- Afrikanische Urmutter
-
 Mitochondrium (schematischer Aufbau)
Mitochondrium (schematischer Aufbau)Die Mitochondriale Eva (engl. Mitochondrial Eve) ist ein Begriff aus der Archäogenetik und bezeichnet die Frau, aus deren mitochondrialer DNA (mtDNA) die mitochondriale DNA aller heute lebenden Menschen hervorgegangen ist. Ihr Gegenstück ist der Adam des Y-Chromosoms.
Inhaltsverzeichnis
Theorie
Dass Wissenschaftler von der Annahme ausgehen, eine solche Vorfahrin habe jemals existiert, beruht auf der Gendrift, einem Phänomen in der Populationsgenetik: Im Verlauf der Evolution nimmt die Häufigkeit der unterschiedlichen Varianten eines DNA-Moleküls (Allele) in der Population durch Zufall zu oder ab.
Man betrachte eine Modell-Population mit 100 Frauen, bei der 50 ein Allel A der mtDNA in sich tragen und 50 ein Allel B. In der folgenden Generation sind die Anteile durch den zufälligerweise etwas größeren Reproduktionserfolg der „Allel-A-Frauen“ geringfügig verschoben, z. B. 51-mal Allel A und nur 49-mal Allel B, in der darauf folgenden Generation liegen 48-mal Allel A und 52-mal Allel B vor. Statistisch haben nach etwa 100 Generationen alle Individuen in der Population entweder Allel A oder B. Eines der Allele ist infolge des Gendrifts in der Population ausgestorben und das andere wurde fixiert.
Im Unterschied zur Selektion, die dazu führt, dass nur die für das Überleben vorteilhaften Allele im Genpool verbleiben, führt Gendrift zum zufälligen Aussterben oder Überleben bestimmter Allele in einer Population. Allgemein erfolgt die Fixierung durch Gendrift viel langsamer als durch positive Selektion. Je größer die effektive Populationsgröße, desto geringer die Gendrift und desto länger dauert die Fixierung eines Allels. Gendrift wird manchmal auch Gründereffekt genannt, weil sie bei kleinen Populationen, wie Gründerpopulationen, stärker zur Geltung kommt.
Die Gendrift führt jedoch in jeder Population zu einer Reduktion der genetischen Vielfalt. In realen Populationen bleibt die genetische Diversität jedoch erhalten, da aus Mutationen ständig neue Allele hervorgehen. In der oberen Modell-Population sind nach etwa 100 Generationen, alle mtDNAs in der Population Kopien desselben DNA-Moleküls, welches etwa 100 Generationen zuvor von einer einzigen Frau an ihre Töchter vererbt wurde. Sie unterscheiden sich lediglich durch die Mutationen die seitdem aufgetreten sind. Sie sind nicht identisch aber monophyletisch.
Die Mitochondriale Eva stellt weder die erste Frau noch die einzige Frau zu einem bestimmten Zeitpunkt der Vergangenheit dar. Eva hatte viele Zeitgenossinnen; die mitochondrialen Erblinien der anderen Frauen starben aber aus, während die von Eva überlebte. Lebenszeit und -ort dieser Vorfahrin lassen sich mit Hilfe der Analyse der mitochondrialen DNA einer repräsentativen Anzahl heute lebender Individuen recht genau eingrenzen.
Bedeutung
Eine Reihe von Eigenschaften machen die mitochondriale DNA zu einem wertvollen Werkzeug für die Erforschung der menschlichen Abstammung.
- Im Vergleich zur DNA des Zellkerns zeigt die mtDNA eine höhere Mutationsrate.
- Da mtDNA nur von der Mutter weiter vererbt wird, ist die effektive Populationsgröße nur ¼ so groß wie bei der autosomalen DNA, bei der beide Elternteile je zwei Kopien tragen. Dementsprechend erfolgt die Fixierung von Allelen auch etwa vier mal so schnell. Die Allele menschlicher mtDNA sind also wesentlich jünger als bei der autosomalen DNA und eignen sich sehr gut (und auch nur) für die Erforschung der jungen Menschheitsgeschichte, wie zum Beispiel der Besiedlung Eurasiens.
- Die im Vergleich zur autosomalen DNA höhere Mutationsrate und Gendrift der mtDNA führen dazu, dass die Häufigkeit der Allele von einer Teilpopulation zur anderen viel stärker schwankt. Aus diesen Unterschieden können Aussagen über Abstammung, Migration, Verdrängung oder Vermischung von Populationen viel einfacher abgeleitet werden, als mit der geographisch homogeneren autosomalen DNA.
- Da eine Zelle viele Mitochondrien enthält und in jeder mehrere Kopien der mtDNA vorliegen, lässt sich oft auch aus Fossilien (z. B. Neandertaler-Knochengewebe) genug mtDNA für die Analyse extrahieren, während die DNA des Zellkerns weitaus seltener ausreichend vollständig überliefert ist.
- Das Fehlen von Rekombination bei der Vererbung der mtDNA ermöglicht Aussagen über spezifische Eigenschaften der weiblichen Erblinie.
Wo und wann lebte Eva?
