- Wasseraffentheorie
-
Unter dem Begriff Wasseraffen-Theorie (Bezeichnungen wie Wassertheorie, Wasseraffen-Hypothese oder Aquatic-Ape-Hypothese wurden auch verwendet) ist eine Reihe von Spekulationen bekannt geworden, nach der die Vorfahren der Menschen in ihrer Evolution – der Hominisation – eine teilweise aquatische (wasserlebende) Phase durchgemacht haben sollen.
Das bedeutet, dass Vormenschen sich verstärkt an und in Flüssen, Seen, Meeren oder am Litoral aufgehalten haben und diese u. a. zur Nahrungsgewinnung nutzten. Nach dieser Phase – die zur Entwicklung einiger besonderer Merkmale führte – seien sie wieder stärker zum Leben an Land übergegangen. Die Hypothese besagt jedoch nicht, dass diese Vorfahren ausschließlich im Wasser gelebt hätten.
Merkmale, die mit Elementen dieser Hypothese erklärt werden können, sind insbesondere
- der aufrechte Gang;
- die verlängerten hinteren Extremitäten;
- die besondere Struktur der Haut und des Unterhaut-Fettgewebes im Vergleich zu Menschenaffen;
- der im Vergleich zu anderen Landsäugern deutlich höhere Fettanteil des Menschen;
- die eigentlich strömungsungünstig nach unten offene Nase;
- der Tauchreflex (Verlangsamung des Herzschlages);
- der Stimmritzenkrampf als Schließreflex des Kehlkopfes;
- die Kontrolle der Atmung beim Tauchen als Voraussetzung zur Sprachentwicklung.
- genetisch veranlagte Affinität zum Lebensraum Wasser
- die überproportionale Anzahl von fossilen Funden in Wassernähe
- die für ein ausschließlich an Land lebendes Säugetier viel zu großen Füße
Die erste Wasseraffen-Theorie: Westenhöfers Aquatile Hypothese
Einige der Hauptargumente der Wasseraffen-Theorie wurden bereits ab 1923 durch den Berliner Pathologen Max Westenhöfer (1871–1957) aufgestellt; eine Zusammenfassung seiner Ideen sind in seinem 1942 publizierten Buch Der Eigenweg des Menschen zu lesen. Er glaubte, dass in der Hominisation eine aquatische Phase stattgefunden habe, und belegte dies anhand anatomischer Vergleiche. Allerdings muss man anmerken, dass er eine Abstammung oder nahe Verwandtschaft von Menschen zu Menschenaffen ausschloss und stattdessen den Menschen als direkten (weil ebenso wenig spezialisierten und im Feuchten lebenden) Nachkommen von Lurchen und Salamandern sah.[1]
Hardys Aquatic Ape Theory und die Weiterentwicklung dieses theoretischen Modells
Vermutlich unabhängig von Westenhöfer kamen Sir Alister Hardy um 1930 die ersten Gedanken zu diesem Konzept. Er las eine Veröffentlichung, in der Besonderheiten des menschlichen Unterhaut-Fettgewebes beschrieben wurden. Dieses ist – im Unterschied zu den meisten Säugetieren – so fest mit der Haut verbunden, dass es mit angehoben wird, wenn man an der Haut zieht. Hardy, ein anerkannter Meeresbiologe, assoziierte damit die Verhältnisse bei Walen und überlegte, dass der Mensch in seiner Vorgeschichte eine aquatische Phase gehabt haben könne. Im Laufe der Jahre sammelte er weitere Indizien, zögerte aber mit der Veröffentlichung.
Erst im März 1960 publizierte Hardy seine Gedanken in dem Artikel Was man more aquatic in the past in der populärwissenschaftlichen Zeitschrift New Scientist. Hardy hat kein Buch über das Thema veröffentlicht.
Diese Theorie wurde 10 Jahre später durch die englische Journalistin Elaine Morgan, aufgegriffen und populärwissenschaftlich und in Verbindung mit feministischen Ansichten weiterentwickelt. Morgan veröffentlichte mehrere Werke zu dem Thema. Weitere Anhänger der AAT sind unter anderen Marc Verhaegen, Erika Schagatay, Renato Bender, Nicole Bender-Oser und Algis Kuliukas.
In der akademischen Welt wurde 1987 in Valkenburg (Niederlande) das erste Symposium zu dieser Hypothese durchgeführt, bei dem Verfechter und Gegner ihre Ansichten austauschten. Das zweite Symposium fand 1999 unter dem Titel „Water and Human Evolution“ in Gent (Belgien) statt.
In jüngerer Zeit fanden die Argumente der Wasseraffen-Theorie bei Chris Stringer (British Natural History Museum) und David Attenborough (u. a. Autor populärwissenschaftlicher Produktionen der BBC zur Hominisation) Beachtung.
Grundprobleme von Hominisations-Hypothesen
Generell unterliegen Hypothesen zur Erklärung der spezifischen Merkmale des Menschen im Verlauf der Evolution einem Problem: Die Argumente beziehen sich in erster Linie auf Vergleiche an rezenten anatomischen Strukturen und an Knochenfossilien. Für die meisten evolutionsbiologisch relevanten Merkmale wäre jedoch eine Weichteilerhaltung der Fossilien notwendig, um den Beweis zu führen. Auch war es bisher nicht möglich, aus der Theorie überprüfbare Vorhersagen abzuleiten, die zu ihrer weiteren Untermauerung hätten beitragen können.
