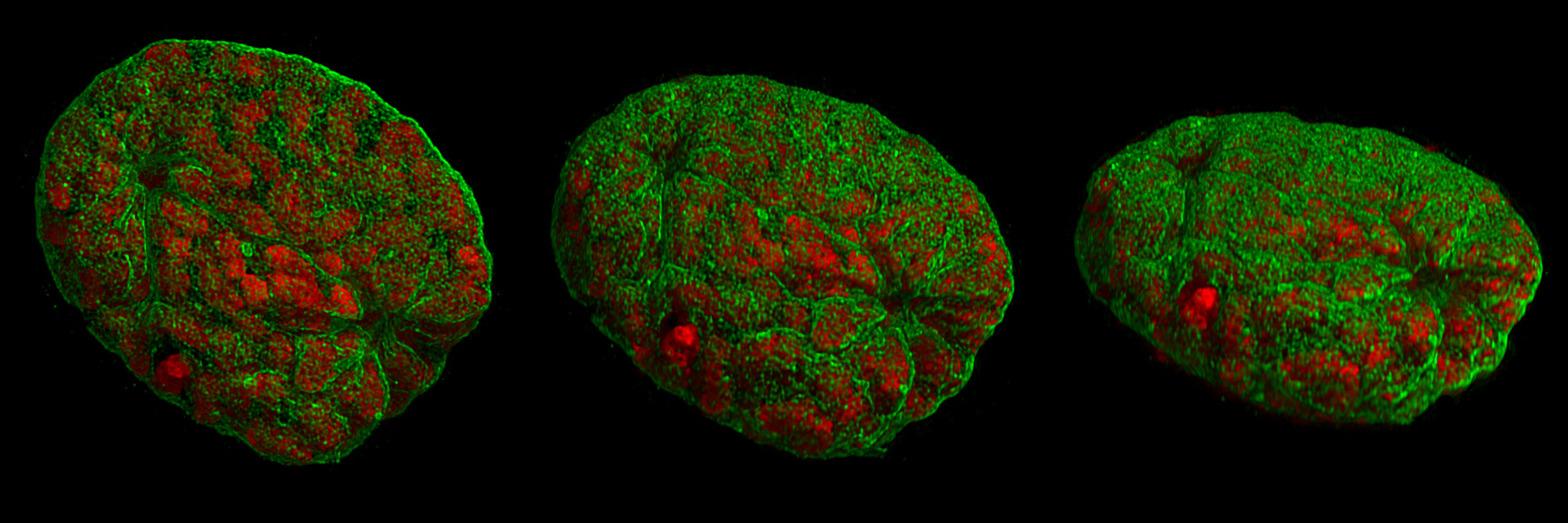
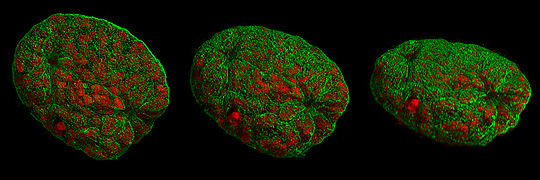 Dreidimensionale Darstellung eines Mauszellkerns aus verschiedenen Blickwinkeln während des Zusammenbruchs der Kernhülle. Die Chromosomen (rot) liegen bereits kondensiert vor. Durch die verbesserte Auflösung des verwendeten 3D-SIM-Mikroskops lässt sich am rechten Ende erkennen, wie die Kernhülle (grün) durch eindringende Mikrotubuli (nicht gefärbt) verformt wird. Die zweite Eindringstelle ist links oben zu sehen. Die Kernhülle zeigt im unteren Bereich in der Mitte einen Riss.
Dreidimensionale Darstellung eines Mauszellkerns aus verschiedenen Blickwinkeln während des Zusammenbruchs der Kernhülle. Die Chromosomen (rot) liegen bereits kondensiert vor. Durch die verbesserte Auflösung des verwendeten 3D-SIM-Mikroskops lässt sich am rechten Ende erkennen, wie die Kernhülle (grün) durch eindringende Mikrotubuli (nicht gefärbt) verformt wird. Die zweite Eindringstelle ist links oben zu sehen. Die Kernhülle zeigt im unteren Bereich in der Mitte einen Riss.Die Kernhülle beginnt sich nun durch Phosphorylierung der Lamine aufzulösen. Nachdem sich die Zentriolen auf den entgegengesetzten Polen organisiert haben, dringt die Spindel in das Nukleoplasma ein. An den Zentromeren der Chromosomen bilden sich nun dreischichtige Kinetochore, an die sich entsprechende Kinetochormikrotubuli anheften. Diese sind für den Transport der später getrennten Chromosomenteile zuständig und ordnen sich parallel zu den Polfasern an.