Erste Untersuchungen der Variation humaner mitochondrialer DNA wurden bereits 1983 durchgeführt.[1][2] Richtig bekannt wurde die Theorie aber erst durch eine Publikation von Cann et al. (1987).[3]
Cann et al. (1987) extrahierten mtDNA aus der Plazenta von Personen aus unterschiedlichen Teilen der Welt. Sie sequenzierten die mtDNA nicht, sondern führten eine Untersuchung mittels Restriktionsfragmentlängenpolymorphismus (RFLP) durch. Sie ordneten die mtDNAs entsprechend ihrer Ähnlichkeit auf einem Stammbaum an und ermittelten schließlich die Wurzel des Stammbaums. Von der ermittelten Wurzel zweigten zwei Haupt-Äste ab: Auf dem einen fanden sich nur Afrikaner, auf dem anderen Personen aus allen Erdteilen. Daraus schlossen die Autoren, dass die Mitochondriale Eva in Afrika gelebt haben muss.
Zudem versuchten sie, mit Hilfe einer molekularen Uhr zu ermitteln, wann die Mitochondriale Eva gelebt hatte. 1987 lagen bereits Daten über die mitochondriale DNA von Fischen, Vögeln und einigen Säugetierarten vor. Aus diesen Daten ging hervor, dass mtDNA sich etwa um 2–4 % pro Million Jahren verändert. Diese Daten wurde zur Kalibrierung der molekularen Uhr verwendet. Da die menschlichen mitochondrialen DNAs in der Studie sich durchschnittlich nur um 0,57 % unterschieden, ergab sich daraus, dass die Mitochondriale Eva vor nur etwa 200.000 Jahren gelebt haben muss.
Da die Fixierung einer Erblinie in einer expandierenden Population unwahrscheinlich ist, argumentierten die Autoren, muss die Mitochondriale Eva gelebt haben, bevor die Vorfahren aller heute lebenden Menschen Afrika verlassen hatten. Für die Autoren ein klarer Hinweis für die Out-of-Africa-Theorie. Die „Multiregionale Theorie“ wurde von den Autoren zurückgewiesen, da die Mitochondriale Eva hierfür sehr viel älter hätte sein müssen (Homo erectus hatte Afrika vor fast 2 Millionen Jahren verlassen).
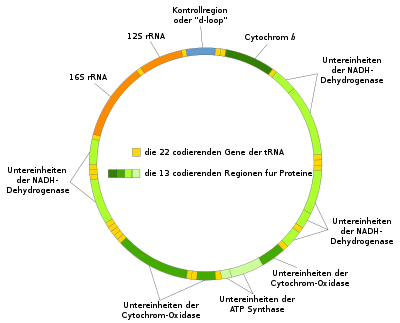 Schematische Darstellung der mtDNA
Schematische Darstellung der mtDNADie Publikation wurde von Anfang an stark kritisiert. Viele Kritikpunkte waren durchaus berechtigt:
- Von den 147 Personen in der Studie stammten nur zwei der 20 „Afrikaner“ wirklich aus Afrika südlich der Sahara. Die anderen 18 waren Afroamerikaner.
- Die Methode, den Stammbaum zu generieren, lieferte nicht unbedingt den statistisch günstigsten Baum.
- Um die Wurzel des Baums zu finden, setzte man die Wurzel in die Mitte des längsten Astes (midpoint rooting). Das kann zu einer falschen Position der Wurzel führen, z. B. wenn die Evolutionsgeschwindigkeit in Afrika höher ist.
- RFLP ist wenig geeignet, um Mutationsraten zu bestimmen, was für eine Molekulare Uhr wichtig ist.
- Schlechte statistische Analyse
Die (seriöse) Kritik war also nicht allgemein gegen das Konzept der Mitochondrialen Eva gerichtet. Jedem, der mit der Materie vertraut war, war klar, dass diese Frau irgendwann und irgendwo gelebt haben muss. Nur das wissenschaftliche Vorgehen der Autoren wurde kritisiert.
Spätere, verbesserte Studien bestätigten und untermauerten jedoch die wichtigsten Aussagen von Cann et al. (1987). Zum Beispiel führten Ingman et al. (2000) eine neue, verbesserte Studie durch.
- Sie nahmen Proben von 53 Personen, 32 von ihnen aus unterschiedlichen Teilen Afrikas südlich der Sahara.
- Sie sequenzierten die kompletten mtDNAs, aber für die Analyse schlossen sie die schnell evolvierende D-Loop-Region aus.
- Die Wurzel des Stammbaums wurde mit Hilfe der mtDNA eines Schimpansen bestimmt (outgroup rooting).
Die Ergebnisse dieser verbesserten Studie waren noch eindeutiger als bei der Studie 1987:
- Vollständige Separation von Afrikanern und Nicht-Afrikanern.
- Die ersten drei Äste des Stammbaums führten nur zu Afrikanern, der vierte führte zu Afrikanern und nicht-Afrikanern.
- Lange Äste in Afrika, aber sternförmige Struktur außerhalb (charakteristisch für kürzliche Expansion).
- 175.000 ± 50.000 Jahre bis zum gemeinsamen Vorfahren (Mitochondriale Eva aller Menschen in der Studie).
- 52.000 ± 28.000 Jahre zur Verzweigung zwischen dem letzten afrikanischen und dem nicht-afrikanischen Zweig (Mitochondriale Eva aller nicht-Afrikaner in der Studie).
- Signal für eine Expansion im nicht-afrikanischen Ast vor etwa 1925 Generationen. Also etwa vor 38.500 Jahren, wenn man von einer Generationszeit von 20 Jahren ausgeht.
