- Ausbreitung des Menschen
-
 Frühe Migration der Menschen: Tausende Jahre vor der Gegenwart
Frühe Migration der Menschen: Tausende Jahre vor der Gegenwart
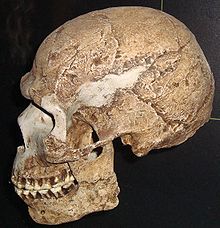 Der Schädel Skhul V, eines der ältesten Fossilien von Homo sapiens außerhalb Afrikas
Der Schädel Skhul V, eines der ältesten Fossilien von Homo sapiens außerhalb AfrikasDie Ausbreitung des Menschen (des Homo sapiens) über die Erde begann den heute vorliegenden wissenschaftlichen Befunden zufolge in Afrika. Zuerst wanderten die Menschen in den Nahen Osten, dann nach Südasien und vermutlich vor etwa 50.000 bis 60.000 Jahren nach Australien. Dabei folgten sie, wie schon in Afrika, dem Verlauf der Küsten. Erst später wurden Zentral- und Ostasien, beide Teile Amerikas und Europa besiedelt. Bis vor wenigen tausend Jahren teilten die modernen Menschen dabei ihren Lebensraum mit weiteren Arten aus der Gattung Homo, in Europa etwa mit den Neandertalern. Seit der Wiederentdeckung Amerikas durch die Europäer 1492 vermischen sich die verschiedenen menschlichen Populationen durch erneute Wanderungsbewegungen in einem Ausmaß, das die genetischen Spuren ihrer Herkunft verwischt.
Diese Rekonstruktion der Ausbreitung des modernen Menschen weist erhebliche Parallelen zur Out-of-Africa-Theorie auf, die sich üblicherweise auf Homo erectus bezieht, aus dem sich in Europa der Neandertaler entwickelte. Die früher verbreitete Ansicht, wonach Homo sapiens sich auf mehreren Kontinenten getrennt voneinander aus Homo erectus entwickelte („Multiregionaler Ursprung des modernen Menschen“), kann heute als widerlegt gelten.
Inhaltsverzeichnis
Stammbäume
 Darwins erste Skizze eines Stammbaums von Tierarten
Darwins erste Skizze eines Stammbaums von TierartenCharles Darwin schrieb 1837 in sein privates Notebook B den Satz „I think“ und zeichnete dann ein System sich gabelnder Linien, um zu erklären, wie er sich die Evolution vorstellte. Seiner epochalen Veröffentlichung über Die Entstehung der Arten von 1859 war ein Evolutionsdiagramm beigefügt, das nun von unten nach oben laufend auch eine zeitliche Abfolge ausdrückte.[1] Statt für Arten können die Linien in solchen Stammbäumen auch für menschliche Populationen stehen.
Paläoanthropologische Befunde
Darwin war es auch, der bereits 1871 in Die Abstammung des Menschen und die geschlechtliche Zuchtwahl vermutete, dass der Mensch aus Afrika stamme. Später waren aber eher „multiregionale“ Modelle beliebt, wonach die verschiedenen menschlichen Rassen getrennt voneinander in verschiedenen Gegenden der Welt entstanden seien.[2] Heute wird diese Ansicht nur noch von Anthropologen in der Volksrepublik China und in Sri Lanka vertreten.
1924 identifizierte der australische Anatom Raymond Dart in Südafrika mit Australopithecus africanus zum ersten Mal einen Vormenschen aus Afrika. In den 1960er- und 1970er-Jahren entdeckten Louis Leakey und andere Mitglieder der Leakey-Familie in Ostafrika dann zahlreiche Exemplare von Homo. Als ältestes Fossil der Gattung gilt heute ein 2,4 Millionen Jahre alter bezahnter Unterkiefer (UR 501) von Homo rudolfensis, den Friedemann Schrenk in Malawi fand. Der früheste Homo erectus (= Homo ergaster) wird auf ein Alter von 1,8 Millionen Jahren datiert. Funde in Dmanisi, Georgien, belegen, dass Vertreter der Gattung Homo nur wenig später bereits Afrika verlassen hatten. Diese Tatsache wird in der Regel als Out-of-Africa-Theorie bezeichnet, aber auch der moderne Mensch hat sich in einer zweiten Auswanderungswelle von Afrika aus über die Welt verbreitet.
