- Fuchsschwanzgewächse
-
Fuchsschwanzgewächse 
Zurückgebogener Amarant (Amaranthus retroflexus)
Systematik Unterabteilung: Samenpflanzen (Spermatophytina) Klasse: Bedecktsamer (Magnoliopsida) Eudikotyledonen Kerneudikotyledonen Ordnung: Nelkenartige (Caryophyllales) Familie: Fuchsschwanzgewächse Wissenschaftlicher Name Amaranthaceae Juss. Die Fuchsschwanzgewächse (Amaranthaceae) sind eine Familie in der Ordnung der Nelkenartigen (Caryophyllales) innerhalb der Bedecktsamigen Pflanzen (Magnoliopsida). Hier ist auch die oft separat geführte Familie der Gänsefußgewächse (Chenopodiaceae) eingegliedert.
Inhaltsverzeichnis
Beschreibung
 Illustration der Sand-Radmelde (Bassia laniflora), Camphorosmoideae.
Illustration der Sand-Radmelde (Bassia laniflora), Camphorosmoideae.
Erscheinungsbild und Blätter
Die meisten Arten sind einjährige oder ausdauernde krautige Pflanzen oder Halbsträucher; es gibt auch einige Sträucher; wenige Arten sind Lianen oder Bäume. Viele Arten sind sukkulent. Bei vielen Arten sind an den Sprossachsen die Knoten (Nodien) verdickt. Das Holz der mehrjährigen Sprossachsenteile weist ein für die Familie typisches „anomales“ Dickenwachstum auf, welches nur den Polycnemoideae fehlt.
Die Laubblätter sind meist wechselständig, gelegentlich auch gegenständig. Nebenblätter sind keine vorhanden. Die Gestalt der Blätter ist äußerst variabel, die Blattränder sind ganzrandig oder gezähnt, ihr Querschnitt ist flächig bis stielrund, bei einigen Arten sind die Blätter auch zu winzigen Schuppen reduziert.
 Fünfzählige Blüten von Nitrophila occidentalis, Polycnemoideae.
Fünfzählige Blüten von Nitrophila occidentalis, Polycnemoideae. Pantoporate Pollenkörner von Halothamnus glaucus, Raster-Elektronen-Mikroskopie
Pantoporate Pollenkörner von Halothamnus glaucus, Raster-Elektronen-MikroskopieBlüten
Die Vorblätter und Tragblätter sind entweder krautig oder trockenhäutig. Die Blütenhülle ist mehr oder weniger krautig oder auch trockenhäutig und besteht aus (selten eines bis) meist fünf (selten bis acht) Tepalen. Die meist in gleicher Anzahl vorhandenen Staubblätter stehen entweder vor den Tepalen oder dazwischen. Sie entspringen am Blütengrund einem Diskus, welcher bei einigen Arten Anhängsel (Pseudostaminodien) trägt. Die Staubbeutel besitzen zwei oder vier Pollensäcke. Bei den Caroxyloneae kommen blasenförmige Staubbeutelanhängsel vor. Die Pollenkörner sind rund mit zahlreichen Öffnungen (pantoporat), wobei die Porenzahl von wenigen bis zu 250 (bei Froelichia) reichen kann.[1] Die meist ein bis drei (selten bis sechs) Fruchtblätter sind zu einem oberständigen Fruchtknoten verwachsen. Im Fruchtknoten befindet sich eine (selten zwei) basale Samenanlage.
Früchte und Samen
Als Ausbreitungseinheit (Diasporen) dienen entweder die Samen, oft sind aber die Blütenhüllblätter auch noch bei der reifen Frucht vorhanden und erfahren Umgestaltungen, die der Ausbreitung dienen. Gelegentlich werden auch die Trag- und Vorblätter in die Diaspore einbezogen. Seltener kommen Kapselfrüchte oder Beeren vor. Der Same ist horizontal oder anders ausgerichtet, oft mit verdickter und verholzter Samenschale. Der Embryo ist grün oder weiß, spiralig (dann ohne Nährgewebe) oder ringförmig (selten gerade).
Chromosomenzahl
Die Chromosomengrundzahl ist (selten sechs) meist acht oder neun (selten 17).