Haplotypen
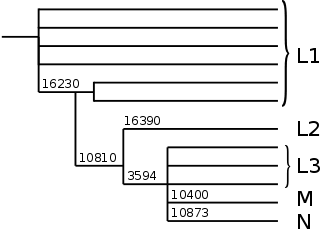 Grober Stammbaum der mitochondrialen DNA beim Menschen. Die Zahlen geben die Position der Mutationen an.
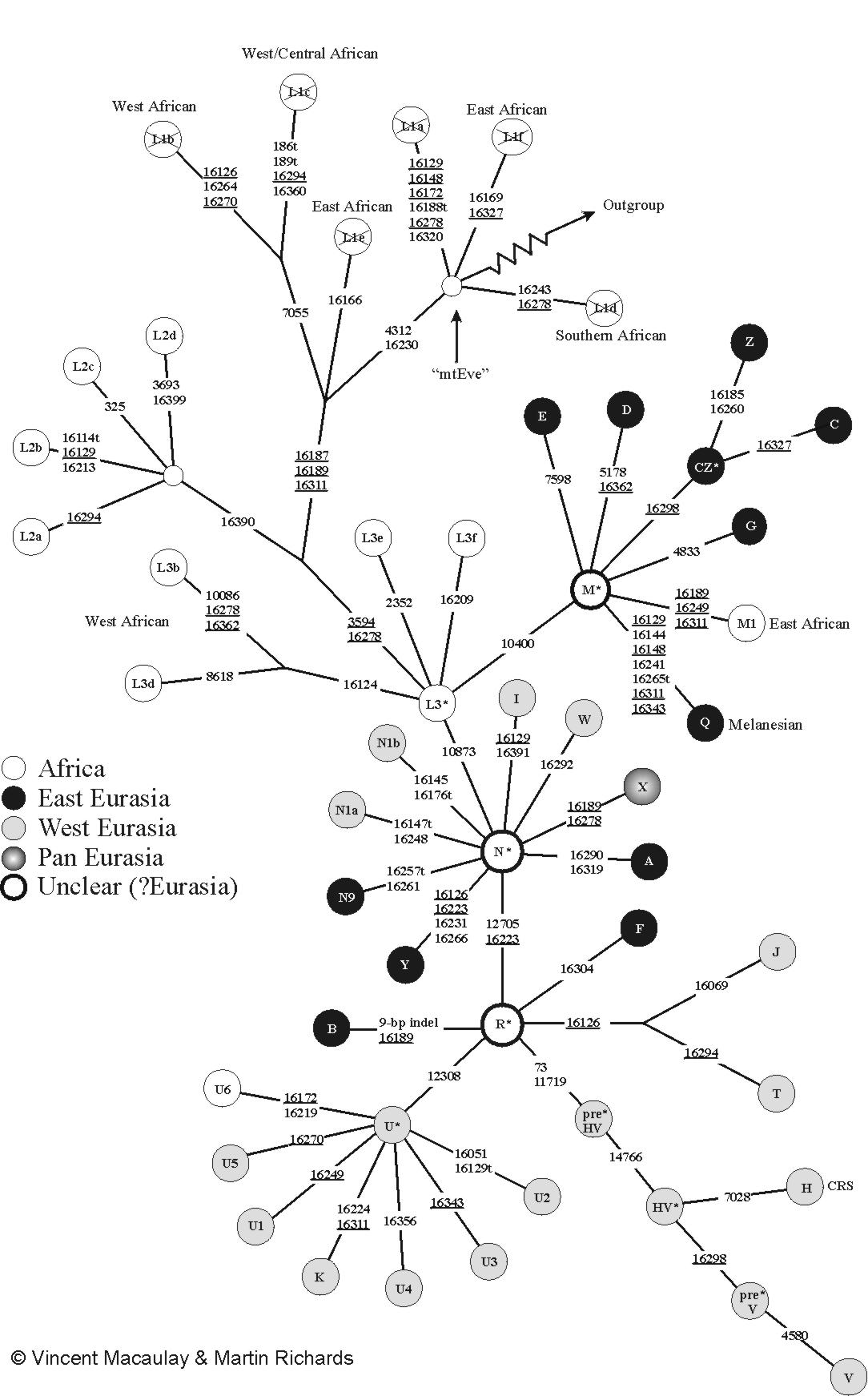
Grober Stammbaum der mitochondrialen DNA beim Menschen. Die Zahlen geben die Position der Mutationen an.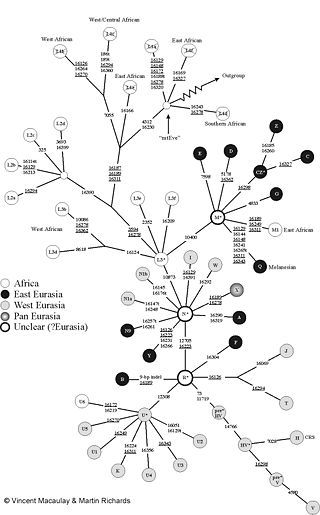 Detaillierter Stammbaum menschlicher mitochondrialer DNA
Detaillierter Stammbaum menschlicher mitochondrialer DNA
Die Zahlen geben die Position der Mutationen an. „mtEve“ ist die Mitochondriale Eva. „Outgroup“ führt zu mtDNA anderer Primaten (z. B. Schimpansen). Die Abbildung benutzt die übliche (falsche) Nomenklatur mit der „L1 Haplogruppe“: L1 bildet jedoch die Wurzel (L1a ist mit L1f nicht näher verwandt als mit V!). Daher wurden die L1-Felder durchgestrichen. Von Macaulay und Richards.Die mitochondriale DNA der Menschen kann in sogenannte Haplogruppen unterteilt werden. Eine Haplogruppe kann ihrerseits weitere Unter-Haplogruppen enthalten, die sich ihrerseits weiter unterteilen lassen. Man versucht, bei der Nomenklatur der Haplogruppen diese Baumstruktur abzubilden und verwendet abwechselnd Buchstaben und Zahlen. Zwei mtDNAs einer Haplogruppe sind dabei stets monophyletisch. Für die Zuordnung verwendet man charakteristische Mutationen in den Gensequenzen der mtDNA, außerhalb des D-Loops.