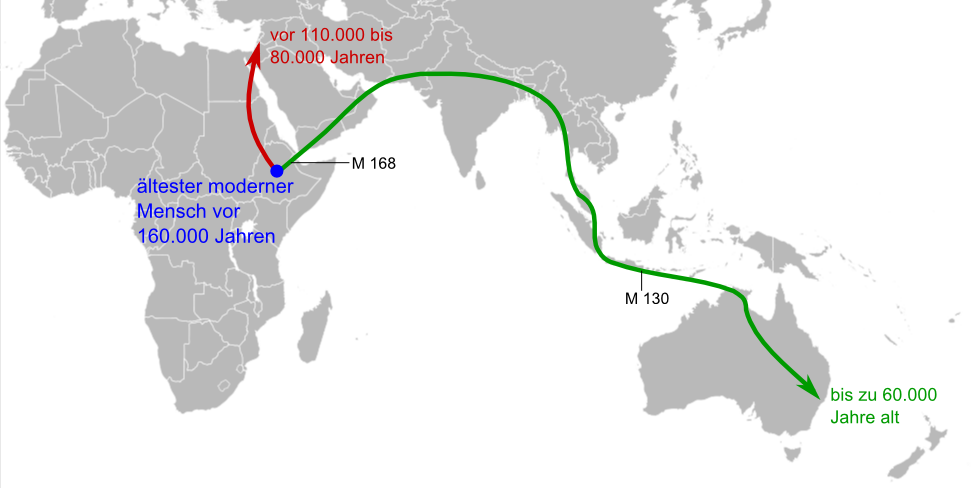
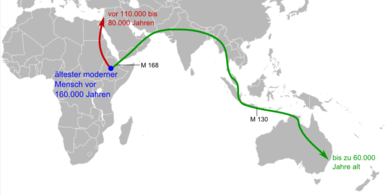 Erste Wanderungen in den Nahen Osten und nach Australien (M 168 und M 130 bezeichnen Marker im Y-Chromosom; siehe unten)
Erste Wanderungen in den Nahen Osten und nach Australien (M 168 und M 130 bezeichnen Marker im Y-Chromosom; siehe unten)Die Frühgeschichte der Art Homo sapiens ist in Afrika – und nur in Afrika – durch eine dichte Reihe von Fossilien repräsentiert. Bereits 1921 wurde in Kabwe (Broken Hill) in Sambia der als Homo rhodesiensis bezeichnete Schädel eines archaischen Homo sapiens gefunden. Fundorte von frühen archaischen Homo sapiens (ca. 500.000–200.000 Jahre alt) liegen außerdem in Saldanha (Südafrika), Ndutu und Eyasi in Tansania, Bodo (Äthiopien) und in Salé (Marokko). Späte archaische Homo sapiens (ca. 200.000–100.000 Jahre alt), die nur noch Überaugenwülste als Reminiszenz an Homo erectus zeigen, stammen aus Florisbad (Südafrika),[3] Omo/Kibish in Äthiopien[4] (Omo 1 und Omo 2), Eliye Springs am Turkana-See in Kenia, Laetoli (Tansania) und Jebel Irhoud (Marokko). Moderne Homo sapiens, die sich in nichts von den heute lebenden Menschen unterscheiden, sind unter anderem in Border Cave und Klasies River Mouth (Südafrika), sowie Omo/Kibish (Äthiopien) nachgewiesen worden. In den Klasies River Mouth-Höhlen zwischen Kapstadt und Port Elizabeth unter Leitung von Hillary Deacon und im Border Cave unter Peter Beaumont wird das erste Erscheinen des modernen Menschen vor 120.000 Jahren angesetzt. Ein fast vollständiger Schädel aus Omo/Kibish ist womöglich sogar 130.000 Jahre alt.[5] Den ältesten modernen Menschen repräsentiert gegenwärtig ein 160.000 Jahre alter Fund aus Herto (Äthiopien); ob auch der Ursprung von Homo sapiens im Nordosten Afrikas lag oder im südlichen Afrika, ist derzeit Gegenstand der wissenschaftlichen Diskussion.[6]
 Schädel von Qafzeh 9
Schädel von Qafzeh 9Die ersten Fossilien des modernen Menschen außerhalb Afrikas sind ab 1931 in Skhul, einer Höhle bei Haifa im Karmel-Gebirge, und kurz danach in Qafzeh nahe Nazareth im heutigen Israel ausgegraben worden. Die ältesten von ihnen lassen sich auf ein Alter von 110.000 Jahren datieren; in der Zeit zwischen 80.000 und 50.000 Jahren versiegen die Nachweise von Menschen in diesen Fundstätten wieder. Das geht mit einem Rückgang der Temperatur von ungefähr 10 °C einher, der sich vor rund 80.000 Jahren im Mittelmeerraum ereignete und die Lebensverhältnisse stark veränderte. Nach diesem ersten Vorstoß aus Afrika starben die Menschen hier entweder einfach aus oder wanderten nach Afrika zurück (und wurden in einigen Fällen durch Neandertaler ersetzt).[7] – Eine mögliche Erklärung bietet die Toba-Katastrophen-Theorie des Anthropologen Stanley Ambrose.[8] [9]
 Mungo Man (oder Mungo 3) aus Australien ist etwa 40.000 Jahre alt.
Mungo Man (oder Mungo 3) aus Australien ist etwa 40.000 Jahre alt.Sieht man von dieser Episode ab, so stammen die ältesten Funde des modernen Menschen außerhalb Afrikas verblüffenderweise aus Australien. Lake Mungo liegt in New South Wales, circa 1000 Kilometer westlich von Sydney. Hier entdeckte Jim Bowler die Überreste eines Menschen namens „Mungo 3“, der inzwischen für 40.000 Jahre alt gehalten wird. Menschliche Artefakte aus einer Sedimentschicht unterhalb dieser Fundstelle werden sogar auf ein Alter von bis zu 60.000 Jahren datiert. Steinwerkzeuge aus einem Fundort im Northern Territory könnten ebenfalls bis zu 60.000 Jahre alt sein.[10] Geht man von diesem Datum aus, benötigte die Menschheit ca. 2000 Generationen seit dem Verlassen Afrikas, um die Erde zu besiedeln. Dabei handelt es sich nicht um zielgerichtete Wanderungen; in den meisten Fällen genügt es anzunehmen, dass eine Gruppe ihren Siedlungsbereich um wenige Kilometer pro Generation erweiterte.
Genetische Stammbäume
Die genetischen Stammbäume geben Auskunft über die prähistorischen Ausbreitungsbewegungen, unter anderem weil die Richtung der Ausbreitung der veränderten Merkmale des Genoms oft noch feststellbar oder nachvollziehbar ist.
Während Skelettmerkmale sich meist graduell voneinander unterscheiden, werden für Stammbäume, die mit genetischen Methoden gewonnen worden sind, so genannte Polymorphismen genutzt, deren Zustand sich sprunghaft ändert. Ein Polymorphismus bezeichnet einen genetischen Unterschied. Auf der Protein-Ebene können das zum Beispiel die vier Blutgruppen des AB0-Systems sein, auf der DNA-Ebene bedeutet zum Beispiel der Polymorphismus M173, dass auf einer bestimmten Position in der Nukleotidabfolge auf dem Y-Chromosom statt eines A ein C steht. Für die Konstruktion von Stammbäumen brauchbare Polymorphismen werden auch Marker genannt.
Bei der zeitlichen Einordnung der Verzweigungen in genetischen Stammbäumen muss bedacht werden, dass die Angaben mit einer großen Ungenauigkeit behaftet sind, weil die Mutationsrate in prähistorischen Populationen schwer zu bestimmen ist. Außerdem gilt für Stammbäume, dass ausgestorbene Zweige keine genetischen Spuren in den lebenden Populationen hinterlassen haben, also allein anhand der Erbinformation rezenter Populationen nicht erkannt werden können.