Inhaltsstoffe
Weitverbreitet bei den Fuchsschwanzgewächsen ist das Vorkommen von Betalainen. Die früheren Chenopodiaceae enthalten oft Isoflavonoide.
Bei phytochemischen Untersuchungen wurden zudem Methylendioxyflavonole, Saponine, Triterpene, Ecdysteroide sowie in den Wurzeln spezielle Kohlenhydrate gefunden[1].
Photosyntheseweg
Mit etwa 800 C4-Arten sind die Fuchsschwanzgewächse die größte Gruppe mit diesem Photosyntheseweg. Innerhalb der Familie gibt es verschiedene Typen von C4-Photosynthese und etwa 17 verschiedene Typen der Blattanatomie. Daher wird angenommen, dass diese Eigenschaft wahrscheinlich etwa 15-mal unabhängig voneinander in der Evolution dieser Familie erworben wurde, davon 2/3 bei den ehemaligen Chenopodiaceae. Das erste Mal trat C4-Photosynthese im frühen Miozän vor etwa 24 Millionen Jahren auf. Bei einigen Gruppen entstand dieser Photosyntheseweg aber erst wesentlich später, vor sechs (oder weniger) Millionen Jahren.
Die mehrfache Entwicklung von C4-Photosynthese bei den Amaranthaceae wird als Anpassung an zunehmende Wasserknappheit und höhere Temperaturen angesehen. Diese Arten waren durch ihre größere Effizienz im Wasserverbrauch in trockenen Lebensräumen im Vorteil und konnten sich dort ausbreiten.[2]
Nutzung
Einige Arten, wie etwa der Spinat (Spinacia oleracea) oder Kulturformen der Rübe (Beta vulgaris) (z. B. Rote Rübe, Mangold), werden als Gemüsepflanzen genutzt. Weitere Formen von Beta vulgaris sind die Futterrübe und die Zuckerrübe. Die Samen mehrerer Amaranthus-Arten und der Quinoa (Chenopodium quinoa), selten auch Reismelde genannt, werden ähnlich wie Getreide (Pseudogetreide) verwendet. Aus mehreren Arten wird Soda gewonnen.
Als Zierpflanzen werden beispielsweise Garten-Fuchsschwanz, Brandschopf und Iresine verwendet.
Verbreitung
Die Familie Amaranthaceae ist weltweit in den gemäßigten Zonen sowie den Subtropen und Tropen verbreitet. Viele Arten sind an Böden mit relativ hohem Salzgehalt angepasst oder kommen in trockenen Steppen- und Halbwüstengebieten vor.
Systematik
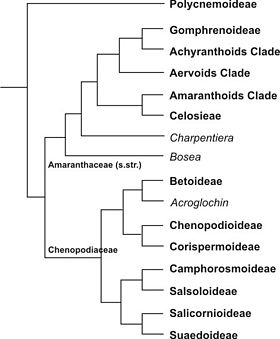 Kladogramm der Pflanzenfamilie Amaranthaceae s. l., verändert und vereinfacht, nach phylogenetischen Arbeiten von Müller & Borsch 2005, Kadereit & al. 2006, Sanchez del-Pino & al. 2009.
Kladogramm der Pflanzenfamilie Amaranthaceae s. l., verändert und vereinfacht, nach phylogenetischen Arbeiten von Müller & Borsch 2005, Kadereit & al. 2006, Sanchez del-Pino & al. 2009.Eine Gruppe von Gattungen, die mehr als die Hälfte der Arten umfasst, wurde traditionell als eigene Familie Gänsefußgewächse (Chenopodiaceae) abgetrennt. Molekularbiologische Untersuchungen haben aber eine enge Verwandtschaft der beiden traditionell unterschiedenen Familien ergeben und nahegelegt, dass die Gänsefußgewächse paraphyletisch sind und nur bei einer Vereinigung mit den Amaranthaceae im engeren Sinn ein monophyletisches Taxon ergeben. Aktuelle Veröffentlichungen verwenden weiterhin den Familiennamen Chenopodiaceae [3][4][5][6][7][8].