Eine Person kann z. B. die Haplogruppe C1a3b2 haben. Seine mtDNA ist dann eng mit einer anderen Person verwandt die z. B. C1a3b4 hat. Natürlich teilt seine mtDNA auch eine gemeinsamen Vorfahrin mit einer dritten Person, die C1a3c5 hat, aber diese gemeinsame Vorfahrin hatte früher gelebt, noch bevor sich die C1a3-Linie aufgespalten hatte. Das heißt, C1a3b4 und C1a3b2 sind gegenüber C1a3c5 monophyletisch. Ebenso sind C1a3b2 und C1a3c5 monophyletisch gegenüber allen H-Haplotypen usw.
Leider ist die Nomenklatur relativ inkonsequent realisiert. Viele Buchstaben wurden benutzt, um die wichtigsten nicht-afrikanischen Haplogruppen zu benennen. Viele alte Haplogruppen kommen jedoch in Afrika vor. Diese bezeichnet man zusammen als „L“ und geht bereits für die Unterteilung der Hauptgruppen zu Ziffern über. Über die Zuordnung mancher afrikanischer Haplotypen (in L1 und L3) besteht bis heute noch kein wissenschaftlicher Konsensus.
Wenn man von der Wurzel anfängt, besteht der mitochondriale Stammbaum des Menschen zunächst aus einer Reihe tiefer Äste. Diese genetischen Linien werden heute L1 genannt. Anders als früher gedacht, ist L1 keine monophyletische Haplogruppe, sondern bildet die Wurzel. L1 sind also eigentlich ein ganzes Paket afrikanischer Haplogruppen, die ähnlich alt sind wie die Mitochondriale Eva und deren genaue verwandtschaftliche Beziehung untereinander noch nicht genau geklärt ist.
Von diesen alten L1-Ästen zweigt ein Ast durch eine Mutation an der Position 10810 ab. Von diesem spaltet sich seinerseits die Haplogruppe L2 durch eine Mutation an der Position 16390 ab. Auch L2 kommt praktisch nur bei Afrikanern südlich der Sahara vor.
Eine Mutation an der Position 3594 bildet den Ast, auf dem die großen Haplogruppen M und N sowie noch zahlreiche weitere afrikanische Haplogruppen, die man heute noch unter L3 und (?) zusammenfasst, liegen. L3 ist, wie L1, keine echte (monophyletische) Haplogruppe. Die Haplogruppen M und N kommen beim aller größten Teil der Nicht-Afrikaner vor. Sie sind in Afrika südlich der Sahara sehr selten, wo L1, L2 und L3 dominieren.
Die Haplogruppe M wird in die großen Haplogruppen M1, Z, C, D, E, G und Q unterteilt. Die Haplogruppe N in N1a, N1b, N9, A, I, W, X und Y, sowie in die Haplogruppe R die die Unter-Haplogruppen B, F, H, P, T, J, U und K bildet.
Die derzeit umfangreichste Untersuchung von mitochondrialer DNA wurde vom Genographic Consortium durchgeführt (s. a. The Genographic Project). In diesen Vergleich wurden 78.590 genotypische Proben einbezogen und die mitochondrialen Haplogruppen (und deren Untergruppen) wurden in einem phylogenetischen Baum dargestellt.[4]
Geographische Verteilung
Die „alten“ Haplotypen aus den L-Ästen dominieren in Afrika südlich der Sahara. Es bestehen keine Zweifel, dass sie ihren Ursprung dort haben. Diese Haplotypen finden sich auch in Nordafrika (ca. 50 % Häufigkeit) und, in geringer Häufigkeit, in Europa und Westasien.
Die Haplogruppen M und N dominieren im Rest der Welt und sind in Afrika südlich der Sahara selten. Spezielle Varianten der Haplogruppe M (M1) kommen mit einer Häufigkeit von etwa 20 % in Äthiopien vor. Daher kann angenommen werden, dass auch die Haplogruppe M ihren Ursprung in Afrika hat.
Bei Amerikanischen Ureinwohnern kommen die Haplogruppen A, B, C, D und X vor. A, B, und X gehören zur Haplogruppe N; C und D gehören zu Haplogruppe M.