Auflösen der Kernhülle
Die Prometaphase beginnt mit der Auflösung der Kernhülle. Die Restteile sind kaum noch unterscheidbar zum endoplasmatischen Retikulum, da beide homologer Abstammung sind.
Bei einigen Eukaryoten Einzellern (Protozoa) bleibt die Kernhülle während des Kernteilungsprozesses enthalten, z.B. bei den Dinoflagellaten. Dort setzt sich die Spindel außerhalb der Kernmembran an und bildet dort kinetochorähnliche Strukturen.
Kinetochorfaserbildung
An den Zentromeren der Chromosomen bilden sich sogenannte dreischichtige Kinetochore. Diese speziellen Strukturen lösen eine Polymerisierung von Mikrotubuli aus, worauf sich in Richtung der Pole jeweils drei Kinetochorfasern bilden. Diese ermöglichen die Bewegung und Ausrichtung sowie Teilung der Chromosomen an der Stelle der Zentromere.
Die Chromosomen ordnen sich nun durch Orientierung an den Polfasern in der Äquatorialebene der Zelle an.
Metaphase
 Chromosomen (blau) und Spindelfasern (grün)
Chromosomen (blau) und Spindelfasern (grün)Die Chromosomen sind jetzt maximal verkürzt. Durch gleichmäßigen Zug des Spindelapparates richten sich die Chromosomen in der Äquatorialebene aus. Damit liegen sie quasi genau zwischen den Zentriolen in einer Ausgangsstellung, aus der heraus die Chromosomenpaare auseinander gezogen werden können.
Die Kinetochormikrotubuli liegen dabei genau parallel zu den Polfasern. Die Zugkräfte an den Chromosomen halten sich in beide Polrichtungen die Waage und werden so von der zentralen Position heraus langsam auseinandergezogen. Die Metaphase tritt genau in dem Moment in die Anaphase über, indem sich die Chromosomenpaare an der Zentromerstelle trennen und die beiden Tochterchromosomen zu den einzelnen Polen wandern.
Nach neueren Forschungen wird nun angenommen, dass nicht Zugkräfte von den Polrichtungen ausschlaggebend für das Auseinanderdriften der Chromatiden sind sondern Proteine an den Zentromerstellen, welche an den Mikrotubulifilamenten in Richtung der Zentromere wandern. Dieser Mechanismus funktioniert dann nach dem gleichen Prinzip, wie die Dynein- bzw. Kinesinproteine dem Mikrotubulus folgen.
Anaphase
Die beiden Chromatiden eines Chromosoms trennen sich nun voneinander. Die Tochterchromosomen, die nur noch aus einem Chromatid bestehen (Ein-Chromatid-Chromosomen) werden mit Hilfe der Zugfasern des Spindelapparates an den Centromeren zu den entgegengesetzten Polen der Zelle transportiert. Dabei verkürzen sich die Kinetochorfasern.
Gleichzeitig verlängern sich die Polfasern, wodurch sich die Pole voneinander abstoßen.
Chromatidenwanderung
Damit sich die beiden Chromatidenteile zu den Polen bewegen können, werden die Kinetochorfasern depolymerisiert. Durch die damit verbundene Verkürzung wandern die Kinetochoren an den Spindelfasern in die Richtung, in die die Dichte der Polfasern zunimmt. Die an die Kinetochoren über das Zentromer gebundene Chromatidenteile werden dabei wie kleine Ärmchen hinterhergezogen.
Anaphase I und Anaphase II
In der Anaphase wird zwischen dem Auseinanderrücken der Chromosomen (Anaphase I) und dem Auseinanderrücken der Spindelpole (Anaphase II) unterschieden.
Einleitung der Zellteilung
Gleichzeitig verlängern sich die Polfasern, mit dem Effekt, dass sich die beiden Polregionen, die sich in der Zelle gebildet haben, voneinander abstoßen und so die Voraussetzung der Zytokinese, also der eigentlichen Zellteilung geben. Die anschließende und für die Zellkernteilung abschließende Telophase tritt mit dem Erreichen der Chromosomen an den Polen ein.