Die Familie Amaranthaceae wurde 1789 durch Antoine Laurent de Jussieu in Genera Plantarum, S. 87–88 aufgestellt. Die Erstveröffentlichung der Familie Chenopodiaceae erfolgte 1799 durch Étienne Pierre Ventenat in Tableau du Regne Vegetal, 2, S. 253. Der älteste und nach der Prioritätsregel gültige wissenschaftliche Name des erweiterten Taxons ist Amaranthaceae.
Aus dem Kladogramm wird deutlich, dass die Einstufung der Chenopodiaceae entscheidend von der Unterfamilie Polycnemoideae abhängt: Wird sie so wie bisher als Teil der Familie betrachtet, müssen auch die Amaranthaceae im engeren Sinn dazugehören, und der Name der erweiterten Familie lautet nach der Prioritätsregel Amaranthaceae. Würden die Polycnemoideae jedoch als eigene Familie abgetrennt, dann wären Chenopodiaceae und Amaranthaceae zwei jeweils monophyletische Verwandtschaftsgruppen, die als separate Familien betrachtet werden könnten.
In der Familie Amaranthaceae im neuen Umfang sind die Gattungen der früheren Familien: Achyranthaceae Raf., Atriplicaceae Durande, Betaceae Burnett, Blitaceae T.Post & Kuntze, Celosiaceae Martynov, Chenopodiaceae Ventenat, nom. cons., Corispermaceae Link, Deeringiaceae J.Agardh, Dysphaniaceae (Pax) Pax, nom. cons., Gomphrenaceae Raf., Polycnemaceae Menge, Salicorniaceae Martynov, Salsolaceae Menge, Spinaciaceae Menge enthalten.
Die Systematik der Familie Amaranthaceae wird seit einigen Jahren intensiv erforscht. Molekularbiologische Studien haben gezeigt, dass die bisherige Einteilung, die auf morphologischen und anatomischen Merkmalen beruhte, nicht die verwandtschaftlichen Beziehungen widerspiegelte. Die früher den Gänsefußgewächsen (Chenopodiaceae) zugerechneten Arten werden derzeit (2011) in etwa acht Unterfamilien eingeordnet (die Bearbeitung ist noch nicht abgeschlossen): die Polycnemoideae[1], welche als ursprünglich angesehen werden, sowie die Betoideae[3], Camphorosmoideae[4], Chenopodioideae[5], Corispermoideae[9], Salicornioideae[6], Salsoloideae[7] und Suaedoideae[10]. Innerhalb der Amaranthaceae s. str. (im engeren Sinne) erwiesen sich die Unterfamilie Amaranthoideae sowie manche Gattungen innerhalb der Gomphrenoideae als polyphyletisch, so dass hier in den nächsten Jahren weitere taxonomische Änderungen zu erwarten sind [11]
Die Familie Amaranthaceae s. l. wird in der vorläufigen Systematik in zehn Unterfamilien gegliedert und enthält etwa 176 Gattungen mit etwa 2050 bis 2500 Arten:
 Achyranthes splendens, Amaranthoideae.
Achyranthes splendens, Amaranthoideae. Illustration von Deeringia amaranthoides, Amaranthoideae.
Illustration von Deeringia amaranthoides, Amaranthoideae. Gomphrena arborescens, Gomphrenoideae.
Gomphrena arborescens, Gomphrenoideae. Zuckerrübe (Beta vulgaris subsp. vulgaris var. altissima), Betoideae.
Zuckerrübe (Beta vulgaris subsp. vulgaris var. altissima), Betoideae. Maireana sedifolia, Camphorosmoideae.
Maireana sedifolia, Camphorosmoideae. Grayia spinosa, Chenopodioideae.
Grayia spinosa, Chenopodioideae. Arthrocnemum perenne, Salicornioideae.
Arthrocnemum perenne, Salicornioideae. Kali-Salzkraut (Kali soda), Salsoloideae.
Kali-Salzkraut (Kali soda), Salsoloideae. Suaeda nigra, Suaedoideae.