In Europa und Westasien ist Haplogruppe M sehr selten. Die Haplogruppe N dominiert klar. Die häufigsten Untergruppen gehören in die Untergruppe R: H, V, T, J, U und K. Daneben kommen auch die Haplogruppen I, W und X mit einer signifikanten Häufigkeit vor. In Europa, dem Kaukasus und dem Nahen Osten finden sich praktisch die gleichen Haplogruppen, nur die Häufigkeiten der einzelnen Haplogruppen schwankt. Vor allem die Haplogruppe H ist im Nahen Osten und im Kaukasus deutlich weniger häufig als in Europa (~25 % versus ~45 %), während die Haplogruppe K deutlich häufiger ist. Innerhalb Europas schwanken die Häufigkeiten der Haplogruppen je nach Region geringfügig.
Süd- und Ostasien unterscheiden sich bei den Haplogruppen sehr stark von Westasien. Hier kommen, aus der Haplogruppe M, die Haplogruppen C, D, E, G, Z und Q vor. Die Haplogruppe N kommt hier auch vor, allerdings ist sie vor allem durch die Haplogruppen A, B, F, Y und X vertreten.
Die Haplogruppe X ist bemerkenswert, da sie in ganz Eurasien und Nordamerika vorkommt, wenn auch nur mit relativ geringer Häufigkeit. Früher wurde angenommen, dass die Haplogruppe X in Europa entstand und nur in Europa vorkommt. Als die Haplogruppe bei amerikanischen Ureinwohnern entdeckt wurde, kam die Hypothese auf, sie sei vor Jahrtausenden von Europa aus auf dem Seeweg durch europäische Emigranten nach Amerika gelangt. Mittlerweile wurde Haplogruppe X jedoch auch in Asien entdeckt, was diese Hypothese hinfällig macht (Derneko et al., 2001).
Alte mitochondriale DNA
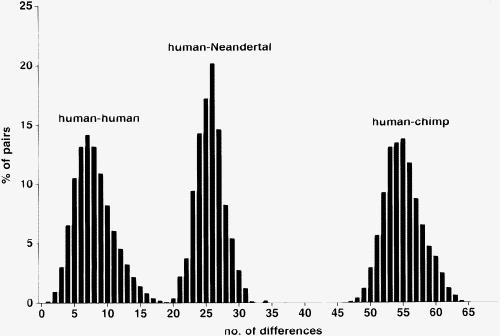 Unterschiede in den Sequenzen der mtDNA zwischen Mensch, Neandertaler und Schimpanse.
Unterschiede in den Sequenzen der mtDNA zwischen Mensch, Neandertaler und Schimpanse.Jede Zelle enthält viele Mitochondrien, die ihrerseits mehrere Kopien der mitochondrialen DNA enthalten. Dies ermöglicht oft eine DNA-Sequenzierung der mtDNA aus Fossilien, die nicht zu alt sind (weniger als 100.000 Jahre). Bis 2006 wurden Teile der mtDNA von zwölf Neandertalern aus Deutschland, Kroatien, Russland, Frankreich, Belgien, Italien und Spanien sequenziert.[5] Das Ergebnis:
- Die mtDNA eines Neandertalers unterscheidet sich etwa drei mal soviel von der eines modernen Menschen wie die mtDNA von zwei modernen Menschen sich untereinander unterscheiden.
- Die mtDNA der Neandertaler unterscheiden sich untereinander nur wenig. Die mtDNA moderner Menschen unterscheiden sich untereinander wesentlich stärker.
- Die mtDNA der Neandertaler zeigt keine größere Ähnlichkeit zu der mtDNA heutiger Europäer als zu jener heutiger Afrikaner oder Asiaten.
Die mtDNA des Neandertalers hat sich also relativ früh von der Linie, die zum modernen Menschen geführt hat, abgespalten. Man schätzt, dass die gemeinsame Mitochondriale Eva vom modernem Menschen und Neandertaler vor 550.000 bis 690.000 Jahren gelebt hat, also deutlich früher als die Mitochondriale Eva des modernen Menschen (Krings et al., 1997).
Laut Nordborg (1998) und anderen kann man trotz dieser Daten nicht ausschließen, dass es eine Vermischung zwischen Neandertalern und modernen Menschen gab. Currat & Excoffier (2004) meinen hingegen, dass man anhand dieser mtDNA-Daten eine Vermischung praktisch ausschließen kann.
Die mtDNA aus fossilen Überresten aller moderner Menschen (Homo sapiens), welche bis jetzt untersucht wurden, sind hingegen eng mit den heute lebender Menschen verwandt (Serre et al., 2004). Die mtDNA von Ötzi zum Beispiel gehörte zur Haplogruppe K1 (Rollo et al., 2006).
Genetische Diversität der mtDNA
Die Diversität der mitochondrialen DNA-Sequenzen ist in Afrika am größten. Insgesamt ist die genetische Diversität beim Menschen, im Vergleich mit den anderen Menschenaffen, gering. In einem bestimmten Segment der mtDNA zeigen Schimpansen die drei- bis vierfache genetische Diversität im Vergleich zu Menschen. Allein die mtDNAs von 19 untersuchten Schimpansen aus dem Taï Nationalpark (Elfenbeinküste) zeigen eine höhere Diversität als die aller Menschen, obwohl sie zu einer kleinen, genetisch durchmischten Gruppe gehören (Gagneux et al., 1999).
Bonobos, Gorillas und Orang-Utans zeigen ebenfalls eine höhere genetische Diversität der mtDNA als der Mensch (Jobling et al., 2004).