Stammbäume aus der Verteilung von Blutgruppen
Während des Ersten Weltkriegs untersuchte das polnische Ehepaar Hirszfeld die Verteilung der Blutgruppen bei Soldaten verschiedener Nationalitäten an der Balkanfront. Aus ihren Ergebnissen formulierten sie 1919 die These, dass die Menschheit aus zwei „reinen“ Populationen mit den Blutgruppen A und B entstanden sei, die sich später durch Wanderungen vermischt hätten. Den Ursprung der Blutgruppe A vermuteten sie in Nordeuropa, von B in Indien. Die Menschheit schien demnach zwei getrennte Ursprünge zu haben.[11]
Ab den 1930er untersuchte der Brite Arthur Mourant Tausende von Blutproben aus Hunderten menschlicher Populationen aus aller Welt auf die Verteilung der AB0-Blutgruppen. 1954 fasste er die Ergebnisse in dem Werk The Distribution of the Human Blood Groups [12] zusammen, das in den nächsten 20 Jahren das Standardlehrbuch der menschlichen Populationsgenetik bildete und die moderne Humangenetik begründete.
Richard Lewontin, damals an der Universität von Chicago tätig und eigentlich an Fruchtfliegen interessiert, analysierte 1972 auf dieser Grundlage, wie sich verschiedene menschliche Populationen in ihrer genetischen Variation unterschieden.[13] Das Ergebnis, das ihn selbst überraschte, war, dass rund 85 Prozent der genetischen Variation innerhalb einer Population auftrat. Lediglich sieben Prozent der genetischen Variation unterschieden Populationen voneinander, etwa Finnen von Portugiesen. Und nur die restlichen acht Prozent unterschieden menschliche „Rassen“ voneinander. Lewontin begriff darunter etwa „Kaukasier“ aus dem westlichen Eurasien, Schwarzafrikaner aus dem Gebiet südlich der Sahara oder australische Ureinwohner. Der Forscher veranschaulichte sein Ergebnis durch ein Gedankenexperiment: Wenn durch irgendein schreckliches Ereignis die gesamte Menschheit bis auf die Kikuyu ausstürbe, dann blieben immer noch 85 Prozent der gesamten genetischen Variation unter den Menschen erhalten (das ist keine Besonderheit der Kikuyu, sondern gilt auch für die meisten anderen Populationen). Lewontins Ergebnis ist seitdem in zahlreichen Studien bestätigt worden. Für die Konstruktion genetischer Stammbäume heißt das, dass nur relativ wenige Polymorphismen als Marker geeignet sind.
Eine andere wichtige Einsicht steuerte der Japaner Motoo Kimura bei, der in den USA arbeitete. In den 1950er Jahren waren die meisten Biologen der Ansicht, dass die Selektion die wichtigste Ursache für die Ausbreitung genetischer Abweichungen sei. Kimura berechnete dagegen, dass die meisten Unterschiede durch genetische Drift entstehen, sich also rein zufällig ausbreiten. In diesem Fall liefert die Mutationsrate auch ein Maß für den Zeitpunkt, zu dem ein Marker entstanden ist. Der Populationsgenetiker Luigi Luca Cavalli-Sforza stellte außerdem 1964 das Prinzip der sparsamsten Erklärung auf (engl. parsimony, vgl. „Ockhams Rasiermesser“), das seitdem in sämtlichen Studien zu diesem Thema befolgt wird. Demnach wird ein Muster von Polymorphismen durch die geringstmögliche Zahl an Änderungen erklärt, die nötig sind, um es zu erreichen. Die menschlichen Populationen, die genetisch am wenigsten voneinander abweichen, gelten also als am stärksten miteinander verwandt. Auf dieser Grundlage untersuchten Cavalli-Sforza und Anthony Edwards die Blutgruppenverteilung in 15 menschlichen Populationen. Während Populationen aus Europa und Ostasien hier nahe verwandt erschienen, wirkten sie mit Afrikanern vergleichsweise wenig verwandt.[14] Cavalli-Sforza präsentierte gemeinsam mit Walter Bodmer auch die ersten Daten, wann sich die menschlichen Populationen voneinander getrennt haben sollen: "Afrikaner" und "Ostasiaten" vor 41.000 Jahren, "Afrikaner" und "Europäer" vor 33.000 Jahren und "Europäer" und "Ostasiaten" vor 21.000 Jahren.[15]
Stammbäume aus Mitochondrien-DNA
Seitdem Walter Gilbert und Fred Sanger unabhängig voneinander Methoden zur Sequenzierung von DNA entwickelt hatten (Nobelpreis 1977), wurde die Konstruktion von Stammbäumen auf dieser Ebene möglich. Das vervielfacht die Anzahl der verfügbaren Polymorphismen: Statt den vier Blutgruppen des AB0-Systems konnte nun potenziell an jedem einzelnen Nukleotid der menschlichen DNA ein Polymorphismus in Form einer Punktmutation nachgewiesen werden. Die menschliche DNA umfasst drei Milliarden Nukleotide.
In den meisten Chromosomen wird das von Mutter und Vater stammende Erbgut allerdings ständig rekombiniert, sodass das ursprüngliche Muster der Polymorphismen nach einigen hundert Generationen verloren geht. Der australische Biochemiker Allan Wilson von der Universität von Kalifornien in Berkeley setzte deswegen aus folgenden Gründen seine Doktorandin Rebecca Cann auf die DNA von Mitochondrien an:
- Mitochondrien werden nur von Müttern vererbt und besitzen ihr eigenes Erbgut außerhalb des Zellkerns;
- die Sequenz dieser mitochondrialen DNA (mtDNA) liegt nur in einer einzigen Version vor;
- die Mutationsrate ist verglichen mit Chromosomen zehnmal so hoch, sodass etwa alle 100 Nukleotide mit einem Polymorphismus zu rechnen ist;
- außerdem enthält eine einzige Körperzelle einige hundert Mitochondrien, sodass für die anfangs noch unempfindlichen Methoden ausreichend DNA zur Verfügung stand.
