- C4-Pflanze
-
C4-Pflanzen nutzen einen Stoffwechselweg, um Kohlenstoffdioxid für die Photosynthese zunächst räumlich vorzufixieren und erst dann wie C3-Pflanzen im Calvin-Zyklus zu Kohlenhydraten aufzubauen. Der Name C4 leitet sich vom ersten Fixierungsprodukt ab, welches durch die Assimilation von Kohlenstoffdioxid entsteht. Während dies bei C3-Pflanzen eine Kohlenstoffverbindung mit drei C-Atomen ist, findet man in C4-Pflanzen eine Verbindung mit vier C-Atomen.
Die Kohlenstoffdioxid-Assimilation und der Calvin-Zyklus erfolgen in C4-Pflanzen räumlich voneinander getrennt. Durch Aufbringung von Energie wird dadurch Kohlenstoffdioxid aktiv angereichert, was zu einer höheren Photosyntheserate – besonders unter Wassermangel – führt. Daher sind C4-Pflanzen den C3-Pflanzen unter ariden Bedingungen überlegen. Durch die aktive Anreicherung findet die Photorespiration deutlich seltener statt. Typische sind insbesondere Gräser, darunter auch bekannte Nutzpflanzen wie Mais, Zuckerrohr und Hirse, aber auch andere Arten, wie Amarant.
Pflanzen mit einem Crassulaceen-Säurestoffwechsel verfahren ähnlich wie C4-Pflanzen, bei ihnen sind Vorfixierung und der Calvin-Zyklus indes zeitlich voneinander getrennt.
Inhaltsverzeichnis
Vorkommen der C4-Photosynthese im Pflanzenreich
 Rispenhirse, eine C4-Pflanze.
Rispenhirse, eine C4-Pflanze.
Etwa 5% der Bedecktsamer betreiben C4-Photosynthese. Bekannte C4-Pflanzen sind Amarant, Hirse, Mais, Zuckerrohr und Chinaschilf. Die meisten gehören zu den Gräsern, gefolgt von Seggen. Doch auch bei einer Reihe von Zweikeimblättrigen gibt es diesen Stoffwechselweg, insbesondere bei den Fuchsschwanzgewächsen und anderen Nelkenartigen, bei Wolfsmilchgewächsen und vereinzelt bei Windengewächsen und Korbblütlern. Da die C4-Photosynthese 50-mal unabhängig voneinander entstanden ist, spricht man von einem polyphyletischen Merkmal.
Der erste Auftreten von C4-Pflanzen ist noch Gegenstand der Forschung. Für die Datierung werden verschiedene Techniken wie DNA-Analysen (phylogenetische Studien), geochemische Signale (z. B. das Isotopenverhältnis von 12C und 13C), Fossile und Mikrofossile (Pollen, Phytolithe) genutzt. Eine starke Ausbreitung von C4-Pflanzen, und damit eine Ausweitung der von C4-Pflanzen dominierenden Ökosystemen, fand zum Ende des Miozäns und Beginn des Pliozäns vor 2–8 Millionen Jahren statt.[1] Als Grund gelten verschiedene Faktoren, wie klimatische Veränderungen, Feuer oder die Koevolution von Weidetieren.
Es gibt immer mehr Hinweise darauf, dass der Ursprung der C4-Photosynthese etwa 25 Millionen Jahren vor unserer Zeit liegt.[2] Sinkende Temperaturen und Kohlenstoffdioxidkonzentrationen kennzeichnen diese Periode. Ob bereits eine C4-Photosynthese sehr viel früher stattgefunden hat, kann durch heutige Untersuchungstechniken nicht eindeutig geschlussfolgert werden. Beispielsweise können Isotopenwerte aus marinen Sedimenten im heutigen Afrika so gedeutet werden, dass erste C4-Pflanzen bereits vor 90 Millionen Jahre existierten. Ein noch früherer möglicher Zeitpunkt wäre der Übergang vom späten Karbon zum frühen Perm vor etwa 300 Millionen Jahren, da das vorherrschende Verhältnis von Sauerstoff zu Kohlenstoffdioxid einen Selektionsdruck zugunsten von C4-Pflanzen ausgelöst hätte.