Genetische Diversität von Menschenaffen im Vergleich zum Menschen Locus Schimpanse vs. Mensch Bonobo vs. Mensch Gorilla vs. Mensch Orang Utan vs. Mensch Referenz Anmerkung mtDNA 3–4 mal so hoch höher höher höher Gagneux et al. (1999) – Y-Chromosom höher höher geringer höher Stone et al. (2002) siehe Adam X-Chromosom 3 mal so hoch keine Daten 2 mal so hoch 3,5 mal so hoch Kaessmann et al. (2001) – Genetische Diversität des Menschen in Afrika, Asien und Europa Locus Afrika Asien Europa Referenz Anmerkung mtDNA (paarweise Unterschiede) 2,08 1,75 1,08 Vigilant et al. (1991) – Y-Chromosom (43 Marker) 0,841 0,904 0,852 Hammer et al. (2001) siehe Adam X-Chromosom (Nukleotid Diversität) 0,035 0,025 0,034 Kaessmann et al. (1999) – Autosomen (Nukleotid Diversität) 0,115 0,061 0,064 Yu et al. (2002) – Eva und die Out-of-Africa-Theorie
- Hauptartikel: Ausbreitung des Menschen
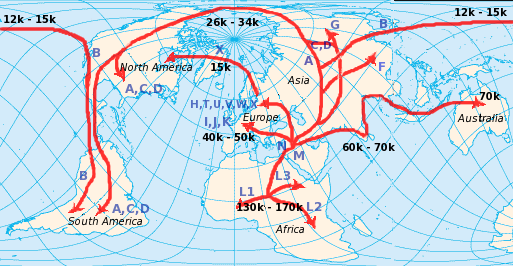
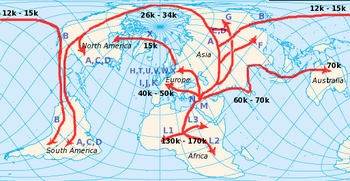 Verbreitung der mtDNA auf der Welt
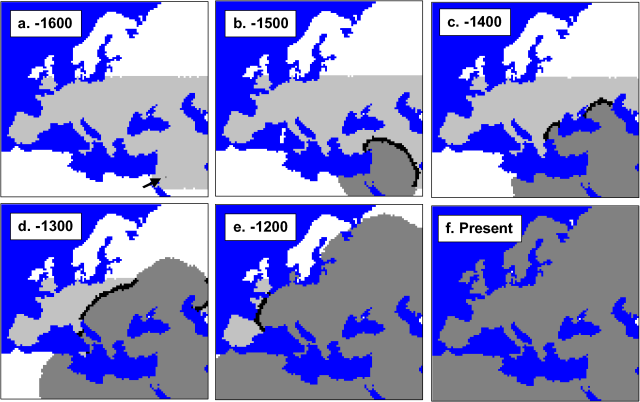
Verbreitung der mtDNA auf der Welt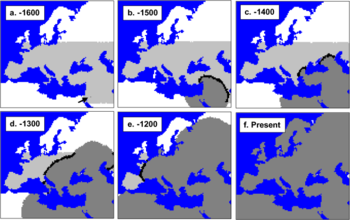 Currat & Excoffier (2004) versuchten die Besiedlung Europas durch moderne Menschen und eine mögliche Vermischung mit Neandertalern zu simulieren. Die Zahlen geben die Zeit in Generationen vor heute wieder. Die schwarzen Streifen sind die Zonen, die von modernen Menschen und Neandertalern besiedelt waren. Die Autoren kamen durch diese Simulationen zu dem Schluss, dass es so gut wie keine Vermischung mit dem Neandertaler gab.
Currat & Excoffier (2004) versuchten die Besiedlung Europas durch moderne Menschen und eine mögliche Vermischung mit Neandertalern zu simulieren. Die Zahlen geben die Zeit in Generationen vor heute wieder. Die schwarzen Streifen sind die Zonen, die von modernen Menschen und Neandertalern besiedelt waren. Die Autoren kamen durch diese Simulationen zu dem Schluss, dass es so gut wie keine Vermischung mit dem Neandertaler gab.Die Theorie der Mitochondrialen Eva ist mittlerweile anerkannt und etabliert, aber was ihre Aussagen über die Fragestellung von Out-of-Africa-Theorie versus „multiregional evolution model“ betrifft, besteht immer noch kein wissenschaftlicher Konsensus. Klar ist, dass die Theorie der Mitochondrialen Eva nicht im Widerspruch zur Out-of-Africa-Theorie steht. Aber sie widerlegt auch nicht zwangsläufig andere Modelle. Schließlich macht sie nur Aussagen über einen einzigen Locus. Die Loci im Zellkern können andere, dank Rekombination wesentlich kompliziertere, Geschichten haben.
Zum Beispiel argumentierte Nordborg (1998), dass das vollständige Fehlen von Neandertaler-mtDNA bei heutigen Europäern durchaus das Ergebnis von Gendrift sein könnte.
Nordborg (1998) nahm in einem einfachen Modell an, dass der moderne Mensch sich zunächst über Europa ausbreitete ohne sich mit Neandertalern zu mischen. Erst nachdem die Expansion abgeschlossen war, mischten sich moderne Menschen mit Neandertalern. Nach diesem Modell könnten die Neandertaler bis zu 25 % Beitrag am Genpool der daraus entstanden Mischpopulation gehabt haben. Durch Gendrift starben die mitochondrialen Erblinien der Neandertaler in der Zwischenzeit aus (siehe oben unter Theorie). Heutige Europäer könnten also in ihrem Zellkern durchaus noch DNA mit Neandertaler-Ursprung haben.
