- DsRNA
-
Die Ribonukleinsäure ist eine Nukleinsäure, das heißt eine Kette aus vielen Nukleotiden (ein so genanntes Polynukleotid). Im internationalen und im wissenschaftlichen Sprachgebrauch wird die Ribonukleinsäure mit der englischen Abkürzung RNA (ribonucleic acid) bezeichnet, im deutschen Sprachraum auch mit RNS.
Eine wesentliche Funktion der RNA in der Zelle ist die Umsetzung von genetischer Information in Proteine. RNA fungiert hierbei als Informationsträger in Form der mRNA (bei RNA-Viren macht sie sogar das Genom selbst aus). Weiterhin bestehen auch die für die Umsetzung dieser Information verantwortlichen Zellbestandteile im Wesentlichen aus RNA: Bei der Reifung der mRNA sind snRNA und snoRNA beteiligt, die katalytischen Bestandteile der Ribosomen bildet die rRNA, und die tRNA transportiert die Bausteine für die Proteine. Ferner sind spezielle RNAs an der Genregulation beteiligt.
 RNA und DNA im Vergleich
RNA und DNA im VergleichInhaltsverzeichnis
Aufbau und Unterschied zur DNA
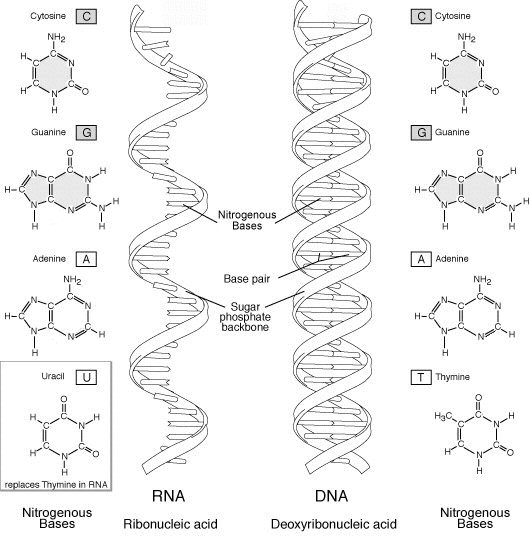
Vom Aufbau her ist die RNA der DNA ähnlich. RNA-Moleküle sind – im Gegensatz zur doppelsträngigen DNA – in der Regel einzelsträngig. Beide sind Polynukleotide, bei denen die Nukleobasen (Adenin, Guanin, Cytosin, Uracil bzw. Thymin bei der DNA) an Zuckern (Ribose für RNA; Desoxyribose für DNA) über Phosphorsäurediester miteinander verknüpft sind. Die Einzelsträngigkeit erhöht die Zahl der Möglichkeiten für dreidimensionale Strukturen der RNA und erlaubt ihr chemische Reaktionen, die der DNA nicht möglich sind. Jedes Nukleotid besteht bei der RNA aus einer Ribose (d. h. einer Pentose: einem Zucker mit fünf C-Atomen), einem Phosphatrest und einer organischen Base. Die Ribose (Zucker) der RNA ist mit derjenigen der DNA identisch, bis auf eine Hydroxylgruppe (statt eines Wasserstoff-Atoms, engl. Hydrogen) an der 2'-Position im Pentose-Ring (daher auch Desoxyribonucleinsäure DNA). Dieser Unterschied macht RNA weniger stabil als DNA, da es eine Hydrolyse durch Basen ermöglicht: Die OH-Gruppe an der 2'-Position des Zuckers wird durch ein negativ geladenes Hydroxidion einer Base ihres Protons beraubt und der dann zurückgebliebene Sauerstoff geht eine Ringbindung mit dem Phosphor ein, wodurch die Bindung zum nächsten Nukleotid jedoch gelöst wird. Die RNA wird so wieder in ihre Nukleotide zerlegt.
In der RNA kommen die folgenden organischen Basen vor: Adenin, Guanin, Cytosin und Uracil. Die ersten drei Basen kommen auch in der DNA vor. Uracil dagegen ersetzt Thymin als komplementäre Base zu Adenin. Vermutlich nutzt RNA Uracil, da dieses energetisch weniger aufwändig herzustellen ist (keine Methyl-Substituierung).
Als Sekundärstrukturen sind bei der RNA vor allem Hairpin-, Stemloop- und Loop-Strukturen bekannt, eine Helix-Konformation ist aber ebenfalls möglich, wobei Hairpin- und Stemloop-Strukturen sowohl Einzelstrang- als Doppelstrangbereiche aufweisen. Die Loop-Strukturen bezeichnen einzelsträngige Schlaufenstrukturen innerhalb eines Moleküls.