Argumente pro und contra
Unterhautfettgewebe
Der Mensch besitzt als einziger Primat und neben den wasserlebenden Walen, Robben und Seekühen als einziges Säugetier ein leicht ausgeprägtes, in Ansätzen auch wärmeisolierend wirkendes Unterhautfettgewebe (Speckschicht). Dies gab Hardy den ersten Anstoß zu seiner Hypothese. Unterhautfettgewebe isoliert im Wasser den Körper besser als nasse Haare. Andere Wissenschaftler sehen jedoch diese Fettschicht – insbesondere im Zusammenhang mit der Haarlosigkeit und den Schweißdrüsen – als Teil eines komplizierten thermoregulatorischen Systems an. Das Unterhautfettgewebe wäre demnach als Kompensation der mangelhaften Isolierung durch die fehlende Behaarung einzuordnen. Eine weitere Erklärung für dieses Phänomen besagt, dass die Speckschicht der Nahrungsreserve diene und insbesondere den erhöhten Energiebedarf des Gehirns bei Nahrungsmangel decke.
Besonders ausgeprägt ist der Unterschied im Anteil des Fettgewebes am Körpergewebe von menschlichen Säuglingen (vergleichsweise hoher Fettanteil) im Vergleich zu Menschenaffen-Säuglingen, die eher dünn sind. Nach der Wasseraffen-Hypothese schützte dies Säuglinge vor dem Auskühlen im Wasser, wenn sie mit ihren Müttern in das Wasser gingen. Das im Vergleich der Geschlechter eher dürfige Unterhautfettgewebe der Männer wird bei dieser Hypothese allerdings nicht erklärt. Zudem ist die durchschnittliche Dichte von menschlichen Säuglingen deutlich geringer als bei anderen Landsäugern, so dass sie mehr Auftrieb haben. Eine erheblich schlüssigere Erklärung sieht in diesem Merkmal eine Energiereserve des Säuglings.
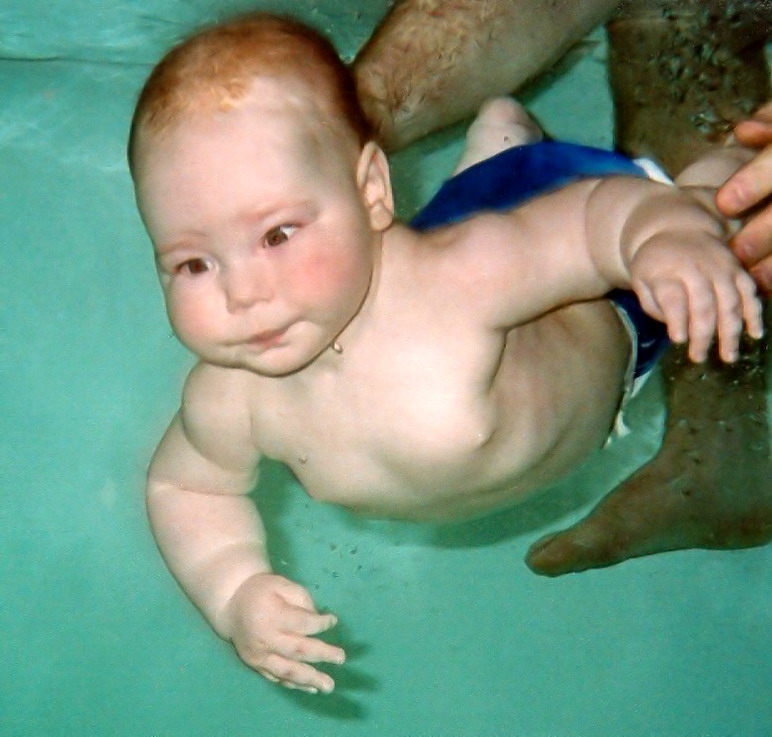
Tauchreflex
 Tauchreflex bei Babys
Tauchreflex bei BabysEin wesentliches Argument für die Hypothese ist das Vorhandensein eines Tauchreflexes beim Menschen. Der Tauchreflex verlangsamt den Herzschlag beim Tauchen und sorgt dafür, dass das Hirn vermehrt mit Blut versorgt wird. Speziell beim Menschen erfolgt der auslösende Reiz durch Benetzung der Gesichtshaut mit Wasser in der Nähe der Nase. Morgan betrachtet dies als eine Anpassung an die aquatische Lebensweise. Dagegen wird eingewendet, dass ein Tauchreflex bei vielen terrestrischen Säugetieren auftauche und wahrscheinlich ein ursprüngliches Merkmal der Säugetiere sei, auch derer, die mit Sicherheit keine aquatische Phase durchgemacht hätten. Immerhin fand dieses Argument im Valkenburg-Symposium insoweit Anerkennung, dass dieses Argument eindeutig für die Wasseraffen-Hypothese spreche und von anderen Hypothesen nicht erfasst werde.
Taucht man einen Säugling unter Wasser, dann wird er kein Wasser verschlucken. Säuglinge sind etwa bis zum zehnten Lebensmonat fähig, den Atem reflexhaft anzuhalten, und erlernen das Schwimmen schnell, dies wird u. a. in Baby-Schwimmkursen genutzt. Morgan sieht in diesem Schutzmechanismus ebenfalls eine Anpassung an das Leben im Wasser.