Currat & Excoffier (2004) entwickelten ein wesentlich komplizierteres Modell für die Durchmischung von modernem Menschen und Neandertaler. Die Besiedlung von Europa und die Verdrängung der Neandertaler durch moderne Menschen dauerte etwa 12.000 Jahre (~500 Generationen). Wenn man bei den Berechnungen davon ausgeht, dass Expansion und Vermischung gleichzeitig abgelaufen sind, bekommen die Neandertaler einen großen Vorteil, weil sie Europa vorher besiedelt hatten.
Wenn z. B. moderne Menschen, von Afrika kommend, Anatolien besiedeln und sich teilweise mit den Neandertalern dort mischen, findet die mtDNA der Neandertaler Eingang in den Genpool der modernen Menschen. Expandiert nun die Population der modernen Menschen, so expandiert auch die importierte Neandertaler-DNA. Wenn jetzt z. B. der Balkan von den „Misch-Nachkommen“ besiedelt wird und es wieder zu einer Vermischung mit den dort lebenden Neandertalern kommt, steigt der Anteil der Neandertaler-DNA im Genpool noch weiter. Je weiter die Expansionswelle moderner Menschen nach Westen und Norden vordringt, und sich dabei immer wieder mit den dort lebenden Neandertalern mischt, desto mehr reichert sich Neandertaler-DNA im Genpool an.
Nach diesem Modell würde selbst eine kleine Mischquote ausreichen, damit die mitochondriale DNA des Neandertalers die des modernen Menschen vollständig verdrängt (was nicht der Fall war). Currat und Excoffier (2004) ermittelten, dass es in den 12.000 Jahren, in denen moderne Menschen und Neandertaler Europa bewohnten, nur maximal 120 „Mischkinder“ geben konnte. Die Autoren vermuteten aus dieser extrem geringen Zahl, dass moderne Menschen und Neandertaler verschiedene Arten sind, die zu keiner gemeinsamen Fortpflanzung fähig waren.
Auch große Teile Asiens waren bereits von Vertretern der Gattung Homo (Homo erectus) bewohnt, als sie von modernen Menschen aus Afrika besiedelt wurden. Auch hier finden sich keine mitochondrialen Erblinien, die auf eine Vermischung hindeuten würden.
Zusammenfassend kann gesagt werden, dass die Mitochondriale Eva die Out-of-Africa-Theorie durch folgende Fakten stützt:
- Die geringe genetische Diversität der mitochondrialen DNA beim Menschen (Gagneux et al., 1999).
- Das relativ junge Alter der Mitochondrialen Eva von nur ~175.000 Jahren (Ingman et al., 2000).
- Der mitochondriale Stammbaum zeigt tiefe Äste in Afrika aber sternförmige Struktur außerhalb (Ingman et al., 2000).
- Die mitochondriale DNA der Neandertaler ist eindeutig verschieden von der heutiger Menschen (Serre et al., 2004).
- Eine Vermischung von modernen Menschen mit Neandertalern ist, nach heutigem Wissen, unwahrscheinlich (Currat & Excoffier, 2004).
Untersuchungen an anderen Loci auf dem X-Chromosom, Y-Chromosom und Autosomen deuten ebenfalls auf einen jungen, afrikanischen Ursprung des Menschen hin (Takahata et al., 2001).
„Eva“ in der Populärkultur
- Bryan Sykes hat ein Buch namens Die sieben Töchter Evas geschrieben.
- In River Out of Eden beschreibt Richard Dawkins die menschlichen Vorfahren im Kontext eines Flusses aus Genen und zeigt, dass die Mitochondriale Eva eine von vielen gemeinsamen Ahnen ist, die wir auf Grund der verschiedenen genetischen Pfade zurückverfolgen können.
- Der Discovery Channel hat eine Dokumentation namens Die wirkliche Eva ausgestrahlt.
- Der japanische Roman, Horrorfilm und die Videospielserien Parasite Eve benutzen die mitochondriale Eva-Theorie als Basis für eine Fantasiegeschichte über einen Wissenschaftler, der seine Frau mit Hilfe von regenerierten Zellen wiederbelebt, was fatale Folgen hat.
- Greg Egan schrieb eine Kurzgeschichte mit dem Titel Mitochondriale Eva.
- Ronald D. Moore definiert den Charakter der Hera als Mitochondriale Eva in seiner Science-Fiction-Serie Battlestar Galactica. Hera ist in diesem Fall der erste Hybrid aus Menschen und Zylonen.