Entdeckung
Die ersten Untersuchungen an einer C4-Pflanze wurden durch Hugo Kortschak durchgeführt. Um 1950 identifizierte er an einem Zuckerrohr-Forschungsinstitut in Hawaii als erstes CO2-Fixierungsprodukt L-Malat und L-Aspartat. Diese sind C4-Verbindungen und standen damit in Widerspruch zu den Befunden von Melvin Calvin, Andrew Benson und James Bassham. Diese hatten gezeigt, dass das erste Stoffwechselprodukt bei der CO2-Fixierung in der Dunkelreaktion eine C3-Verbindung, 3-Phosphoglycerat, ist. Die Ergebnisse Kortschaks wurden aber erst zehn Jahre später veröffentlicht. Auch der Russe Yuri Karpilov entdeckte an Mais, dass das erste Fixierungsprodukt eine C4-Verbindung ist die auch den Namen des Photosynthesetyps prägte.
Erst die australischen Forscher Marshall Davidson Hatch und Charles Roger Slack konnten mit jenen Ergebnissen und eigenen Untersuchungen die Biochemie des Stoffwechselweges entschlüsseln. Die Funktion und Bedeutung wurde Ende der 1970er-Jahre veröffentlicht.[3] Infolgedessen wird der C4-Stoffwechsel nach seinen Entdeckern auch als Hatch-Slack-Weg bzw. Hatch-Slack-Zyklus bezeichnet.
Anatomie
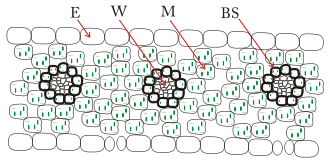 Schematischer Querschnitt eines Blattes einer C4-Pflanze:. E: Epidermis; M: Mesophyllzelle; W: Xylem/Phloem; BS: Bündelscheidenzelle. Auf der Blattunterseite befinden sich die Schließzellen.
Schematischer Querschnitt eines Blattes einer C4-Pflanze:. E: Epidermis; M: Mesophyllzelle; W: Xylem/Phloem; BS: Bündelscheidenzelle. Auf der Blattunterseite befinden sich die Schließzellen.Die Reaktionen des C4-Stoffwechselweges erstrecken sich meistens über zwei benachbarte Zelltypen, welche den C4-Pflanzen auch eine typische Anatomie verleihen. Hierbei wird CO2 zunächst in Mesophyllzellen (A, vgl. Abbildung) in eine C4-Verbindung vorfixiert. Mesophyllzellen enthalten kein RubisCO. Das in Form von jener C4-Verbindung gebundene CO2 wird über Plasmodesmen in Bündelscheidenzellen (B, vgl. Abbildung) transportiert und dort freigesetzt. Diese können aufgrund ihrer Enzymausstattung den Calvin-Zyklus ausführen. Da die Bündelscheidenzellen von den Mesophyllzellen kranzförmig umgeben sind, spricht man auch von einer Kranzanatomie. Die bei der Freisetzung entstehende C3-Verbindung wird wieder in die Mesophyllzelle zurücktransportiert.
Die CO2-Fixierung (in Mesophyllzellen) und die eigentliche Kohlenstoffassimilation im Calvin-Zyklus (in Bündelscheidenzellen) sind räumlich getrennt.
Biochemie
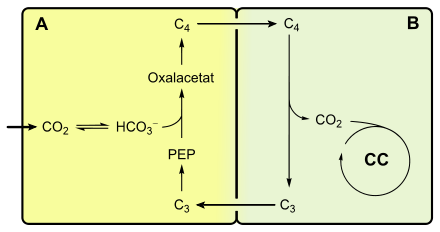 Übersicht über den C4-Stoffwechsel: A: Mesophyllzelle; B: Bündelscheidenzelle; PEP: Phosphoenolpyruvat; O: Oxalacetat; M: Malat; Pyr: Pyruvat; CC: Calvin-Zyklus
Übersicht über den C4-Stoffwechsel: A: Mesophyllzelle; B: Bündelscheidenzelle; PEP: Phosphoenolpyruvat; O: Oxalacetat; M: Malat; Pyr: Pyruvat; CC: Calvin-ZyklusDieser Vorgang stellt eine CO2-Pumpe dar, die mit der Fixierung von CO2 in Form von Bicarbonat (HCO3−) beginnt. Die Bildung von HCO3− aus CO2 wird durch eine Carboanhydrase katalysiert.