Schwimmhäute
Bei den meisten Menschen ist vor allem zwischen den Fingern, teilweise auch zwischen den Zehen eine flache Hautpartie ausgebildet, die als Rudiment von Schwimmhäuten interpretiert werden kann. Schimpansen und andere Menschenaffen sollen dieses Merkmal nicht zeigen. Bei einigen Menschen kommt es zu einer fast vollständigen Ausbildung solcher „Schwimmhäute“ (Atavismus). Diese Hautbespannungen deutet Morgan auch als „Schwimmhäute“, die während der aquatischen Phase erworben wurden und möglicherweise wieder teilweise reduziert wurden. Langdon setzt dem entgegen, dass diese „Schwimmhäute“ bei Feten aller Primaten ganz ausgebildet sind und vor der Geburt reduziert werden. Beim Menschen sei die Reduzierung der „Schwimmhäute“ deswegen unvollständig, weil der Mensch an sich neoten sei, das heißt, dass typische Merkmale jugendlicher Stadien bis ins Erwachsenenalter beibehalten werden. Möglicherweise ist dies allerdings lediglich eine Folge der verkürzten Schwangerschaft und der damit einhergehenden Vorverlegung der Geburt, um trotz der Verdreifachung des späteren Gehirnvolumens und der damit einhergehenden Zunahme des Kopfumfanges eine Passage des Geburtskanals zu ermöglichen.
Der aufrechte Gang
 aufrecht stehende Pinguine
aufrecht stehende PinguineDer aufrechte Gang und die Umstrukturierung des Beckens werden als weitere Belege für die Hypothese dargebracht. Wenn sich Menschenaffen gezwungen sehen, in das Wasser zu gehen, dann richten sie sich in der Regel auf und gehen auf den Hinterbeinen. Wenn der Mensch sich über lange Zeit im und am Wasser aufgehalten hat und sich schwimmend fortbewegte, dann musste er sich dem Wasser stromlinienförmig anpassen. Das erklärt, genau wie beim Pinguin, das Kippen des Beckens und die Verlagerung der Beine, unter den Körper. Bei einem Übergang zu einer halb-aquatischen Lebensweise böten dann Mutationen des Beckens und der unteren Extremitäten Selektionsvorteile. Zugleich werden die Arme frei für Werkzeuggebrauch analog zum Verhalten der Seeotter, ein Vergleich, den bereits Hardy anführte. Andere Erklärungen für den aufrechten Gang gehen davon aus, dass Schimpansen bei Waffengebrauch (Stöcke, Wurfsteine) sich des Öfteren aufrichten. Zunehmende kriegerische Gruppenauseinandersetzungen hätten dann zum aufrechten Gang geführt. Wieder andere Erklärungen führen zunehmende Jagd als veränderte Selektionsbedingung an. Nach der Savannen-Hypothese bot der aufrechte Gang mehr Übersicht im Grasland – analog zu Straußenvögeln. Gegner wenden zudem ein, dass es kein wasserlebendes zweibeiniges Säugetier gibt. Zweibeinige Säugetiere (wie Kängurus und zeitweise Primaten) verwenden den aufrechten Gang zur Fortbewegung, zur Nahrungsaufnahme und um die Umgebung zu erkunden. Dagegen wird wiederum eingewendet, dass zahlreiche semiaquatisch lebende Dinosaurier, zum Beispiel Edmontosaurus, sich zweibeinig fortbewegten. Zur Anatomie der Beine wird eingewendet, dass viele Schreitvögel Unterschenkel aufweisen, die länger als die Oberschenkel sind, beim Menschen sei dies nicht der Fall. Allerdings gilt dieses Gegenargument nur für die Knochen; die funktionelle Länge des Oberschenkels ist geringer als die des Unterschenkels, da ein Teil des Oberschenkels im Gesäß verborgen ist.
Der Haarstrich
Die Körperbehaarung des Menschen weist ein stromlinienförmiges Muster auf, welches der Richtung des umgebenden Wassers beim Vorwärtsschwimmen entspricht. Dabei könnte es sich um eine Zwischenphase der Anpassung vor dem Verlust des Felles gehandelt haben. Desmond Morris (Der Mensch mit dem wir leben, Kapitel: Verhalten im Wasser). Dieses Phänomen lässt sich aber auch durch schnelleres Abperlen von Regenwasser bei aufrechtem Gang erklären. Möglicherweise aus dem gleichen Grund besitzen auch andere landlebende Säugetiere (wie z.B. Hunde) ein ganz ähnliches Muster in ihrem Haarstrich.