Siehe auch
Quellenangaben
Einzelnachweise
- ↑ Aquadro, C.F. & Greenberg, B.D. (1983): Human mitochondrial DNA variation and evolution: analysis of nucleotide sequences from seven individuals. In: Genetics. Bd. 103, S. 287–312. PMID 6299878 PDF
- ↑ Johnson, M.J. et al. (1983): Radiation of human mitochondria DNA types analyzed by restriction endonuclease cleavage patterns. In: J. Mol. Evol. Bd. 19, S. 255–271. PMID 6310133 doi:10.1007/BF02099973
- ↑ Cann, R.L. et al. (1987): Mitochondrial DNA and human evolution. In: Nature. Bd. 325, S. 31–36. PMID 3025745 doi:10.1038/325031a0
- ↑ Behar, D.M. et al. (2007): The Genographic Project public participation mitochondrial DNA database. In: PLoS Genet. Bd. 3, S. e104. PMID 17604454 doi:10.1371/journal.pgen.0030104
- ↑ Green, R.E. et al. (2006): Analysis of one million base pairs of Neanderthal DNA. In: Nature. Bd. 444, S. 330–336. PMID 17108958 doi:10.1038/nature05336
weitere Quellen
- Currat, M. & Excoffier, L. (2004), ‚Modern humans did not admix with Neanderthals during their range expansion into Europe.‘, PLoS Biol 2(12), e421. DOI:10.1371/journal.pbio.0020421
- Derenko, M.V.; Grzybowski, T.; Malyarchuk, B.A.; Czarny, J.; Sliwka, D.M. & Zakharov, I.A. (2001), ‚The presence of mitochondrial haplogroup x in Altaians from South Siberia.‘, Am J Hum Genet 69(1), 237-241. PMID 11410843
- Gagneux, P.; Wills, C.; Gerloff, U.; Tautz, D.; Morin, P.A.; Boesch, C.; Fruth, B.; Hohmann, G.; Ryder, O.A. & Woodruff, D.S. (1999), ‚Mitochondrial sequences show diverse evolutionary histories of African hominoids.‘, Proc Natl Acad Sci U S A 96(9), 5077--5082. DOI:10.1073/pnas.96.9.5077
- Hammer, M. F.; Karafet, T. M.; Redd, A. J.; Jarjanazi, H.; Santachiara-Benerecetti, S.; Soodyall, H. & Zegura, S. L. (2001), ‚Hierarchical patterns of global human Y-chromosome diversity.‘, Mol Biol Evol 18(7), 1189—1203. PMID 11420360
- Ingman, M.; Kaessmann, H.; Pääbo, S. & Gyllensten, U. (2000), ‚Mitochondrial genome variation and the origin of modern humans.‘, Nature 408(6813), 708-713. doi:10.1038/35047064
- Jobling, Mark A.; Tyler-Smith, Chris; Hurles, Matthew. Human Evolutionary Genetics. Origins, Peoples and Disease. ISBN 0-8153-4185-7
- Kaessmann, H.; Heissig, F.; von Haeseler, A. & Pääbo, S. (1999), ‚DNA sequence variation in a non-coding region of low recombination on the human X chromosome.‘, Nat Genet 22(1), 78—81. doi:10.1038/8785 10.1038/8785 10.1038/8785
- Kaessmann, H.; Wiebe, V.; Weiss, G. & Pääbo, S. (2001), ‚Great ape DNA sequences reveal a reduced diversity and an expansion in humans.‘, Nat Genet 27(2), 155—156. doi:10.1038/84773
- Krings, M.; Stone, A.; Schmitz, R.W.; Krainitzki, H.; Stoneking, M. & Pääbo, S. (1997), ‚Neandertal DNA sequences and the origin of modern humans.‘, Cell 90(1), 19-30. doi:10.1016/S0092-8674(00)80310-4
- Nordborg, M. (1998), ‚On the probability of Neanderthal ancestry.‘, Am J Hum Genet 63(4), 1237-1240. PMID 9758610
- Rollo, F.; Ermini, L.; Luciani, S.; Marota, I.; Olivieri, C. & Luiselli, D. (2006), ’Fine characterization of the Iceman’s mtDNA haplogroup.', Am J Phys Anthropol 130(4), 557-564. doi:10.1002/ajpa.20384
- Serre, D.; Langaney, A.; Chech, M.; Nicola, M.T.; Paunovic, M.; Mennecier, P.; Hofreiter, M.; Possnert, G. & Pääbo, S. (2004), ‚No evidence of Neandertal mtDNA contribution to early modern humans.‘, PLoS Biol 2(3), E57. doi:10.1371/journal.pbio.0020057
- Stone, A. C.; Griffiths, R. C.; Zegura, S. L. & Hammer, M. F. (2002), ‚High levels of Y-chromosome nucleotide diversity in the genus Pan.‘, Proc Natl Acad Sci U S A 99(1), 43—48. doi:10.1073/pnas.012364999
- Vigilant, L.; Stoneking, M.; Harpending, H.; Hawkes, K. & Wilson, A. C. (1991), ‚African populations and the evolution of human mitochondrial DNA.‘, Science 253(5027), 1503—1507. doi:10.1126/science.1840702
- Takahata, N.; Lee, S.H. & Satta, Y. (2001), ‚Testing multiregionality of modern human origins.‘, Mol Biol Evol 18(2), 172-183. PMID 11158376
- Yu, N.; Chen, F.; Ota, S.; Jorde, L. B.; Pamilo, P.; Patthy, L.; Ramsay, M.; Jenkins, T.; Shyue, S. & Li, W. (2002), ‚Larger genetic differences within africans than between Africans and Eurasians.‘, Genetics 161(1), 269—274. PMID 12019240
Literatur
- Bryan Sykes: Die sieben Töchter Evas, Lübbe-Verlag, Juni 2001, ISBN 3-7857-2060-2
Weblinks



Wikimedia Foundation.