Suaeda nigra, Suaedoideae.- Unterfamilie Amaranthoideae Burnett: Sie unterscheidet sich von der Unterfamilie Gomphrenoideae durch Staubbeutel mit zwei Theken und vier Pollensäcken. Bislang zählten zur Unterfamilie etwa 57 Gattungen und etwa 330 Arten. Der Schwerpunkt der Verbreitung liegt im tropischen Amerika, im tropischen und südlichen Afrika und in Australien. Die Unterfamilie erwies sich als polyphyletisch, auch ihre bisherigen Untergliederungen (Tribus Amarantheae Rchb. mit den zwei Subtribus Amaranthinae und Aervinae) entsprechen bis auf die Tribus Celosiae nicht echten Verwandtschaftslinien [1]. Daher werden hier noch taxonomische Umgruppierungen erfolgen. Müller & Borsch (2005) unterscheiden mehrere Claden[1]:
- „Basale Gruppe“:
- Bosea L.: Sie kommen auf den Makaronesichen Inseln, Zypern und im westlichen Himalaja vor.
- Charpentiera Gaudich.: Sie kommen auf Pazifikinseln (Hawaii und Tubuai-Inseln) vor.
- Cladus Amaranthoids:
- Amarant (Amaranthus L., Syn.: Acnida L., Amblogyna Raf.): Mit etwa 60 Arten, auch außerhalb der Tropen in gemäßigten Breiten.
- Chamissoa Kunth: Mit etwa 24 Arten in Mittel- und Südamerika.
- Tribus Celosiae: Kennzeichnend sind mehrere Samenanlagen im Fruchtknoten. Diese Tribus ist als einzige monophyletisch[1]. Mit etwa fünf Gattungen und etwa 69 Arten:
- Brandschopf (Celosia L.): Mit etwa 45 bis 60 Arten.
- Deeringia R.Br. (Syn.: Cladostachys D.Don, Dendroportulaca Eggli): Mit etwa sieben Arten.
- Henonia Moq.
- Hermbstaedtia Rchb.: Sie kommen im südlichen Afrika vor.
- Pleuropetalum Hook. f.
- Cladus Aervoids: In tropischen Gebieten von Afrika und Asien:
- Aerva Forssk.: Mit etwa zehn Arten.
- Nothosaerva Wight
- Ptilotus R.Br. (Syn.: Dipteranthemum F.Muell., Trichinium R.Br.): Mit etwa 80 Arten in Australien:
- Cladus Achyranthoids: In tropischen Gebieten von Afrika und Asien und auf Pazifischen Inseln
- Achyranthes L.: Mit etwa 15 Arten, beispielsweise:
- Achyranthes mutica A.Gray
- Nototrichium Hillebr.
- Calicorema Hook. f.
- Cyathula Blume: Mit etwa 27 Arten.
- Pandiaka (Moq.) Hook. f.
- Psilotrichum Blume (Syn.: Psilostachys Hochst.): Mit etwa 14 Arten.
- Pupalia Juss.
- Sericostachys Gilg & Lopr.
- Achyranthes L.: Mit etwa 15 Arten, beispielsweise:
- Zahlreiche Gattungen wurden bislang noch nicht auf ihre Zuordnung untersucht:
- Achyropsis (Moq.) Hook. f.
- Allmania R.Br. ex Wight: Mit der einzigen Art:
- Allmania nodiflora (L.) R.Brown ex Wight: Sie kommt im tropischen Asien vor.
- Allmaniopsis Suess.
- Arthraerva (Kuntze) Schinz
- Centema Hook. f.
- Centemopsis Schinz
- Centrostachys Wall.
- Chionothrix Hook. f.
- Dasysphaera Volkens ex Gilg: Die etwa vier Arten kommen im tropischen Ostafrika vor
- Digera Forssk.
- Eriostylos C.C.Towns.
- Hebanthodes Pedersen
- Herbstia Sohmer (Syn. Siamosia K.Larsen & Pedersen)
- Indobanalia A.N.Henry & B.Roy (Syn.: Banalia Moq.)
- Kyphocarpa (Fenzl) Lopr. (Syn.: Cyphocarpa (Fenzl) Lopr. orth. var.): Sie kommen im südlichen Afrika vor.
- Lagrezia Moq. (Apterantha C.H.Wright)
- Lecosia Pedersen
- Leucosphaera Gilg
- Lopriorea Schinz
- Marcelliopsis Schinz (Syn.: Marcellia Baill.)