Nacktheit
Die spärliche Behaarung des Menschen wird ebenfalls als Argument für die Wasseraffen-Hypothese angeführt. Tatsächlich besitzen die am besten an das Leben im Wasser angepassten Säugetiere, die Wale, praktisch keine Behaarung. Weitgehende Nacktheit ist bei Säugern zwar selten, aber durchaus bei verschiedenen Gruppen vorhanden. Neben den Walen sind auch Seekühe und Nilpferde größtenteils unbehaart, auch der in der Erde lebende Nacktmull besitzt praktisch kein Fell. Aber auch weniger an ungewöhnliche Lebensräume angepasste Säuger wie das Erdferkel, der Hirscheber, der Elefant, das Nashorn oder das Warzenschwein sind nur äußerst spärlich behaart. Zudem sind die Menschenaffen zwar noch mit deutlich längerem und flächendeckenderem Haar bedeckt als Menschen, aber auch ihre Haare stehen nur äußerst weit auseinander, und sie besitzen auch kein Unterfell wie die meisten anderen Säuger, im Grunde genommen haben Menschenaffen lediglich eine im Verhältnis zum Menschen noch stark ausgeprägte Körperbehaarung. Außerdem sind auch bei Gorillas, Schimpansen und Orang-Utans viele Körperregionen nur sehr schwach, oder sogar gar nicht behaart, der direkte Unterschied zum spärlich behaarten Menschen ist an sich relativ gering. Verfechter der Wasseraffen-Hypthese ziehen ins Feld, Haarlosigkeit biete im Wasser einen geringeren Strömungswiderstand als Fell. Zudem sei die Funktion des Felles für den Schutz gegen UV-Strahlen bei amphibischer Lebensweise weniger wichtig, außer am Kopf. Eine weitere These erklärt eine einfache Auslese von "stark behaarten" Wasseraffen durch die Erkältungsproblematik bei nassem Fell. Vorwiegend Frauen mit wenig Körperbehaarung hatten bei der ständigen Nahrungssuche in hüfttiefen Gewässern bessere Überlebenschancen (Frauen → weniger Behaarung). Es ist nicht so sehr die effiziente Schwimmanpassung erfolgt, sondern die allgemeine Adaption an Flachwassergebiete (Ausgehend von einer humanoiden Existenzperiode ohne Poleiskappen).[2] Die Gegenargumentation weist darauf hin, dass Robben, Fischotter und Biber als ebenfalls an das Wasser angepasste Lebewesen sehr wohl ein Fell hätten und damit gut schwämmen. Auch die frühen Wale und Seekühe werden zweifelsfrei noch ein Fell gehabt haben. Es wird auch nicht darauf eingegangen, weshalb der Mensch ausgerechnet am Kopf derart ungewöhnlich langes Haar entwickelte, denn dieses wäre für einen tauchenden Hominiden äußerst störend und Sicht-behindernd gewesen. Zudem sei die Anzahl der Haare bei Menschenaffen geringer als bei Menschen. Dagegen wird wiederum eingewandt, dass insbesondere in der Savanne Nacktheit einen erheblichen Selektionsnachteil bedeutet hätte und dass es keine Säugetiere in der Savanne gibt, die ohne Fell auskommen, ohne jedoch zu bedenken, dass etwa Elefant, Spitzmaul-und Breitmaulnashorn, Warzenschwein und Erdferkel, alles Tiere mit extrem spärlicher Behaarung, die gleichen Lebensräume bewohnen wie die frühen Hominiden, wobei für Elefanten und Nashörner aufgrund ihrer Größe und des daraus folgenden Wärmeüberschusses ein Fell mit einem Selektionsnachteil verbunden wäre. Auch die teilweise heute noch unter sehr einfachen Bedingungen lebenden afrikanischen Buschmänner kommen problemlos ohne Fell oder Kleidung in der afrikanischen Savanne aus. Bereits Charles Darwin (Descent of Man, Kapitel 20) bemerkte: „The loss of hair is an inconvenience and probably an injury to man, even in a hot climate, for he is thus exposed to the scorching of the sun, and to sudden chills, especially during wet weather (…) No one supposes that the nakedness of the skin is any direct advantage to man; his body therefore cannot have been divested of hair through natural selection“. Darwin erklärte allerdings die Nacktheit als sekundäres Geschlechtsmerkmal, da in allen Teilen der Welt Frauen weniger behaart seien als Männer.
Geruchssinn
Ein weiteres Argument ist der reduzierte Geruchssinn des Menschen. Morgan zufolge sei der Geruchssinn verkümmert, da er im Wasser kaum eine Bedeutung habe. Zwar ist die Wahrnehmung von Gerüchen im Wasser sehr gut möglich, jedoch nur mit entsprechend angepassten Riechorganen, wie bei Haien und anderen Fischen. Der Geruchssinn der Primaten ist primär jedoch an der Übertragung von Duftstoffen über die Luft angepasst. Morgan schließt daraus, dass der Geruchssinn sich zurückgebildet habe, weil er im Wasser nicht mehr nützlich war. Als Gegenargument wird genannt, dass der hypothetisch aquatische Affe nahezu die ganze Zeit im Wasser gelebt haben müsse, damit eine Reduzierung des Geruchssinnes keinen Selektionsnachteil darstelle. Aufgrund der sehr unvollständigen Anpassung des aquatischen Affen an das Wasser ist dies jedoch unwahrscheinlich. Morgan berücksichtige zudem nicht, dass sämtliche Trockennasenaffen, eine Unterordnung der Primaten, zu der auch der Mensch zählt, einen schlecht entwickelten Geruchssinn besitzen, während stark ans Wasser gebundene Säuger wie Otter, Biber, Wasserspitzmäuse oder selbst Robben einen recht guten Geruchssinn besitzen.