In der Arbeit, die Cann 1987 veröffentlichte, identifizierte sie erstmals eine gemeinsame Urahnin:[16] „Alle diese mitochondrialen DNAs stammen von einer einzigen Frau ab, die vermutlich vor rund 200.000 Jahren wahrscheinlich in Afrika lebte.“[17] In der Tagespresse wurde diese Urahnin schnell als „Eva“ bezeichnet. Das bedeutet aber nicht, dass in dieser Population nur eine Frau gelebt hätte; die Mitochondrien aller anderen damals lebenden Frauen sind nur im Laufe der Zeit ausgestorben, da es keine rein weibliche Ahnenkette zu ihnen zurück gibt.
Schwarzafrikaner zeigten die größte genetische Variabilität in ihren mtDNA-Sequenzen – bei einer als konstant angenommenen Mutationsrate hatten sie also die längste Zeit gehabt, um zu evolvieren. Mit anderen Worten: Der Ursprung von Homo sapiens in Afrika ließ sich auch mit genetischen Methoden bestätigen. Spätere Studien haben dieses Ergebnis bekräftigt, wenn auch der Zeitpunkt der ersten Aufspaltung im Stammbaum auf vor 150.000 Jahren korrigiert wurde.[18]
Mit einer weiteren Methode kann der Zeitpunkt kalkuliert werden, ab dem Populationen begonnen haben, sich exponentiell zu vermehren. Polymorphismen reichern sich mit einer konstanten Rate an und bilden für eine Population eine glockenförmige Verteilung. Der Anthropologe Henry Harpending von der Pennsylvania State University untersuchte so genannte mismatches in der mtDNA von 25 menschlichen Populationen und fand in 23 von ihnen die typischen Zeichen für ein kürzlich durchlaufenes exponentielles Wachstum. Die Bevölkerungsvermehrung setzte demnach in Afrika vor 60.000 Jahren, in Asien vor 50.000 und in Europa vor 30.000 Jahren ein.[19]
Stammbäume aus Y-chromosomaler DNA
Ein weiterer Abschnitt in der menschlichen DNA kommt ebenfalls nur in einer Version vor: das Y-Chromosom. Es wird ausschließlich von Vätern an Söhne weitergegeben. Zwar kommt auf dem Y-Chromosom nur etwa alle 1000 Nukleotide ein Polymorphismus vor, aber da es etwa 50 Millionen Nukleotide lang ist – im Gegensatz zur mitochondrialen DNA mit lediglich rund 16.000 Nukleotiden – hat es sich inzwischen als der bei weitem geeignetste Ansatzpunkt für die Konstruktion genetischer Stammbäume erwiesen.
Die Ergebnisse bestätigen die mit mitochondrialer DNA gewonnenen Stammbäume: Auch die anhand Y-chromosomaler DNA konstruierten Stammbäume wurzeln in Afrika. Das Alter des ersten gemeinsamen männlichen Vorfahren beträgt hier allerdings lediglich 59.000 Jahre.[20] Das ist kein Widerspruch zum mtDNA-Stammbaum, denn die Tiefe eines Stammbaums hängt von der Anzahl der verfügbaren Polymorphismen ab. Da das Y-Chromosom aber nur relativ wenige Polymorphismen aufweist, läuft der damit gewonnene Stammbaum früher zusammen und liefert keine Informationen mehr über ein Alter von 59.000 Jahren hinaus. Umgekehrt setzt dieses Ergebnis aber eine obere zeitliche Grenze für die Auswanderung aus Afrika: Noch vor 59.000 Jahren hatte demnach noch kein Angehöriger der Art Homo sapiens Afrika verlassen (mit einem Fehler zwischen 40.000 und 140.000 Jahren). Der ökologische Raum „Afrika“ umfasst hier allerdings auch den Nahen Osten, wie die erwähnten Fossilfunde aus Israel belegen. Die weiteren Ergebnisse sind weiter unten in einer Zusammenschau dargestellt.
Stammbäume aus Einzelnukleotid-Polymorphismen
Heute beziehen Forscher das gesamte Erbgut in ihre Analysen ein. An der DNA-Sequenz des menschlichen Genoms ist die Evolution des Homo sapiens und seiner Vorformen mit Methoden der Humangenetik nur in bestimmten Grenzen ablesbar, weil DNA in fossilen Knochen oft zu weit zerfallen ist. Es ist daher nur selten möglich, Aufschluss über die Ausbreitung menschlicher Populationen durch den direkten Vergleich alter und neuer Genome zu bekommen. Allerdings befinden sich seit der Entschlüsselung des Humangenoms andere Analyseverfahren in schneller Entwicklung, die aus Resten des Genoms in fossilen Knochenfunden und aus den Genomen heutiger Populationen auf frühere Selektionsprozesse in einzelnen Populationen rückschließen lassen. Die daraus gewonnenen Erkenntnisse belegen in groben Zügen, aber immer sicherer, wie sich die Menschen ausbreiteten.
Grundlage einiger Verfahren sind die Einzelnukleotid-Polymorphismen (SNPs). Hierbei handelt es sich um Punktmutationen, das heißt um genetische Veränderungen (Allele), die sich zu einem gewissen Grad im Genpool einer Population durchgesetzt haben. Wenn solche Punktmutationen zu unterschiedlichen Zeitpunkten aufgetreten sind und sich danach in der Population verbreitet haben, kann aus ihrer Häufigkeit und ihrer Kombination auf verwandtschaftliche Beziehungen zwischen den aus einer Ursprungspopulation hervorgegangenen, mittlerweile von einander getrennt lebenden Folgepopulationen und – hieraus abgeleitet – auf deren Ausbreitung geschlossen werden.