RNA kann, wie DNA ebenfalls als doppelsträngiges Molekül vorliegen. Sie weist dabei die typischen Merkmale einer Watson-Crick-Helix auf: antiparallele Anordnung der RNA-Stränge und rechtsgewundene Helix. Sie nimmt dabei die Form einer A- oder A´-Helix an (Vgl. DNA). Die A-RNA wird auch als RNA-11 bezeichnet, homolog zur A´-RNA, die als RNA-12 bezeichnet wird. Hierbei gibt die Zahl nach dem Spiegelstrich die Anzahl der Basenpaare je Helixwindung wieder. A´-RNA kommt häufig bei hohen Salzkonzentrationen vor (über 20 %).
A-RNA: 11 Basenpaare pro Helixwindung, Ganghöhe 2,7 nm bis 2,8 nm, Neigungswinkel zur Helixachse ca. 14°
A´-RNA: 12 Basenpaare pro Helixwindung, Ganghöhe 3 nm, Neigungswinkel zur Helixachse 16° bis 19°Synthese von RNA
Das Enzym RNA-Polymerase katalysiert an der DNA durch den Prozess der Transkription aus Nukleosidtriphosphat (NTP) die RNA. Dafür setzt sich die RNA-Polymerase an eine Promotor genannte DNA-Sequenz (Transkriptionsinitiation). Dann trennt sie die DNA-Doppelhelix durch Lösen der Wasserstoffbrücken in einem kurzen Bereich in zwei DNA-Einzelstränge auf. Am codogenen Strang der DNA lagern sich durch Basenpaarung komplementäre Ribonukleotide an. Sie werden unter Eliminierung eines Pyrophosphat durch eine esterartige Bindung zwischen Phosphorsäure und Ribose miteinander verknüpft. Die Ableserichtung der DNA verläuft vom 3'-Ende zum 5'-Ende, die Synthese der komplementären RNA dem entsprechend 5'→3'. Die Öffnung der DNA-Doppelhelix erfolgt nur in einem kurzen Bereich, so dass der bereits synthetisierte Teil der RNA aus dieser Öffnung heraushängt und zwar mit dem 5'-Ende der RNA voran. Die Synthese der RNA wird an einer Terminator-Sequenz beendet. Danach wird das RNA-Transkript entlassen und die RNA-Polymerase löst sich von der DNA.
Biologische Bedeutung
RNA-Moleküle können unterschiedliche Funktionen ausüben. Zum einen kann RNA genetische Information übertragen. Andere RNA-Moleküle tragen zur Übersetzung dieser Information in Proteine bei, sowie bei der Regulation der Gene. Darüber hinaus kann RNA auch katalytische Funktionen ähnlich einem Enzym innehaben. RNA wird daher – je nach ihrer Funktion – auch verschieden benannt. Vorangestellte Kleinbuchstaben kennzeichnen die unterschiedlichen RNA-Typen:
- Die mRNA, Boten-RNA (engl. messenger RNA) kopiert die in einem Gen auf der DNA liegende Information und trägt sie zum Ribosom, wo mit Hilfe dieser Information die Proteinbiosynthese stattfinden kann. Jeweils drei im Leseraster des Polynukleotidstrang nebeneinander liegende Nukleotide bilden ein Codon, mit dessen Hilfe sich eine spezifische Aminosäure, die in ein Protein eingebaut werden soll, eindeutig bestimmen lässt. Dieser Zusammenhang wurde 1961 von Heinrich Matthaei und Marshall Warren Nirenberg gefunden. Die Entschlüsselung des genetischen Codes markiert einen Neubeginn in fast allen Bio-Wissenschaften.
Die folgenden RNA-Klassen werden allgemein als non-coding RNA (nicht Protein-kodierend) bezeichnet.
- Die tRNA, Transfer-RNA kodiert keine genetische Information, sondern dient als Hilfsmolekül bei der Proteinbiosynthese, indem sie eine einzelne Aminosäure aus dem Cytoplasma aufnimmt und zum Ribosom transportiert. Die tRNA wird durch ein bestimmtes 'RNA-Gen' kodiert.
- Die rRNA, ribosomale RNA, trägt, ähnlich wie die tRNA, keine genetische Information, sondern ist am Aufbau des Ribosoms beteiligt und ist bei der Knüpfung der Peptidbindung auch katalytisch aktiv.
- Die hnRNA, heterogene Kern-RNA (engl. heterogeneous nuclear RNA), kommt im Zellkern von Eukaryoten vor und ist eine Vorstufe der reifen mRNA, häufig wird sie daher auch als prä-mRNA (oder engl. pre-mRNA für precursor mRNA) bezeichnet.