Form der Nase
Die menschliche Nase weist nach unten, was für einen Jäger (nach vorne gerichtete Augen) eher ein Nachteil sein müsste, da so beim Laufen die Atemluft nicht von vorne einströmen kann. Die meisten Affenarten, besonders die Menschenaffen, haben nach vorne geöffnete Nasenlöcher. Die Erklärung für die Nasenform des Menschen beruht ebenfalls auf der Wasseraffen-Theorie, da eine nach unten offene Nase beim Tauchen einen Luftpfropfen wie bei einer Tauchglocke bildet. Dieser Pfropfen ermöglicht ein freihändiges Tauchen, was einem Affen mit nach vorne geöffneter Nase so nicht möglich ist. Arme und Hände können für Schwimmbewegungen genutzt werden und müssen nicht die Nase verschließen. Auffallend ist auch die Möglichkeit, die Nasenflügel bewegen zu können. Manche Menschen sind in der Lage, die Nase fast ganz zu schließen und damit das Eindringen von Wasser zu verhindern. Über diese Fähigkeit verfügen auch andere Meeressäuger. Es ist nicht auszuschließen, dass sich diese Fähigkeit inzwischen sogar etwas zurückentwickelt hat. Dagegen kann argumentiert werden, dass diese Fähigkeit auch für Tiere wie Kamele, die in einem trockenen, staubigen Lebensraum zuhause sind, von Nutzen ist.
Schwimmen und Tauchen
Von einigen Vertretern wird auch auf die für Primaten außergewöhnlichen Leistungen im Schwimmen und Tauchen hingewiesen. So sind trainierte Menschen in der Lage, mehrere Kilometer weit zu schwimmen und ohne Atemgerät über 80 Meter tief zu tauchen. Auch die Tauchzeit ist mit rund einer Minute, bei trainierten Personen sogar mehrere Minuten (siehe Apnoetauchen) vergleichsweise hoch. Zudem sind Menschen mit bis zu 6 km/h Geschwindigkeit im Wasser weitaus schneller als andere primär landlebende Säuger. Die dem zugrunde liegenden anatomischen Besonderheiten des Bewegungsapparates (lange, vergleichsweise strömungsgünstige Körperform, hinten liegende Extremitäten mit höherem Wirkungsgrad, Nutzung der Arme für gerichteten Vortrieb) können allerdings auch als Präadaptation des heutigen Menschen gedeutet werden, die aus einem in anderem Kontext erworbenen aufrechten Gang resultiert. Auch die Nase liegt so, dass durch das Schwimmen weniger Wasser eindringen kann.
Tränenflüssigkeit
Als Beleg für eine marine Episode unserer Vorfahren wird jedoch die Salinität der menschlichen Tränenflüssigkeit gewertet. Mit einem durchschnittlichen Salzgehalt von 3,5 % liegt diese im Bereich des Meerwassers. Diese Konzentration ist typisch für Meeres-Säuger. Terrestrische Säugetiere haben dagegen i. d. R. deutlich weniger Salze in ihrer Tränenflüssigkeit (Ausnahme: der Elefant, für den allerdings eine Abstammung von marinen Vorfahren vorgeschlagen wird).
Als Hinweis auf marine Vorfahren wird auch der – im Vergleich zu anderen Säugern – „verschwenderisch“ hohe Salzgehalt von menschlichem Schweiß interpretiert. Er könnte zum Beispiel zur Ausscheidung überflüssiger, durch die Nahrung aufgenommener Salze gedient haben.
Ernährung und Gehirnentwicklung
Der vergleichsweise hohe menschliche Bedarf und Gehalt von Omega-3-Fettsäuren im Gewebe des Menschen ist ein Indiz für eine auf maritime Nahrungsquellen angepasste Ernährungsumstellung. Diese essentiellen Fettsäuren kommen in terrestrischen Nährstoffen nur in geringen Mengen vor – im Gegensatz zu Nahrungsketten, die auf marinen Algen aufbauen. Die langkettigen Fettsäuren und das auch in Meeresfrüchten reichlich vorhandene Spurenelement Iod sind für die fetale Gehirnentwicklung von entscheidender Bedeutung. Die ausreichende Versorgung mit diesen Stoffen kann in der Vergangenheit Voraussetzung für die sich von den anderen Primaten unterscheidende Gehirnentwicklung gewesen sein.
Strandbewohner
Ein – hypothetischer – Wasseraffe könnte also ein Strandbewohner gewesen sein, der sich seine Nahrung aus dem Meer beschaffte. Der Seeotter nutzt als Nahrungsquelle vor allem marine Krebse und Muscheln. Zum Aufbrechen dieser Schalentiere verwenden die Seeotter Steine als Werkzeuge. Die Wasseraffen-Hypothese benennt für die Entstehung des Menschen also ein Biotop, in dem der Gebrauch von Werkzeugen auch für eine andere Spezies einen unmittelbaren Selektionsvorteil darstellt. Allerdings wird nicht angenommen, dass die Anpassung an das aquatische Biotop bei den Prä-Hominiden so weit wie beim Seeotter ging. Vielmehr war nach der Wasseraffen-Hypothese die partielle Anpassung an den aquatischen Lebensraum zwar eine wesentliche Voraussetzung für die Eroberung terrestrischer Biotope. Sie blieb aber lediglich eine Episode in der Evolution des Homo sapiens.
Psychologische Aspekte
Das Geräusch des Meeresrauschens (wie auch vieler anderer Wassergeräusche) wird nicht als Lärm empfunden, sondern wirkt beruhigend auf den Menschen. Der Mensch hat eine Affinität zum Wasser. Dafür sprechen die hohen Preise für Grundstücke am Wasser und auch die sich großer Beliebtheit erfreuenden Urlaubsreiseziele an Meeren, Seen und Flüssen.