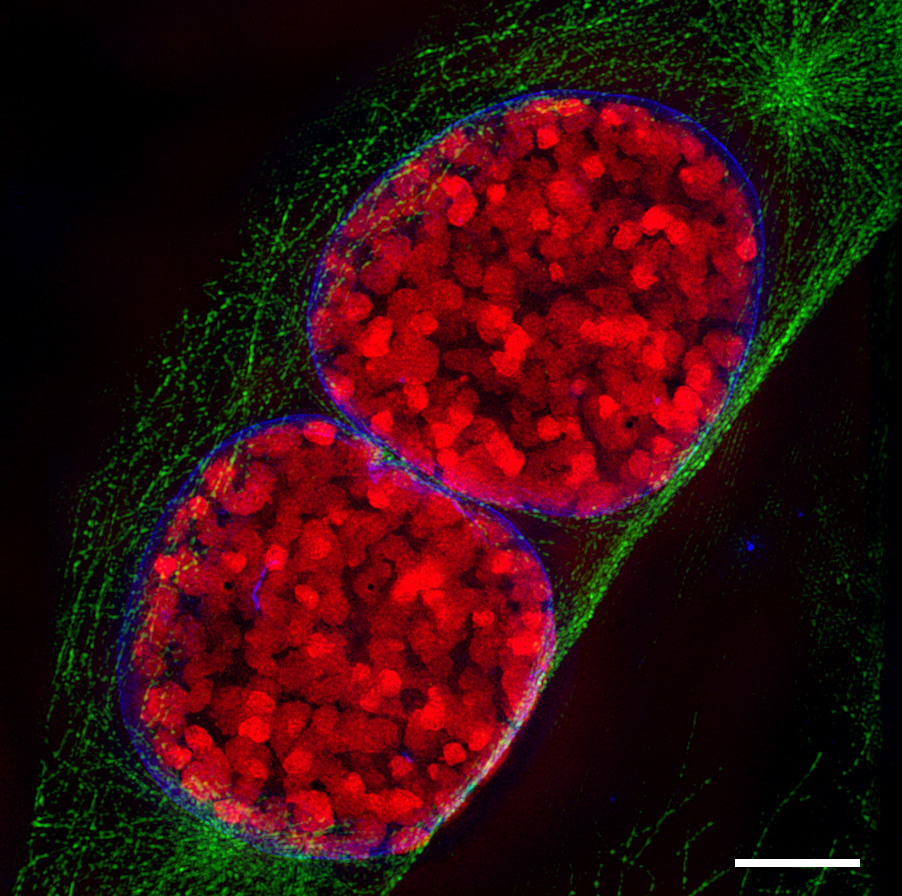
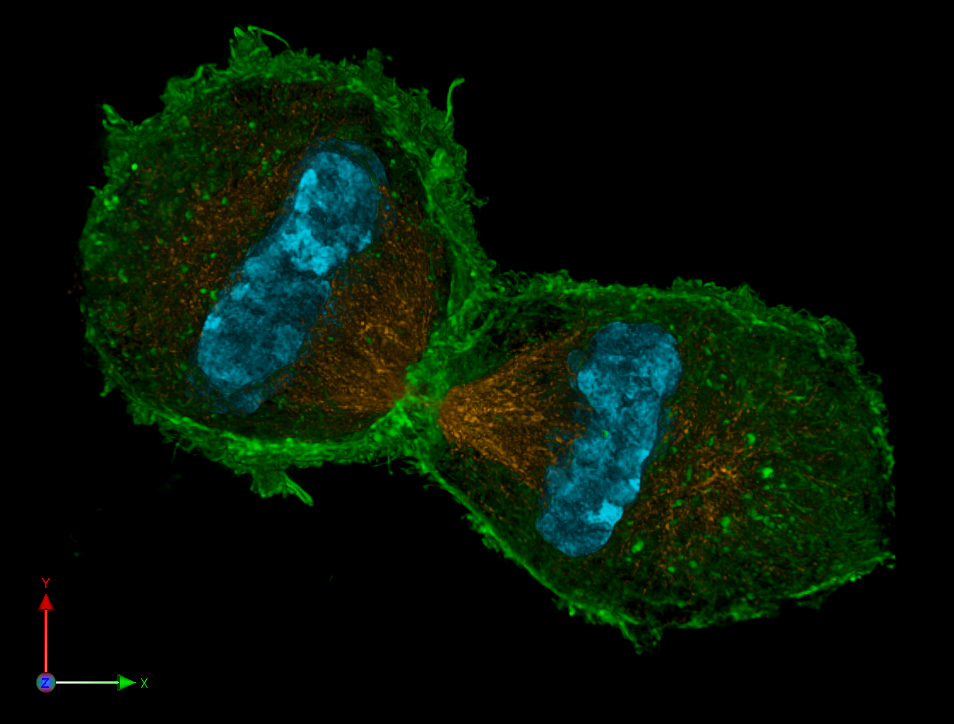
Telophase
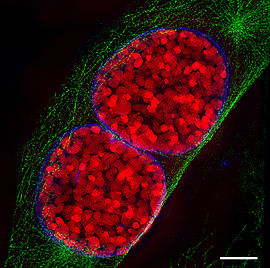
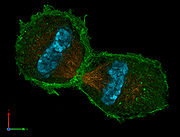 Dreidimensionale Darstellung zweier Tochterzellen in der Telophase. Zu sehen ist der Spindelapparat (anti-Tubulin-Immunfärbung; orange), Aktin-Zytoskelett (Phalloidinfärbung; grün) und das Chromatin (DAPI-Färbung; cyan).