Ein bestimmter SNP („FY*O“) kommt beispielsweise bei allen Populationen nördlich der Sahara kaum vor, ist jedoch im subsaharischen Afrika weit verbreitet.[21] Daraus schließen die Autoren dieser SNP-Analyse, dass es – ihren Angaben zufolge vor 75.000 bis 50.000 Jahren – zu einer langfristigen Trennung der nordafrikanisch-kleinasiatisch-eurasischen Populationen von den subsaharischen Populationen gekommen sein muss, was sie als Bestätigung der Out-of-Afrika-Theorie bewerten.
Eine weitere Punktmutation unterscheidet die heutigen Populationen Ostasiens sowie die Urweinwohner von Melanesien, Papua-Neuguinea und Amerika von den Populationen Europas, des Nahen Ostens, aus Zentralasien und Südasien: Diese Verteilung spiegelt vermutlich die Trennung der Menschen in eine ostasiatische und eine westeurasische Population wider, deren genauer Zeitpunkt unbekannt ist, die sich aber vor rund 30.000 bis 20.000 Jahren zugetragen haben dürfte. Studien der Polymorphismen an einzelnen Genen, etwa dem Beta-Globin-Gen, dem CD4-Gen und einer Region auf dem Chromosom 21, zeigen für afrikanische Populationen die bei weitem höchste genetische Variabilität. „Die heutige geografische Verbreitung von Allelen entspricht weiterhin stark den frühen vorzeitlichen Ausbreitungsbewegungen der Menschen in Europa und Asien. Denn die Muster im Genom, die damals entstanden, haben sich deutlich erhalten. Später kamen nur erstaunlich wenige Feinanpassungen an lokale Gegebenheiten und heutige Umwelten hinzu.”[22]
Nach anderen Verfahren werden Abweichungen in bestimmten Informationseinheiten untersucht, etwa bei Genen (Haplotyp-Varianten), oder die Variationen in der Anzahl von definierten Genomabschnitten, so genannte Kopieanzahl-Variationen (copy number variations, CNVs). Dies kann mit Druckfehlern in einzelnen Buchstaben, mit Veränderungen in einem ganzen Wort oder mit dem Hinzufügen oder Entfernen von ganzen Abschnitten in einem Text verglichen werden.
Linguistische Methoden zur Konstruktion von Stammbäumen
Die Entwicklung der Sprache muss bei der Ausbreitung des modernen Menschen eine wesentliche Rolle gespielt haben. Die Linguistik kann zwar nur wenige Aussagen über sehr frühe sprachliche Entwicklungen während der Ausbreitung der Menschen machen; wohl aber können die Beziehungen zwischen der genetischen und der linguistischen Evolution untersucht werden, was ebenfalls zu einem genaueren Verständnis der Ausbreitung der Menschen beitragen kann.[23] So fand sich in einer Analyse von 504 Sprachen aus aller Welt die größte Vielfalt an Phonemen in Afrika; in Südamerika und Ozeanien ist sie am geringsten. Der Autor erklärt dies durch eine Reihe von Gründereffekten: Bei der Wanderung über die Welt wurden neue menschliche Populationen meist nur von wenigen Individuen gegründet. Bei diesem Prozess gingen von den in Afrika üblichen Phonemen immer mehr verloren. Innerhalb von Afrika liegt nach diesen Ergebnissen der Ursprung der ersten menschlichen Sprache, oder Sprachen, in West-, Zentral- oder dem südlichen Afrika.[24]
Die Ausbreitung der Menschen über die Erde
Ursprünge
Innerhalb von Afrika war der moderne Mensch spätestens vor 60.000 und nach dem Grabungsbefund in Herto bereits vor 160.000 Jahren vorhanden. Die größte genetische Vielfalt und damit die ältesten Linien der Menschheit finden sich südlich der Sahara. Vor allem die San – früher als Buschleute bekannt – zeigen die Art von genetischer Vielfalt, die für die frühesten Menschen charakteristisch ist.[25] Auch die mtDNA-Stammbäume und die eigentümlichen Klicksprachen der San legen eine Verbindung zu den frühesten menschlichen Vorfahren nahe. Aus Grabungen ist bekannt, dass das Verbreitungsgebiet der San früher bis nach Äthiopien und Somalia reichte. Insgesamt gilt für alle Menschen: Je weiter der Abstand zu Äthiopien ist, desto geringer wird die genetische Vielfalt.[26]
Die Besiedlung Australiens über Südasien
Ein universell vorhandenes Ausbreitungsmuster ist darin zu erkennen, dass der Mensch dem Verlauf der Küsten folgte (noch heute siedelt der größte Teil der Menschheit in Meeresnähe).[27] Abfallhaufen von Muschelschalen, die auch Steinwerkzeuge enthielten, sind an der Höhle von Pinnacle Point am Indischen Ozean in Südafrika auf ein Alter von 164.000 Jahren,[28] an der Küste von Eritrea auf ein Alter von 125.000 Jahren datiert worden.[29] Asien muss nicht unbedingt auf dem Landweg über die Sinai-Halbinsel und den „levantinischen Korridor“ besiedelt worden sein. Verschiedene Wissenschaftler schlagen vielmehr einen Weg über den Bab al-Mandab, die arabische Halbinsel und die Straße von Hormus vor. Gestützt wird diese Version durch den Fund von Steinwerkzeugen, die auf 70.000 bis 80.000 Jahre datiert worden sind, in Shi'bat Dihya 1 (Jemen) durch ein französisches Team, sowie von 125.000 Jahre alten Werkzeugen durch ein internationales Team in Jebel Faya 1 (auch: FAY-NE1, Vereinigte Arabische Emirate).[30][31][32] Jedoch fehlen hier bisher Fossilien der Werkzeugmacher.