Reaktionen in der Mesophyllzelle
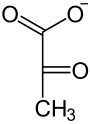
Eine Phosphoenolpyruvat-Carboxylase katalysiert die Kondensation eines Moleküls Phosphoenolpyruvat mit HCO3−, so dass Oxalacetat entsteht. Dieses wird in L-Malat ("Malatbildner"), bei manchen C4-Pflanzen auch in L-Aspartat, umgesetzt.
ATP, AMP,
Pi PPi

PPDK 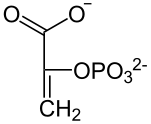
HCO3− Pi
PEP-C 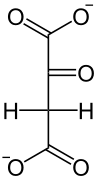
Pyruvat Phosphoenolpyruvat Oxalacetat NADPH
+H+ NADP+

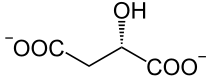
MDH L-Malat Glu KGS
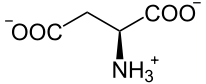
ASAT Aspartat In dieser Form wird es durch die Plasmodesmen in die Bündelscheidenzellen transportiert.
Reaktionen in der Bündelscheidezelle
Wie nun das Kohlenstoffdioxid wieder freigesetzt und in den Calvin-Zyklus eingespeist wird, ist bei C4-Pflanzen unterschiedlich.
- Ein NADP-abhängiges Malatenzym katalysiert die Decarboxylierung von Malat zu Pyruvat. Hierbei wird entweder
- NADP+ bei einem NADP-Malat-Enzymtyp in den Chloroplasten reduziert. Oder
- dies erfolgt mit NAD+ bei einem NAD-abhängigen Malatenzym in den Mitochondrien.
- Bei dem Phosphoenolpyruvat-Carboxykinasetyp wird im Cytosol Oxalacetat zu Phosphoenolpyruvat unter ATP-Verbrauch decarboxyliert.
Die Zellwände der Bündelscheidenzellen sind meist suberinisiert, wodurch das frei werdende CO2 nicht aus der Zelle diffundieren kann. Es sammelt sich in der Zelle an, so dass eine hohe CO2-Konzentration für die RubisCO herrscht, die es zur Assimilation in den Calvin-Zyklus einführt.
Bei Pflanzen mit dem NADP-abhängigen Malatenzym kann das entstehende NADPH, im reduktiven Teil des Calvin-Zyklus genutzt werden. So finden sich in deren Bündelscheidenzellen oft relativ große Chloroplasten ohne Granastapel. Dementsprechend fehlt ihnen das Photosystem II (PS II), welches normalerweise in den Granamembranen der Thylakoide lokalisiert ist. Dadurch fehlt die Möglichkeit zum linearen Elektronentransport der Lichtreaktion der Photosynthese, der normalerweise das im Calvin-Zyklus benötigte NADPH erzeugt.
Nach der Freisetzung des CO2 werden Phosphoenolpyruvat (PEP) bzw. Pyruvat durch Plasmodesmen wieder zurück in die Mesophyllzelle transportiert. Bei Pflanzen mit dem NAD-Malat-Enzymtyp wird Pyruvat vorher durch Transaminierung zu L-Alanin umgewandelt und dieses dann transportiert. Pyruvat selbst wird durch eine Pyruvat-Phosphat-Dikinase in den Chloroplasten der Mesophyllzelle unter ATP-Verbrauch zu PEP umgesetzt und kann damit wieder mit Bicarbonat kondensieren.