Dreidimensionale Darstellung zweier Tochterzellen in der Telophase. Zu sehen ist der Spindelapparat (anti-Tubulin-Immunfärbung; orange), Aktin-Zytoskelett (Phalloidinfärbung; grün) und das Chromatin (DAPI-Färbung; cyan).Erreichen die Chromosomen die Pole, depolymerisieren die sich immer weiter verkürzten Kinetochorfasern schließlich. Die Kernhülle und Nukleolus werden gebildet und der Spindelapparat löst sich auf. Die Chromosomen entschrauben (dekondensieren) sich. Die eigentliche Teilung des Zytoplasmas und damit der Zelle wird durch die Zytokinese beschrieben.
Chromosomenwanderung
Die Chromosomen, die sich in der Anaphase getrennt haben, werden in der Anaphase und zu Beginn der Telophase zu den Polen der sich teilenden Zelle gezogen. Dabei verkürzt sich der Spindelapparat, indem die Untereinheiten der Kinetochor-Mikrotubuli, die sich in der Zentromer-Region an jedes Chromosom geheftet haben, offenbar am Kinetochor jedes Chromosoms abgebaut werden. Die Chromosomen selbst fangen nach dieser Depolymerisierung sofort an zu dekondensieren und sich zu entfalten.
Einleitung der Zytokinese
In der späteren Telophase sind die Chromosomen an den beiden Polen der Zelle angekommen und dekondensieren dort. Es wird um beide neuen Zellkerne eine Kernhülle aufgebaut, die zum großen Teil aus Fragmenten der alten Zellkernmembran der Mutterzelle bestehen.
Zudem verlängern sich die Polfasern noch weiter, bis die Pole die maximale Abstoßung voneinander erreicht haben und die Zellmembran in der Zytokinese eingeschnürt werden kann und so die zwei Tochterzellen entstehen lässt.
Im Anschluss an die Zellteilung treten die nun vollständig getrennten Tochterzellen wieder in die Interphase ein, der Zellphase, in der die normalen Zellfunktionen ausgeübt werden bis durch die Chromatidverdopplung eine neue Mitose angestoßen wird.
Die Mitose und der Zellzyklus
Die Zytokinese wird im allgemeinen nicht mehr zur Mitose selbst gezählt. In dieser Phase wird nicht die Kernteilung, sondern die abschließende Zellteilung betrachtet. Schon während der Telophase oder sogar Anaphase wird ein kontraktiler Ring aus Aktinfasern gebildet, der zusammen mit Myosin soweit verengt wird, bis die Plasmamembranen fusionieren und sich die Tochterzellen voneinander trennen.
Im Anschluss an die vollständige Zellteilung kann sich der diploide Ein-Chromatid-Chromosomensatz in der Interphase wieder zur einem diploiden Zwei-Chromatid-Chromosomensatz replizieren, um eine neue Mitose anzustoßen.
Besonderheiten
Im Anschluss an die Mitose muss nicht in jeden Fall eine Zytokinese stattfinden. So durchläuft der Zellkern der Wimpertierchen (Ciliata) in der Konjugation zwei Mitosen, ohne das Zellplasma zu teilen.
Bei den Geißeltierchen (Flagellata) kommt es vor, dass sich der Zellkern bis zu 8 Mal teilt, so dass eine Zelle mit 16 Zellkernen vorkommt. Solch eine Zelle wird auch Plasmodium genannt. Erst dann wird die Zelle in einem Schritt in 16 Tochterzellen getrennt.
Plasmodien der Schleimpilze können sogar Tausende von Zellkernen aufweisen. Einige dieser Ansammlungen kommen aber eher durch Zellverschmelzungen oder Aggregationen zustande. In diesem Fall spricht man eher von einem Synzytium, einer Riesenzelle, deren Zellkerne von vielen verschiedenen Zellen stammen und nicht von einer Urzelle.
Siehe auch
Weblinks
- Die Mitose im Zusammenhang der Zellbiologie – Grafiken
- Vergleich Mitose und Meiose
- Zell- und Kernteilung – Mitose
- Bilder, Zeichentrick
- Lern- und Informations- System Botanik - Mitosedarstellung an einer Pflanzlichen Zelle
- Online-Biologie Kurs
- Darstellung der Mitose
- Darstellung der Meiose
- eingefärbte Zellteilung (gelb: Zentrosomen, blau: Chromosomen, grün: Mikrotubuli)

{kind=link}
Wikimedia Foundation.