Die frühen Funde des Java-Menschen (Homo erectus) lassen vermuten, dass es Ausbreitungsrouten entlang der asiatischen Südküste gegeben hat. Fossilien erhalten sich allerdings unter Küstenbedingungen besonders schlecht, vor allem aber liegt die Küstenlinie der Kaltzeiten heute unter Wasser, so dass mögliche Fundorte aus dem überwiegenden Teil des Pleistozäns heute im Indischen Ozean liegen. Steinwerkzeuge, die dem frühen Homo sapiens zugeschrieben werden, werden in Indien als Middle Palaeolithic bezeichnet (abweichend zum europäischen Mittelpaläolithikum der Neandertaler) und sind dort sehr selten. Vermeintliche Steinwerkzeuge aus der Fundstelle Jwalapuram im südindischen Bundesstaat Andhra Pradesh wurden auf 74.000 Jahre vor heute datiert.[33] Die Funde sind umstritten, zumal es keine zugehörigen Fossilien gibt.[30] Auch weitere Fundorte Indiens, wie die Billasurgam-Höhlen in Kurnool, Andhra Pradesh, werden unmittelbar vor die Eruption des Toba datiert und führen einige Wissenschaftler zur Toba-Katastrophen-Theorie, die vor etwa 70.000 Jahren zu einer starken Dezimierung der Bevölkerung Südasiens geführt habe.[34] Dem wird entgegengehalten, dass die benachbarten Flores-Menschen den Ausbruch jedoch überlebt haben, das Klimaereignis demnach überinterpretiert werde. Zudem gibt es aus dem Jungpaläolithikum ab etwa 40.000 Jahren vor heute eine Reihe von Fundplätzen, an denen Klingen hergestellt worden sein müssen, was nur mit dem Homo sapiens in Verbindung gebracht werden kann, da diese Fertigkeit als maßgebliche technische Veränderung hervortritt und vergleichbaren Weiterentwicklungen im europäischen Jungpaläolithikum ähnlich ist.[35] In diese Zeit werden auch die ältesten gesicherten Fossilbelege Australiens datiert (vgl. Mungo Man, Mungo Lady). Neue und mit genaueren Methoden [36]unternommene Untersuchungen des zuletzt gefundenen Mongo 3 datieren ihn auf 57.000 bis 69.000 Jahre.
 Australische Aborigines
Australische AboriginesWährend der Grabungsbefund also dürftig ist, belegen genetische Analysen die Wanderung entlang der südasiatischen Küste. Der erste Marker im Y-chromosomalen Stammbaum, der Nicht-Afrikaner von den meisten Afrikanern unterscheidet, heißt M168 und muss im Zeitraum zwischen 31.000 und 79.000 Jahren vor unserer Zeit entstanden sein. Er deckt sich mit dem Marker L3 im mitochondrialen Stammbaum vor 50–60.000 Jahren. Die Abkömmlinge von M168 und L3 leben im heutigen Äthiopien und Sudan sowie in der gesamten Welt außerhalb Afrikas. Unmittelbar nach der Verzweigung L3 im Stammbaum der mitochondrialen DNA folgen zwei weitere Marker, die den Cluster M definieren. 20 Prozent der Menschen in Indien und nahezu 100 Prozent der australischen Ureinwohner tragen diesen Cluster M.[37] Im Y-chromosomalen Stammbaum folgt auf M168 der Marker M130. Er ist westlich des Kaspischen Meers praktisch nicht vorhanden, kommt in Indien in fünf Prozent aller Männer vor, in Malaysia zu zehn Prozent, zu 15 Prozent auf Neu-Guinea und zu 60 Prozent unter australischen Ureinwohnern. Die mit Mitochondrien wie durch Y-Chromosomen gewonnenen Stammbäume bestätigen also eine Wanderungsbewegung durch Südasien in Richtung Australien. So genannte „Negritos“, die in isolierten Gegenden Südasiens – etwa auf den Andamanen-Inseln – leben, könnten die unmittelbaren Nachfahren dieser ersten Migranten sein.
Die Besiedlung Australiens ist besonders bemerkenswert, da der Kontinent von Asien aus gesehen auch während der Eiszeit hinter dem Horizont lag. Spätestens vor rund 50–60.000 Jahren müssen also Menschen fernab sichtbarer Küsten die hohe See befahren haben. Die Rekonstruktion der Erstbesiedelung Australiens und Asiens wird auch durch eine Untersuchung der Varianten des Magenbakteriums Helicobacter pylori unterstützt. Demnach trugen die Menschen der ersten Wanderungswelle eine Variante in ihren Mägen, die noch heute bei den Ureinwohnern Australiens und der Hochebene Neuguineas nachweisbar ist. Eine zweite, abweichende Variante weisen hingegen die Ureinwohner Taiwans, Melanesiens und Polynesiens auf.[38]
Die Besiedlung Asiens in einer zweiten Siedlungswelle
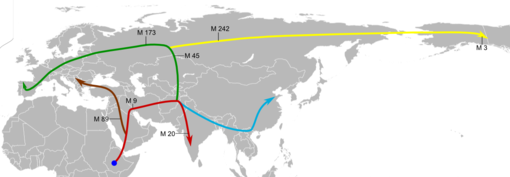 Die zweite Siedlungswelle ab etwa 40.000 Jahren vor heute (M bezeichnet Marker auf dem Y-Chromosom; siehe Text)