- Die snRNA, small nuclear-RNA, im Zellkern von Eukaryoten, ist verantwortlich für die Prozessierung der hnRNA im Spliceosom.
- Die snoRNA, small nucleolar-RNA, finden sich im Nukleolus, und die eng verwandten scaRNAs in den Cajal Bodies.
- Die siRNA, small interfering RNA, entsteht bei einem Signalweg der Zelle, der als RNAi (RNA Interference) zusammengefasst wird. Dabei wird dsRNA (doppelsträngige RNA; englisch double-stranded RNA) durch das Enzym Dicer in viele kleinere Fragmente von ca. 22 Nukleotiden Länge zerteilt (die siRNAs) und in den Enzymkomplex RISC (RNA-induced silencing complex) eingebaut. Mithilfe der inkorporierten RNA-Fragmente bindet RISC komplementär an DNA, z.B. Genbereiche, oder mRNA und kann diese damit „abschalten“. siRNA's werden aktuell (2006) intensiv auf ihre Beteiligung an verschiedenen Zellvorgängen und Krankheiten erforscht.
- Die microRNAs sind eng verwandt mit den siRNAs und dient der Regulation zellulärer Prozesse wie z.B. Proliferation und Zelltod.
- Die asRNA, antisense-RNA, dient der Regulation der Genexpression.
- Die Riboswitches dienen der Genregulation. Sie können entweder aktivierend oder reprimierend wirken.
- Die Ribozyme sind katalytisch aktive RNA-Moleküle. Sie katalysieren wie Enzyme chemische Reaktionen.
In der Mehrzahl der Lebewesen spielt die RNA als Informationsträger eine der DNA untergeordnete Rolle: Die DNA ist hier das permanente Speichermedium für die genetische Information, die RNA dient als Zwischenspeicher. Nur RNA-Viren (die Mehrzahl aller Viren) nutzen RNA anstelle der DNA als permanentes Speichermedium. Zur Taxonomie von Viren unterscheidet man folgende RNA-Typen:
- dsRNA: Doppelstrang-RNA
- ss(+)RNA: Einzelstrang-RNA, die als mRNA verwendet wird.
- ss(−)RNA: Einzelstrang-RNA, die als Matrize zur mRNA-Produktion dient
Darüber hinaus nutzen einige Viren die RNA als Replikationsintermediat (z.B. Retroviren und Hepadnaviren)
Abbau von RNA
Da ständig neue RNA gebildet wird und da zu unterschiedlichen Zeitpunkten verschiedene Transkripte benötigt werden (differentielle Genexpression), darf die RNA in der Zelle nicht zu stabil sein, sondern muss auch einem Abbau unterliegen. Dies geschieht mit Hilfe von RNasen, Enzymen, die die Verbindungen des Zucker-Gerüstes der RNA trennen und somit die Monomere (bzw. Oligomere) bilden, welche wieder zur Bildung neuer RNA verwendet werden können. Wann eine RNA abgebaut werden soll, wird dabei vor allem (aber nicht ausschließlich) durch die Länge des Poly-A-Schwanzes bestimmt, der mit zunehmender Verweildauer der RNA im Cytoplasma sukzessive verkürzt wird. Sinkt die Länge dieses Schwanzes unter einen kritischen Wert wird die RNA schnell degradiert. Zusätzlich können die RNAs stabilisierende oder destabilisierende Elemente enthalten, die eine weitere Regulation ermöglichen.
Zumindest bei der mRNA von Eukaryoten findet der RNA-Abbau nicht irgendwo im Cytoplasma statt, sondern in den so genannten „P-Bodies“ (Processing Bodies), die sehr reich an RNasen und anderen, am RNA-turnover (-Abbau)-beteiligten Enzymen sind. Zusammen mit „Stress-Granules“ dienen diese Körper weiterhin der kurzzeitigen Lagerung von mRNA und demonstrieren so wiederum die enge Verknüpfung des RNA-Metabolismus (hier Translation und RNA-Abbau).
Die RNA-Welt-Hypothese
Die RNA-Welt-Hypothese wurde erstmals 1986 von Walter Gilbert vorgeschlagen und besagt, dass RNA-Moleküle in der chemischen Evolution die Vorläufer der Organismen waren.
Die Hypothese lässt sich ableiten aus der Fähigkeit der RNA zur Speicherung, Übertragung und Vervielfältigung genetischer Informationen sowie aus ihrer Fähigkeit, als Ribozyme Reaktionen zu katalysieren. In einer Evolutionsumgebung würden diejenigen RNA-Moleküle gehäuft vorkommen, die sich selbst bevorzugt vermehren.