- Mechowia Schinz
- Nelsia Schinz
- Neocentema Schinz
- Nyssanthes R.Br.
- Omegandra G.J.Leach & C.C.Towns.
- Pedersenia Holub
- Pleuropterantha Franch.
- Polyrhabda C.C.Towns.
- Pseudosericocoma Cavaco: Mit der einzigen Art:
- Pseudosericocoma pungens (Fenzl) Cavaco: Sie kommt im südwestlichen und südlichen Afrika vor.
- Psilotrichopsis C.C.Towns.: Mit ein bis drei Arten in Thailand und der Malaiischen Halbinsel.
- Quaternella Pedersen: Mit der einzigen Art:
- Quaternella confusa Pedersen: Sie ist in Südamerika verbreitet.
- Rosifax C.C.Towns.: Mit der einzigen Art:
- Rosifax sabuletorum C.C.Towns.: Sie kommt nur in Somalia vor.
- Saltia R.Br. ex Moq. (Syn.: Psilodigera Suess.): Mit der einzigen Art:
- Saltia papposa (Forssk.) Moq.: Sie kommt nur auf der südlichen Arabischen Halbinsel vor.
- Sericocoma Fenzl: Die etwa zwei bis drei kommen im südlichen Afrika vor.
- Sericocomopsis Schinz: Die etwa zwei Arten kommen im tropischen Ostafrika vor.
- Sericorema (Hook. f.) Lopr.: Die etwa zwei Arten kommen im südlichen Afrika vor.
- Stilbanthus Hook. f.: Mit der einzigen Art:
- Stilbanthus scandens Hook. f.: Sie kommt in Asien vor.
- Trichuriella Bennet (Syn.: Trichurus C.C.Towns.): Mit der einzigen Art:
- Trichuriella monsoniae (L. f.) Bennet: Sie kommt in Südostasien vor.
- Volkensinia Schinz: Mit der einzigen Art:
- Volkensinia prostrata Schinz: Sie kommt in Ostafrika vor.
- „Basale Gruppe“:
- Unterfamilie Gomphrenoideae Kostel.: Kennzeichnend sind Staubbeutel mit nur einer Theke und 2 Pollensäcken. Die Unterfamilie bildet nach Sanchez Del-Pino (2009) ein monophyletisches Taxon. Die bisherige Gliederung in zwei Tribus (Gomphreneae und Pseudoplantageae) entspricht aber nicht den phylogenetischen Verhältnissen, statt dessen werden drei Claden erkannt [11]. Das Zentrum der Diversität liegt in Zentral-Amerika, Mexiko und den Trockenwäldern und Dornbuschsavannen von Südamerika[1]. Mit etwa 13 Gattungen und etwa 306 Arten:
- Cladus Iresinoids (Schwestercladus der beiden anderen Claden):
- Irenella Suess.
- Iresine P.Browne (Syn.: Dicraurus Hook. f.): Mit etwa 40 bis 80 Arten.
- Woehleria Griseb.: Mit nur einer Art in Kuba.
- Cladus Alternantheroids:
- Papageienblatt (Alternanthera Forssk., Syn.: Brandesia Mart.): Mit etwa 100 Arten.
- Cladus Gomphrenoids:
- Blutaparon Raf. (ehemals Philoxerus): Mit etwa fünf Arten, beispielsweise:
- Froelichia Moench: Mit etwa zwölf Arten.
- Kugelamarant (Gomphrena L., Syn.: Bragantia Vand.): Mit etwa 120 Arten. Diese Gattung ist polyphyletisch, so dass taxonomische Neugruppierungen zu erwarten sind[11].
- Niedlicher Kugelamarant (Gomphrena pulchella)
- Niederliegender Kugelamarant (Gomphrena decumbens)
- Echter Kugelamarant (Gomphrena globosa)
- Guilleminea Kunth (Syn.: Brayulinea Small)
- Lithophila Sw.
- Pfaffia Mart. (Syn.: Hebanthe Mart.): Mit etwa 35 Arten.
- Pseudogomphrena R.E.Fr.