Die zweite Siedlungswelle ab etwa 40.000 Jahren vor heute (M bezeichnet Marker auf dem Y-Chromosom; siehe Text)Der Marker M9 ist vor rund 40.000 Jahren in der Gegend des heutigen Iran oder in Südzentralasien entstanden. Er wird von fast sämtlichen Männern getragen, deren Vorfahren Eurasien, Nord- und Südamerika besiedelt haben. Diese Eurasier dehnten ihren Lebensraum zunächst nach Osten aus, wobei sie auf das Gebirgssystem des Hindukusch, Himalaya und Tianshan stießen. Eine Gruppe orientierte sich nach Süden und besiedelte den indischen Subkontinent. Sie wird durch den zusätzlichen Marker M20 charakterisiert, der in Südindien bei etwa der Hälfte aller Männer auftritt, außerhalb Indiens dagegen nur noch zu ein bis zwei Prozent im Nahen Osten. Die Besiedlung Südindiens durch Männer mit dem Marker M20 muss vor etwa 30.000 Jahren abgelaufen sein. Hier liegt einer der wenigen Fälle vor, in denen der Y-chromosomale und der mitochondriale Stammbaum voneinander abweichen, denn Männer mit dem Marker M130 aus der ersten Besiedlungswelle sind kaum noch nachweisbar, während der mitochondriale Cluster M (siehe oben) gut repräsentiert ist. Dies wird dahingehend interpretiert, dass die männliche Bevölkerung aus der ersten Besiedlungswelle getötet wurde oder sich jedenfalls nicht mehr fortpflanzen konnte, während die Einwanderer sich die Frauen nahmen.[20]
Für Indien ist noch eine besondere Gruppe von Mutationen, die Haplogruppe M2 innerhalb des mitochondrialen Clusters M, interessant. Sie findet sich mit einem Anteil von etwa 10 Prozent in sämtlichen 24 indischen Stämmen, die in einer großen Studie unter Satish Kumar analysiert wurden. Diese Haplogruppe M2 hält sich nicht an die Sprachgrenze zwischen den dravidischen und austroasiatischen Sprachen. In diesem Fall hätte also eine Rekonstruktion nach linguistischen Kriterien zu falschen Abstammungsverhältnissen geführt. Die M2-Haplogruppe lässt sich, jedoch weniger häufig, auch unter den höheren Kasten der Brahmanen und Kshatriyas nachweisen. Einerseits bestätigen die Ergebnisse damit die Hypothese, dass die Angehörigen höherer Kasten auf eine Einwanderung vor etwa 10.000 Jahren zurückgehen könnten, andererseits belegen sie aber auch, dass es durchaus zu Vermischungen zwischen den verschiedenen Kasten kam.[39]
Wie bedeutend die Route durch die Steppen nördlich des Hindukusch war, ist neuerdings wieder umstritten (Marker M45 vor 35.000 Jahren in Zentralasien entstanden). Steinwerkzeuge in den Altai-Bergen sind mit rund 40.000 Jahren (Jungpaläolithikum) der bisher frühestdatierte Beleg für Menschen in diesem Raum. Die bisher größte Studie zur Genetik der asiatischen Völker des „HUGO Pan-Asian SNP Consortium“, die auf rund 50.000 SNPs basiert, veranschlagt den Anteil dieser Nordroute für die Besiedlung Ostasiens als nicht sehr hoch, wenn überhaupt vorhanden. Demnach wären China und Zentralasien fast ausschließlich von Südostasien her besiedelt worden. Trotzdem sind die Han-Chinesen genetisch deutlich in eine nördliche und eine südliche Population unterteilt.[40]
Die Besiedlung des Nahen Ostens und Europas
Auf der M168-Linie tritt auch der Marker M89 auf. Er koexistiert nie mit M130, muss also entstanden sein, nachdem die erste Gruppe Menschen bereits Afrika in Richtung Australien verlassen hatte. M89 charakterisiert Männer aus dem nordöstlichen Afrika und dem Nahen Osten und ist vor rund 40.000 Jahren (plus/minus 10.000 Jahre) entstanden. Das stimmt mit den ersten Funden von Steinwerkzeugen im Nahen Osten überein, die etwa 45.000 Jahre alt sind (abgesehen von den Funden in Qafzeh und Skhul im heutigen Israel, die aus dem folgenlosen, ersten Vorstoß vor 110.000 Jahren stammen, siehe oben). Einige dieser Menschen sind in den Balkanraum weitergewandert. Allerdings tragen nur wenige Prozent der europäischen Männer den M89-Marker, sodass die meisten Europäer aus einer anderen Wanderungsbewegung stammen müssen.
Praktisch alle Europäer stammen von einer Population aus Zentralasien mit dem Marker M173 ab. Heute wird M173 am häufigsten in den am stärksten isolierten europäischen Populationen, etwa in den keltischen Siedlungsgebieten und im Baskenland, gefunden. Weitere Marker (deren Analysemethode hier nicht besprochen worden ist: Es handelt sich um Mutationen in Mikrosatelliten-DNA) datieren die Entstehung von M173 auf ein Alter von 30.000 Jahren. In jener Phase der Eiszeit hatten sich die vorher nur zentralasiatischen Steppen bis ins heutige Frankreich ausgedehnt. Die ersten modernen Europäer werden häufig nach Cro-Magnon benannt, einem Fundort in Frankreich, wo die ersten Knochen 1868 ausgegraben wurden. Die Cro-Magnon-Menschen waren häufig über 180 cm groß und hatten einen feingliedrigen Körperbau. Der älteste Schädel eines modernen Menschen in Europa stammt aus der rumänischen Höhle Peștera cu Oase und wird als „Oase 2“ bezeichnet. Er wurde auf 40.500 Jahre datiert.[41] Zwei Milchzähne aus der Grotta del Cavallo in Apulien wurden sogar auf ein Alter von 45.000 bis 43.000 Jahren vor heute datiert.[42] Eine direkte Datierung von Knochenfunden aus der Fundstelle Buran-Kaya III auf der Halbinsel Krim (Ukraine) ergab ein Alter von 31.900 +240/−220 BP.[43] Die nächst jüngeren Fossilien aus Mladeč in Tschechien, der englischen Kent's Cavern sowie einigen französischen Fundstellen sind allesamt 32.000 bis 30.000 Jahre alt.[44] Die dazu passende Kultur ist das Aurignacien mit den ältesten Belegen aus Höhlen der Schwäbischen Alb und der Fundstelle Willendorf II in Niederösterreich (bis zu 40.000 Jahre alt).