Fossilien-Lücke
Ein in der Frühzeit der Wasser-Affen-Hypothese genanntes Argument besagte, dass man so die frühere Fossilien-Lücke (Mitte des 20. Jahrhunderts waren nur Hominiden-Fossilien datiert, die jünger als ca. 1 Million Jahre waren) erklären könne. Am und im Wasser lebende Fossilien hätten nicht so erhalten bleiben können wie die damals bekannten Funde aus Höhlen. Inzwischen sind aus dem Zeitbereich von vor 5–1 Millionen Jahren zahlreiche Fossilien bekannt, so dass diese Argumentation keine Grundlage mehr hat. Andererseits stammen diese jüngeren Funde fast ausnahmslos aus Sedimenten, die im Wasser entstanden sind, so dass gewissermaßen eine implizite Vorhersage der Hypothese bestätigt wurde.
Falls sich ein Teil der Menschwerdung an der afrikanischen Ostküste abgespielt hat, wären diese Gegenden auch heute für Grabungen unzugänglich, da der Meeresspiegel durch die derzeitige Klimaperiode z. T. mehrere hundert Meter über den Pegeln länger zurückliegender Kaltzeiten liegt.
Kuliukas Wat-Affen-Modell: Eine Bestätigung der Wasseraffen-Hypothese?
 Ein Gorillaweibchen bei der Durchquerung eines Gewässers
Ein Gorillaweibchen bei der Durchquerung eines GewässersDer Londoner Anthropologe Algis Kuliukas unterzog in seiner Master-Arbeit 2001 die Wasseraffen-Hypothese einer Untersuchung vor dem Hintergrund aktueller Forschungsergebnisse zu Hominiden-Fossilien sowie der Verhaltensforschung. Dabei modifizierte und präzisierte er das Modell zu einem „Wat-Affen-Modell“ (wading ape) zur Erklärung der zweibeinigen Fortbewegung des Menschen. Zugleich versuchte er Kriterien der Überprüfbarkeit anzuführen: Falls das Wat-Affen-Modell zutrifft, so müssten rezente Menschenaffen ebenfalls waten, wenn sie angeregt werden, sich in das Wasser zu begeben. Falls sie sich dagegen vierfüßig in das Wasser begeben, so wäre das Modell widerlegt.
Kann dies als Bestätigung der „Wat-Affen-Hypothese“ angesehen werden? Tatsächlich ist bekannt, dass Tiefland-Gorillas, die in sumpfigem Gelände leben, ein zweibeiniges Waten beim Eintritt in das Wasser zeigen. Aus der Literatur ist außerdem bekannt, dass dieses Verhalten auch bei Orang-Utans, Schimpansen und Bonobos auftritt.
Darauf folgende Untersuchungen an Bonobos in einem Wildpark bei Brüssel zeigten, dass diese (dem Menschen genetisch am nächsten verwandten) Menschenaffen an Land lediglich zu 2 Prozent den aufrechten Gang benutzten. Sobald sie teilweise im Wasser waren, stieg die Rate auf 50 Prozent. Dabei benutzten sie ihre Arme fast immer zum Halten der Balance. Bei Bonobos, die vollständig im Wasser waren, wurde zu 92 Prozent aufrechter Gang beobachtet, und zwar fast immer ohne Unterstützung durch die Arme.
Um die richtige historische Perspektive zu bewahren ist es wichtig zu erkennen, dass Kuliukas' Thesen keine Neuerung in der Wasseraffe-Diskussion darstellen. Als Beispiel sei folgende Aussage aus einem Artikel über die Entwicklung der Bipedie des Menschen zu erwähnen, die im Jahr 1997 im Anthropologischer Anzeiger veröffentlicht wurde:
- „Die Vorteile des aufrechten Gangs lassen sich durch die Vielseitigkeit dieser Fortbewegungsart in einer semi-aquatischen Lebensweise verstehen. Die Bipedie der frühen Hominiden kann nicht als eine ideale Anpassung an eine isolierte Fortbewegungsart bezeichnet werden, denn die menschliche Bipedie deckt ein großes Spektrum von lokomotorischen Beanspruchungen ab wie das Tauchen, Schwimmen, Waten, Klettern auf Bäumen und Felsen und terrestrische Fortbewegung“ (Bender, Verhaegen und Oser 1997, 1; s. auch Bender 1999, S.140ff.).
Außerdem ist es fragwürdig, die Falsifikation oder Bestätigung der Wassertheorie von einem einzigen Verhaltensmerkmal (Waten bei Menschenaffen) abhängig zu machen, denn der wichtigste Aspekt dieser Theorie basiert auf der Berücksichtigung verschiedener Merkmale des Menschen, die als Konvergenzen zu dem Merkmalsmosaik anderer Lebewesen gedeutet werden. Und schließlich war es schon vor der Aufstellung Kuliukas' Thesen bekannt, dass Primaten (darunter auch Menschenaffen) im seichten Wasser häufig waten (s. Bibliographie dazu in Bender 1999, S. 109ff.), so dass die in der Wissenschaft übliche Reihenfolge „Hypothese aufstellen - Hypothese überprüfen“ hier nicht eingehalten wurde. Kuliukas verdient trotzdem Anerkennung in der Wassertheorie-Debatte, unter anderem für seinen Versuch, eine einheitliche Definition der Wassertheorie zu liefern.