- Pseudoplantago Suess.: Mit je einer Art in Venezuela und Argentinien.
- Tidestromia Standl.: Mit etwa sechs Arten.
- Xerosiphon Turcz.: Mit etwa zwei Arten in Brasilien.
- Cladus Iresinoids (Schwestercladus der beiden anderen Claden):
- Unterfamilie Betoideae Ulbr.: Mit etwa fünf bis sechs Gattungen [3], zu den enthaltenen Gattungen siehe Hauptartikel Betoideae:
- Cladus Beteae: Mit nur einer Gattung.
- Cladus Hablitzieae: Mit etwa fünf Gattungen.
- Unterfamilie Camphorosmoideae A.J.Scott: Mit nur einer Tribus und etwa 20 Gattungen [4], zu den enthaltenen Gattungen siehe Hauptartikel Camphorosmoideae:
- Tribus Camphorosmeae:
- Bassia/Camphorosma-Clade: Sie ist weit verbreitet in Eurasien und im südlichen Afrika.
- Chenolea-Clade: Mit einem disjunkten Verbreitungsgebiet mit vier Gattungen und etwa fünf Arten.
- Sclerolaena-Clade: Mit etwa 147 Arten aus Australien und etwa drei Arten aus Zentralasien.
- Tribus Camphorosmeae:
- Unterfamilie Chenopodioideae Burnett: Mit drei Tribus und etwa 25 Gattungen, zu den enthaltenen Gattungen siehe Hauptartikel Chenopodioideae:
- Unterfamilie Corispermoideae Ulbr.: Mit nur einer Tribus [9], zu den enthaltenen Gattungen siehe Hauptartikel Corispermoideae:
- Tribus Corispermeae: Mit etwa drei Gattungen [13]. Es sind einjährige Pflanzen.
- Unterfamilie Polycnemoideae Ulbr. [1]: Mit nur einer Tribus [14], zu den enthaltenen Gattungen siehe Hauptartikel Polycnemoideae:
- Tribus Polycnemeae: Mit etwa drei Gattungen.
- Unterfamilie Salicornioideae Kostel.: Mit nur einer Tribus [1][6], zu den enthaltenen Gattungen siehe Hauptartikel Salicornioideae:
- Tribus Salicornieae: Mit etwa elf Gattungen und etwa 90 Arten.
- Unterfamilie Salsoloideae Ulbr.: Mit zwei Tribus [1] und etwa 35 Gattungen, zu den enthaltenen Gattungen siehe Hauptartikel Salsoloideae:
- Unterfamilie Suaedoideae Ulbr. [8][10]: Mit zwei Tribus und zwei oder drei Gattungen, zu den enthaltenen Gattungen siehe Hauptartikel Suaedoideae:
- Tribus Bienertieae Ulbr.: Mit nur einer Gattung.
- Tribus Suaedeae: Mit ein oder zwei Gattungen.
Quellen
- Die Familie der Amaranthaceae bei der APWebsite. (Abschnitte Beschreibung, Photosyntheseweg, Inhaltsstoffe, Systematik)
- Kai Müller & Thomas Borsch: Phylogenetics of Amaranthaceae using matK/trnK sequence data – evidence from parsimony, likelihood and Bayesian approaches, In: Annals of the Missouri Botanical Garden, Volume 92, 2005, S. 66-102.
Einzelnachweise
- ↑ a b c d e f g h i j Kai Müller & Thomas Borsch: Phylogenetics of Amaranthaceae using matK/trnK sequence data – evidence from parsimony, likelihood and Bayesian approaches, In: Annals of the Missouri Botanical Garden, 92, 2005, S. 66-102.
- ↑ Gudrun Kadereit, Thomas Borsch, Kurt Weising, und Helmut Freitag: Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis. In: International Journal of Plant Sciences, Volume 164 (6), 2003, S. 982.