Ausgangspunkt sind einfache sich selbst replizierende RNA-Moleküle. Einige davon erhalten die Eigenschaft, die Synthese von Proteinen zu katalysieren, die selbst wieder die Synthese der RNA und ihre eigene Synthese katalysieren (Entwicklung der Translation). Einige RNA-Moleküle lagern sich zu doppelsträngigen RNA-Molekülen zusammen, die sich zu DNA-Molekülen und Trägern der Erbinformation weiterentwickeln (Entwicklung der Transkription).
Als Grundlage dienen bestimmte RNA-Moleküle, die von beliebigen RNA-Vorlagen und damit von sich selbst Kopien erzeugen können. Jennifer A. Doudna und Jack W. Szostak benutzten als Vorlage zur Entwicklung dieses RNA-Typs das selbst-spleißende Intron des eukaryotischen Einzellers Tetrahymena thermophila. Damit besteht die Möglichkeit, dass in den Ribosomen die eigentlich katalytischen Moleküle die rRNA sind und somit RNA die Eiweißsynthese katalysiert. Einschränkungen bestehen allerdings darin, dass bei der selbstreplizierenden RNA als Bausteine nicht Mononukleotide sondern Oligonukleotide und Hilfsstoffe benötigt werden. 2009 identifizierten Lincoln und Joyce allerdings ein Ribozym, dass sich aus Mononukleotiden repliziert [1].
2001 wurde entdeckt, dass die wichtigen katalytischen Zentren der Ribosomen von RNA und nicht, wie vorher angenommen, von Proteinen gestellt werden. Dies zeigt, dass eine katalytische Funktion der RNA, wie sie in der RNA-Welt-Hypothese vorgeschlagen wurde, heute von Lebewesen genutzt wird. Da Ribosomen als sehr ursprüngliche Zellbausteine gelten, gilt diese Entdeckung als wichtiger Beitrag zur Untermauerung der RNA-Welt-Hypothese. Man ist nun sicher, dass RNA-Moleküle - zumindest prinzipiell - in der Lage sind, Aminosäuren zu Proteinen zu verketten. In diesem Zusammenhang ist auch die PNA (Peptid-Nukleinsäure) als mögliches Vorläufermolekül der RNA von Interesse.
Siehe auch: Quasispezies, Manfred Eigen
Nobelpreise
Für die Forschung an RNA sind bereits mehrere Nobelpreise verliehen worden. Schon 1959 erhielten S. Ochoa und A. Kornberg den Nobelpreis für Medizin für ihre Studien zur Synthese der RNA durch RNA-Polymerasen. Für die Entdeckung der katalytischen Aktivität von RNA Molekülen (vgl. Ribozym) wurden S. Altman und T. Cech 1989 mit dem Nobelpreis für Chemie geehrt. 1993 schließlich erhielten R. Roberts und P. Sharp den Nobelpreis für Medizin für ihre Studien zur Prozessierung der RNA in Eukaryoten (siehe auch Splicing). Und zu guter Letzt bekamen im Jahr 2006 Andrew Fire und Craig Mello den Nobelpreis für Medizin für die Entdeckung der RNA-Interferenz, fast gleichzeitig wurde Roger Kornberg (der Sohn des früheren Preisträgers A. Kornberg) für seine Studien zur RNA-Polymerase geehrt.
Literatur
- J. Marx: P-Bodies Mark the Spot for Controlling Protein Production. In: Science. Bd. 310, Nr. 5749, S. 764–765. 2005, PMID 16272094 doi:10.1126/science.310.5749.764.
- Seyffert: Lehrbuch der Genetik. 2. Auflage, S. 42. Spektrum Akademischer Verlag, 2003.
- Albert Gossauer: Struktur und Reaktivität der Biomoleküle - Eine Einführung in die organische Chemie. S. 525. Wiley-VCH, 2006.
- Brosius, J. & Tiedge, H. (2004): RNomenclature. In: RNA Biol. 1(2):81-83. PMID 17179746 PDF
Einzelnachweise
- ↑ Lincoln, Tracey; Joyce, Gerald F. (January 8, 2009): "Self-Sustained Replication of an RNA Enzyme". Science (New York: American Association for the Advancement of Science). doi:10.1126/science.1167856. ISSN 1095-9203. PMID 19131595. http://www.sciencemag.org/cgi/content/abstract/1167856. Abgerufen am 13.1.2009. http://www.sciencemag.org/cgi/content/abstract/1167856 Bd. 409, S. 797-801. 2001.
Siehe auch
Weblinks
Wikimedia Foundation.