Neuere Untersuchungen des Paläoklimas von Hominiden-Fundstätten zeigen, dass das Klima dort zur Lebenszeit der Hominiden eher feucht war. Nahezu alle Fundstätten früher Hominiden, die Skelettmerkmale aufrechten Ganges zeigen oder für die aufrechter Gang durch Fußabdrücke nachgewiesen ist, weisen aquatische Sedimente und ehemalige aquatische Biotope in engem örtlichem Zusammenhang auf (Flüsse, Seen). Dies gilt für
- Orrorin tugenensis (wahrscheinlich kein direkter Vorfahr des Menschen, vor 5,6-6,3 Millionen Jahren),
- Ardipithecus ramidus, Äthiopien, vor 4,4-5,8 Millionen Jahren
- Australopithecus anamensis, Turkanasee, vor 3,6 Millionen Jahren
- Australopithecus afarensis, vor 3,2-4,1 Millionen Jahren
- Kenyanthropus platyops, vor 3,3-3,5 Millionen Jahren.
Lediglich ein Fundort (Laetoli) weist keine aquatisch entstandenen Sedimente, sondern vulkanische Tuffe auf. Spätere Funde von Australopithecus africanus (vor 2 Millionen Jahren) liegen dagegen häufig nicht in aquatischem Kontext. Bei der Interpretation dieser Fakten muss allerdings berücksichtigt werden, dass Sedimente von Flüssen und Seen eine deutlich höhere Wahrscheinlichkeit der Erhaltung von Fossilien aufweisen als Sedimente, die in trockenen Gebieten entstanden sind. Die Entdeckung von fossiler Hominiden in mariner Ablagerungen allein sind kein Beweis für die Wasseraffen-Theorie, denn dies könnte im Rahmen der Freilandhypothesen („Savannentheorien“) interpretiert werden. Viel wichtiger als die Sedimente, in welcher die Frühhominiden gefunden werden, ist eine kritische, historische Überprüfung der Argumentationsführung der Freilandhypothesen, ein Thema, das in der paläoanthropologischen Literatur weitgehend vernachlässigt wurde (die Geschichte der Paläoanthropologie befasst sich fast ausschließlich mit der Geschichte der Entdeckung der Fossilien; nur wenige Autoren haben sich überhaupt mit der Geschichte von Theorien in der Paläoanthropologie auseinandergesetzt).
Andere „aquatische“ Primaten
 Japanmakaken in heißer Quelle
Japanmakaken in heißer QuelleIm Zusammenhang mit dem teilweise vehement ausgetragenen Streit um die Wasseraffen-Hypothese stellt sich die Frage, ob die postulierte Anpassung der menschlichen Vorfahren an das Wasser ein einmaliger Vorgang in der Gruppe der Primaten sei.
Tatsächlich gibt es mindestens drei Beispiele für einen unabhängig erfolgten sekundären Übergang von Affen an eine semi-amphibische Lebensweise:
- der Nasenaffe (Nasalis larvatus), der in den Regenwäldern des Tieflands und den Mangrovensümpfen von Borneo lebt. Diese Tiere gehören zu den so genannten Schlankaffen und zeichnen sich durch gutes Schwimm- und Tauchvermögen[1] und eine große Nase aus. Bei Weibchen ähnelt diese der menschlichen Nase, bei Männchen ist sie extrem groß (Sexualdimorphismus).
- der Javaneraffe (Macaca fascicularis), der in Südostasien von Myanmar über Thailand, Indonesien bis zu den Philippinen vorkommt. Javaneraffen gehören zu den Pavianartigen. Sie leben in Wäldern, bevorzugt in der Nähe von Gewässern, auch an der Meeresküste. Diese Tiere schwimmen und tauchen häufig. Zu ihrer Lieblingsnahrung gehören Krabben, die sie am Strand ausgraben und waschen. Sie tauchen mit geöffneten Augen. Das Waschen der Nahrung (auch Früchte werden gewaschen) gilt als nicht ererbte, sondern kulturell weitergegebene Verhaltensweise [2].
- der Japanmakak, (Macaca fuscata) der ebenfalls zu den Pavianartigen gehört. Er lebt auf den japanischen Inseln und badet – insbesondere im Winter – gern in heißen Quellen. Eine Makakenpopulation auf der Insel Koshima ist seit 1948 unter wissenschaftlicher Beobachtung. Besonders bekannt wurde das Weibchen Imo aus dieser Population, das 1953 das Waschen von Nahrung erfand. Diese Verhaltensweise hat sich seitdem durch Tradierung durchgesetzt. Später setzte sich auch das Verhalten durch, Nahrung im Meer zu waschen und damit zu salzen. Seitdem setzte sich bei dieser Makakengruppe das Leben im Meer erstaunlich schnell durch, obwohl Japanmakaken generell das Meer meiden. Mitglieder dieser Population entwickelten seitdem das Schwimmen, Tauchen und die Nutzung von Seetang als Nahrung. Zudem begannen diese Affen, die Hände zum Tragen des Futters zu benutzen und größere Strecken (bis 30 m) zweibeinig zu laufen. Anfang der 80er Jahre des 20. Jahrhunderts begannen die Makaken von Koshima, rohen Fisch zu essen.
Allen drei Beispielen gemeinsam ist, dass diese Arten zwar weiterhin primär Landbewohner bzw. Baumbewohner sind, aber sekundär zusätzlich den Lebensraum Wasser zumindest teilweise erschlossen haben. Grundsätzlich ist also der Bauplan von Primaten auch geeignet, um eine sekundär aquatische Lebensweise zu entwickeln.