- ↑ a b c Gudrun Kadereit, S. Hohmann & Joachim W. Kadereit: A synopsis of Chenopodiaceae subfam. Betoideae and notes on the taxonomy of Beta, In Willdenowia, Volume 36, 2006, Seite 9-19. pdf-Datei
- ↑ a b c Gudrun Kadereit & Helmut Freitag: Molecular phylogeny of Camphorosmeae (Camphorosmoideae, Chenopodiaceae): Implications for biogeography, evolution of C4-photosynthesis and taxonomy, In: Taxon, Volume 60 (1), 2011, S. 51-78. pdf-Datei
- ↑ a b c d Gudrun Kadereit, Evgeny V. Mavrodiev, Elizabeth H. Zacharias & Alexander P. Sukhorukov: Molecular phylogeny of Atripliceae (Chenopodioideae, Chenopodiaceae): Implications for systematics, biogeography, flower and fruit evolution, and the origin of C4 Photosynthesis, In: American Journal of Botany, Volume 97 (10), 2010, S. 1664-1687.pdf-Datei
- ↑ a b c Gudrun Kadereit, Ladislav Mucina & Helmut Freitag: Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology, In: Taxon, Volume 55 (3), 2006, Seiten 617–642. pdf-Datei
- ↑ a b c d Hossein Akhani, Gerald Edwards & Eric H. Roalson:Diversification Of The Old World Salsoleae S.L. (Chenopodiaceae): Molecular Phylogenetic Analysis Of Nuclear And Chloroplast Data Sets And A Revised Classification, In: International Journal of Plant Sciences, 168 (6), 2007, S. 931–956. pdf-Datei
- ↑ a b Maxim V. Kapralov, Hossein Akhani, Elena V. Voznesenskaya, Gerald Edwards, Vincent Franceschi & Eric H. Roalson: Phylogenetic Relationships in the Salicornioideae / Suaedoideae / Salsoloideae s.l. (Chenopodiaceae) Clade and a Clarification of the Phylogenetic Position of Bienertia and Alexandra Using Multiple DNA Sequence Datasets, In: Systematic Botany, 2006. pdf-Datei
- ↑ a b Alexander P. Sukhorukov: Fruit anatomy and its taxonomic significance in Corispermum (Corispermoideae, Chenopodiaceae). In: Willdenowia, Band 37, 2007, ISSN 0511-9618, S. 63-87, doi:10.3372/wi.37.37103, Fulltext - pdf-Datei.
- ↑ a b Peter Schütze, Helmut Freitag & Kurt Weising: An integrated molecular and morphological study of the subfamily Suaedoideae Ulbr. (Chenopodiaceae), In: Plant Systematics and Evolution, Volume 239, 2003, S. 257-286. Zusammenfassung: doi:10.1007/s00606-003-0013-2
- ↑ a b c Ivonne Sánchez del-Pino, Thomas Borsch & Timothy J. Motle: trnL-F and rpl16 Sequence Data and Dense Taxon Sampling Reveal Monophyly of Unilocular Anthered Gomphrenoideae (Amaranthaceae) and an Improved Picture of Their Internal Relationships, In: Systematic Botany, Volume 34 (1), 2009, S. 57-67. doi:10.1600/036364409787602401
- ↑ Chenopodieae bei GRIN.
- ↑ Corispermae bei GRIN.
- ↑ Polycnemoideae bei GRIN.
Weblinks
 Commons: Fuchsschwanzgewächse – Sammlung von Bildern, Videos und Audiodateien
Commons: Fuchsschwanzgewächse – Sammlung von Bildern, Videos und Audiodateien - Unterfamilie Amaranthoideae Burnett: Sie unterscheidet sich von der Unterfamilie Gomphrenoideae durch Staubbeutel mit zwei Theken und vier Pollensäcken. Bislang zählten zur Unterfamilie etwa 57 Gattungen und etwa 330 Arten. Der Schwerpunkt der Verbreitung liegt im tropischen Amerika, im tropischen und südlichen Afrika und in Australien. Die Unterfamilie erwies sich als polyphyletisch, auch ihre bisherigen Untergliederungen (Tribus Amarantheae Rchb. mit den zwei Subtribus Amaranthinae und Aervinae) entsprechen bis auf die Tribus Celosiae nicht echten Verwandtschaftslinien [1]. Daher werden hier noch taxonomische Umgruppierungen erfolgen. Müller & Borsch (2005) unterscheiden mehrere Claden[1]:













Wikimedia Foundation.