Ökonomische und ökologische Aspekte
- C4-Pflanzen können zur Produktion von Biomasse für die Energiegewinnung genutzt werden. Chinaschilf erreicht Erträge von 15 bis 25 Tonnen Trockenmasse je Hektar und Jahr.[4]
- Obwohl zu den Gräsern gehörend, gehört Reis zu den C3-Pflanzen. Versuche, durch Einbringen verschiedener Gene aus C4-Pflanzen die Photosyntheserate zu erhöhen, waren bis jetzt wenig erfolgversprechend.[5]
In den letzten dreißig Jahren ist eine Ausbreitung von C4-Pflanzen auch auf warmen, sonnigen Standorten in Mitteleuropa zu beobachten. Zumeist handelt es sich um hirseartige Gräser und Fuchsschwanzarten. Deren Ausbreitung wird zumindest bisher nicht als Gefahr für die heimische Flora gewertet.[6]
Wasserbedarf
C4-Pflanzen sind den meisten C3-Pflanzen insofern überlegen, als dass sie durch ihre Kohlenstoffdioxidanreichung Wasser ökonomischer nutzen können (WUE, water use efficiency): Die optimale Wachstumstemperatur liegt zwischen 30 und 40 °C, für C3-Pflanzen dagegen bei 20–30 °C.[7]
Bei steigender Temperatur löst sich Sauerstoff besser im Vergleich zu CO2, so dass es bei C3-Pflanzen zu größeren Verlusten durch Photorespiration aufgrund der Oxygenaseaktivität der RubisCO kommt, die bei C4-Pflanzen reduziert bis vollständig unterdrückt werden kann.
Während C4-Pflanzen zur Bildung von 1 g Trockenmasse 230–250 ml Wasser benötigen, liegt der Bedarf für C3-Pflanzen zwei bis dreimal so hoch.
Stickstoffbedarf
Der Stickstoffbedarf für C4-Pflanzen ist niedriger, da sie weniger RubisCO benötigen. Diese kann nämlich aufgrund der höheren CO2-Sättigung effizienter arbeiten, Verluste durch Photorespiration sind minimal. Man hat berechnet, dass bei 30 °C ein C4-Blatt etwa 13-20 % der Menge an RubisCO eines C3-Blattes benötigt, um dieselbe Photosyntheserate (bei gesättigter Lichtstärke) zu erreichen.[8] Es muss dabei aber angemerkt werden, dass typische C4-Enzyme – wie die PPDK und PEPC – einen erhöhten Stickstoffbedarf nach sich ziehen.
Insgesamt schätzt man, dass die sogenannte Stickstoffnutzungseffizienz (NUE, nitrogen use efficiency) in C4-Pflanzen mindestens doppelt so hoch ist wie in C3-Pflanzen.
Wachsende Aufmerksamkeit gewinnen auch tropische C4-Futtergräser, die mit stickstofffixierenden Bakterien vergesellschaftet sind und somit kaum einer Zusatzdüngung bedürfen.
Energiebedarf
Der Energiebedarf einer C4-Pflanze (NADP-ME-Typ und NAD-ME-Typ) beträgt 5 ATP und 2 NADPH pro fixiertem CO2-Molekül und liegt somit höher als der einer C3-Pflanze. C3-Pflanzen benötigen 3 ATP und 2 NADPH pro fixiertem CO2-Molekül, wobei diese Werte die photorespiratorischen Verluste außer Acht lassen.
C4-Pflanzen des PEPCK-Typs benötigen 3,6 ATP und 2,3 NADPH pro fixiertem CO2-Molekül.[9]
Isotopendiskriminierung
C4-Pflanzen lassen sich durch das Verhältnis der beiden Kohlenstoff-Isotope 12C und 13C erkennen. Die beiden Isotope kommen in der Atmosphäre mit 98,89 % und 1,11 % vor (das radioaktive Isotop 14C spielt in diesem Zusammenhang keine Rolle). Das Enzym RubisCO reagiert mit 12CO2 und diskriminiert gegen 13CO2, bei C4-Pflanzen ist daher das 13C/12C Verhältnis höher als in C3-Pflanzen. Es wird als δ-13C-Wert ausgedrückt:
Als Standard ist ein bestimmtes Kalkgestein definiert (Pee Dee Belemnite). Produkte der C3-Photosynthese besitzen δ-13C-Werte von rund −28 ‰.