Das Beispiel der Japanmakaken von Koshima zeigt, dass bei der Diskussion von Hominisationsmodellen nicht nur die klassischen Evolutionsmechanismen (Mutation / Selektion) zu berücksichtigen sind. Vielmehr können bereits bei Affen durch Entdecken, Lernen und Tradieren neue Verhaltensweise entstehen und – unabhängig von der Vererbung – über Generationen weitergegeben werden (Meme). Solche neuen Verhaltensweisen können – wie die Japanmakaken zeigen – zu veränderten Lebensbedingungen – einschließlich einer Habitatveränderung und einer Änderung der ökologischen Nische führen und damit auf die Evolutionsvorgänge zurückwirken.
Dies kann erstaunlich rasch geschehen, im Fall der Japanmakaken innerhalb von kaum 50 Jahren. Es müssen also nicht zwingend externe ökologische Faktoren (Klimaänderungen, geologische Änderungen etc.) eingewirkt haben, vielmehr könnten neue Nahrungsvorlieben und Verhaltensweisen entstanden sein, die dann die Lebensweise und den Lebensraum der Vorfahren des Menschen änderten. Damit wären sekundär andere Umwelt- und damit Selektionsbedingungen geschaffen, die nun die Durchsetzung genetisch bedingter Anpassungen begünstigen würden.
Literatur
Artikel
- Bender, Renato ; Verhaegen, Marc und Oser, Nicole: Der Erwerb menschlicher Bipedie aus der Sicht der Aquatic Ape Theory in: Anthropologischer Anzeiger 55 (1), 1-14 (1997). (Der erste Artikel eindeutig pro AAT, der in einer wissenschaftlichen anthropologischen Zeitschrift veröffentlicht wurde.)
- Bender, Renato und Oser, Nicole : Gottesanbeterinnen, Maulwürfe und Menschen. Unipress (Berichte über Forschung und Wissenschaft an der Universität Bern, herausg. v. der Pressstelle) 95, 20-26 (1997) (in dieser Arbeit befassen sich die Autoren mit den Argumenten der Freilandhypothesen und der AAT)
- Hardy, Alister: Was man more aquatic in the past in: New Scientist vom 17. März 1960
- Langdon, John H.: Umbrella hypotheses and parsimony in human evolution in: Journal of Human Evolution 33.1997 – S. 479–494 (In diesem Artikel werden Argumente für die Aquatic-Ape-Hypothese zusammengefasst und kritisch aus dem Blickwinkel der Savannah-Hypothese betrachtet)
Monographien
- Bender, Renato: Die evolutionsbiologische Grundlage des menschlichen Schwimmens, Tauchens und Watens: Konvergenzforschung in den Terrestrisierungshypothesen und in der Aquatic Ape Theory. Dimplomarbeit am Institut für Sport und Sportwissenschaft, Universität Bern (1999). (in dieser Arbeit wurden die Freilandhypothesen zum ersten Mal historisch analysiert. Die Argumente sowohl der Freilandhypothesen als auch der verschiedenen Wassertheorien wurden unter der Perspektive der Konvergenzforschung analysiert. Der Autor vertritt eine Haltung pro AAT)
- Bender-Oser, Nicole: Die Aquatile Hypothese zum Ursprung des Menschen: Max Westenhöfers Theorie und ihre Bedeutung für die Anthropologie. - Inaugural-Dissertation zur Erlangung der Doktorwürde der Humanmedizin der Medizinischen Fakultät der Universität Bern (2004). (eine ausführliche Arbeit zur Geschichte der Aquatilen Hypothese; gleichzeitig werden hier die Argumente der Aquat. Hypoth. und der AAT verglichen. Die Arbeit ist pro AAT)
- Niemitz, Carsten: Das Geheimnis des aufrechten Gangs: unsere Evolution verlief anders. München: Beck, 2004. – ISBN 978-3-406-51606-1
- Morgan, Elaine: Der Mythos vom schwachen Geschlecht: wie die Frauen wurden, wie sie sind. München: Goldmann, 1990. ISBN 3-442-11457-8 (Dieses feministische Buch greift die androzentrischen Erklärungsversuche der Menschwerdung an (Savannah-Hypothese); dem stellt sie ihre Menschwerdungshypothese gegenüber (Aquatic-Ape-Hypothese))
- Morgan, Elaine: The Aquatic Ape: A Theory of Human Evolution
- Morgan, Elaine: The Scars of Evolution: What Our Bodies Tell us about Human Evolution
- Morgan, Elaine: The Aquatic Ape Hypothesis: The Most Credible Theory of Human Evolution
- Westenhöfer, Max: Der Eigenweg des Menschen: dargestellt auf Grund von vergleichend morphologischen Untersuchungen über die Artenbildung und Menschwerdung. Berlin: Mannststaedt, 1942
- Westenhöfer, Max : Die Grundlagen meiner Theorie vom Eigenweg des Menschen: Entwicklung, Menschwerdung, Weltanschauung. Heidelberg: Winter, 1948 (in diesem Buch wird die Aquatile Hypothese nicht behandelt)
Weblinks
- Dr. med vet. Corina Gericke: Affenkultur (Beobachtungen an Japanmakaken)
- Text aus Agemus 26/1991 nach Vortrag von Michael Bujatti-Narbeshuber am 2. Symposium für Neurobiologie in Salzburg 1991
Einzelnachweise



Wikimedia Foundation.