Die PEP-Carboxylase präferiert 12CO2 weniger stark als RubisCO, in C4-Pflanzen wird jedoch fast das gesamte CO2 durch die PEP-Carboxylase vorfixiert. Durch die hohe interne CO2-Konzentration in den Bündelscheidenzellen kommt auch die Diskriminierung der RubisCO nicht zum Tragen. Daraus ergibt sich für C4-Pflanzen ein δ-13C-Wert von durchschnittlich −14 ‰. Durch Bestimmung des δ-13C-Wertes mittels Massenspektrometrie kann man daher unterscheiden, ob Zucker aus der Zuckerrübe (C3) oder aus Zuckerrohr (C4) stammt.
Literatur
- Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3
- Ghannoum. O. (2009): C4 photosynthesis and water stress. In: Ann Bot. 103(4); 635–644; PMID 18552367; PDF (freier Volltextzugriff, engl.)
- Hibberd, JM., Sheehy, JE. und Langdale, JA. (2008): Using C4 photosynthesis to increase the yield of rice-rationale and feasibility. In: Curr Opin Plant Biol. 11(2); 228–331; PMID 18203653; doi:10.1016/j.pbi.2007.11.002
- Osborne, CP. und Beerling, DJ. (2006): Nature's green revolution: the remarkable evolutionary rise of C4 plants. In: Philos Trans R Soc Lond B Biol Sci. 361(1465); 173–194; PMID 16553316; PDF (freier Volltextzugriff, engl.)
- Hans W. Heldt und Birgit Piechulla: Pflanzenbiochemie. 4 Auflage. Spektrum Akademischer Verlag, 2008, ISBN 978-3-8274-1961-3, S. 211ff.
- Elmar Weiler, Lutz Nover: Allgemeine und molekulare Botanik. Thieme, Stuttgart 2008, ISBN 978-3-13-147661-6.
- Eduard Strasburger (Begr.), Peter Sitte, Elmar Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner: Lehrbuch der Botanik für Hochschulen. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X.
Einzelnachweise
- ↑ Colin P. Osborne: The Geologic History of C4 plants. In: Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3; S. 339–357
- ↑ Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3, S. xix
- ↑ Slack, CR. und Hatch, MD. (1967): Comparative studies on the activity of carboxylases and other enzymes in relation to the new pathway of photosynthetic carbon dioxide fixation in tropical grasses. In: Biochem J. 103(3); 660–665; PMID 4292834; PDF (freier Volltextzugriff, engl.)
- ↑ Produktionsaspekte Weiden und Miscanthus
- ↑ James N. Burnell: Hurdles to Engineering greater photosynthetic rates in crop plants: C4 rice. In: Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3; S. 361–378
- ↑ Brandes, Dietmar (1995) Breiten sich C4-Pflanzen in Mitteleuropa aus?
- ↑ Rowan F. Sage, Ferit Kocacinar und David S. Kubien: C4 Photosynthesis and temperature. In: Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3; S. 161–195
- ↑ Michael B. Jones: C4 species as energy crops. In: Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3; S. 379–397
- ↑ Gerald E. Edwards und Elena V. Voznesenskaya: C4 photosynthesis: Kranz Forms and Single-Cell C4 in terrestrial plants. In: Agepati S. Raghavendra (Hrsg.) und Rowan F. Sage (Hrsg.): C4 Photosynthesis and Related CO2 Concentrated Mechanisms (Advances in Photosynthesis and Respiration). Springer Netherlands 2011; ISBN 978-90-481-9406-3; S. 29–61
Weblinks
Kategorien:- Biologischer Prozess
- Pflanzentyp
- Photosynthese










Wikimedia Foundation.