Die Besiedlung Amerikas
 Projektilspitze der Clovis-Kultur
Projektilspitze der Clovis-KulturDer älteste Fundort Ostsibiriens (“Yana Rhinoceros Horn Site”) ist bis zu 30.000 Jahre alt[45], andere Fundorte in Dyuktai (südöstlich von Jakutsk) und am Ushki-See auf der Halbinsel Kamtschatka etwa 20.000 Jahre. Eine Abfolge der Marker M45, M242 – der in Sibirien am häufigsten ist – und M3 – der ausschließlich in Amerika vorkommt – definiert eine Wanderungsbewegung von Sibirien nach Amerika (siehe Karte zur zweiten Siedlungswelle). Dabei lässt sich ein Datum früher als vor 20.000 Jahren ausschließen. Der Weg über die Landbrücke Beringia – über die heutige Beringstraße – war allerdings bis 15.000 Jahre vor unserer Zeit durch Eis blockiert. Erst danach kann der moderne Mensch nach Amerika eingewandert sein, falls er den Landweg benutzt hat; tatsächlich haben Ausgrabungen in Texas mit dem Buttermilk Creek Complex die ältesten bisher bekannten Artefakte freigelegt, die in die Zeit zwischen 15.500 und 13.200 Jahren vor heute datiert wurden.[46] Um dieselbe Zeit wanderte auch der Grizzlybär aus Sibirien nach Nordamerika ein. Die genetischen Daten erlauben in diesem Fall auch die Größe der Ursprungspopulation abzuschätzen: Zehn bis zwanzig Individuen würden ausreichen, um die in Amerika anzutreffende genetische Vielfalt zu erklären; es können aber auch wenige hundert Individuen gewesen sein.
Funde in den Paisley-Höhlen, Oregon bestätigen das für die modernen Menschen typische Wanderungsmuster entlang der Küste: Innerhalb von nur rund tausend Jahren legten sie den Weg bis an die Spitze Südamerikas zurück, wie Funde in Monte Verde, Chile belegen. Erst später wurde das Landesinnere besiedelt. Die Clovis-Kultur mit ihren charakteristischen Projektilspitzen war die erste flächenhaft verbreitete Kultur.
Einige Funde[47] mit umstrittener Datierung[48] deuten auf eine wesentlich frühere Besiedlung Amerikas hin. Falls sich die Datierungen bestätigen sollten, wären diese frühen Migranten ausgestorben, denn im Erbgut der rezenten Amerikaner lassen sich keine Spuren einer früheren Besiedlung finden.
Thor Heyerdahl vermutete eine Migration mittels Booten über den Atlantik in der Antike von Nordafrika nach Südamerika; diese Vermutung gilt aber heute als widerlegt.
Etwa 1000 n. Chr. wurde Nordamerika von Wikingern besucht, über etwa 50 Jahre fand eine Migration sehr geringen Ausmaßes statt.
Die restliche Erde
In der DNA der meisten Polynesier fehlen neun Nukleotide (Deletion) und drei weitere Nukleotide sind ausgetauscht (Substitution). Zwei von diesen drei Substitutionen kommen bei den Ureinwohnern Amerikas nicht vor, was - neben vielen anderen Argumenten - gegen die von Thor Heyerdahl vertretene Theorie spricht, dass der pazifische Raum von Amerika her besiedelt worden sei. Tatsächlich sind die pazifischen Inseln von West nach Ost und damit erstaunlicherweise gegen die vorherrschende Windrichtung kolonisiert worden. Die genaue Abfolge ist von Archäologen erarbeitet worden: Bis um 1200 v. Chr. hatten Polynesier die Salomonen östlich von Neuguinea erreicht. Um diese Zeit drang eine Gruppe von Menschen aus dem Bismarck-Archipel bis zu den Fidschi-Inseln, Samoa und Tonga vor. Sie sind archäologisch durch ihre spezielle Keramik im Lapita-Stil gut fassbar. Dann folgte eine Pause von etwa 1500 Jahren Dauer. Zwischen 600 und 800 n. Chr. wurden die am besten zugänglichen Inseln von Ost-Polynesien besiedelt: die Cook-, Gesellschafts- und Marquesas-Inseln. Erst 1200 n. Chr. wurde Neuseeland kolonisiert, womit dann sämtliche geeignete Inseln bis nach Hawaii und der Oster-Insel von Menschen bewohnt waren.[49]
Aus historischen Quellen ist bekannt, dass das Festland der Antarktis erstmals 1820 von einer russischen Expedition unter Fabian Gottlieb von Bellingshausen gesichtet wurde, im folgenden Jahr landeten dort Robbenfänger. Ab 1899 überwinterten Menschen auf dem antarktischen Festland.[50]
 Der Profigolfspieler Tiger Woods ist afrikanischer, indianischer, chinesischer, thailändischer und europäischer Abstammung
Der Profigolfspieler Tiger Woods ist afrikanischer, indianischer, chinesischer, thailändischer und europäischer AbstammungDer bislang detaillierteste Stammbaum ist 2008 von der Arbeitsgruppe von Cavalli-Sforza vorgelegt worden.[51] Die bisher dargestellten Ergebnisse beziehen sich allerdings immer auf die jeweiligen Urbevölkerungen. Nach der Entdeckung Amerikas durch Kolumbus 1492 verdrängten Menschen europäischer Herkunft in Nord- und Südamerika sowie in Australien die Ureinwohner aus weiten Gebieten bzw. bildeten Mischbevölkerungen. Die Urbevölkerung war u.a. gegen die Krankheiten, die die Eindringlinge aus Europa mitbrachten, schutzlos.[52] Heute gibt es praktisch keine vollkommen isoliert lebenden menschlichen Gemeinschaften mehr. Die Vermischung von Menschen aus aller Welt führt dazu, dass Analysen der hier beschriebenen Art bald nicht mehr möglich sein werden.
Begegnung mit anderen Arten der Gattung Homo
Die Menschheit befindet sich heute in einer außergewöhnlichen Situation, da sie inzwischen die einzige Art der Gattung Homo repräsentiert. In Eurasien war das Gebiet, das von Homo sapiens besiedelt wurde, bereits von Homo erectus bewohnt gewesen. Im Nahen Osten und in Europa sind Homo sapiens und der „Neandertaler“, nach derzeitiger Interpretation eine eigene Art, sogar noch aufeinander getroffen. Analysen von Svante Pääbo, damals Universität München, an mitochondrialer DNA des Typus-Exemplars des Neandertalers haben gezeigt, dass diese mtDNA stark von allen Sequenzen abweicht, die von Homo sapiens bekannt sind.[53] Jedoch ergab eine 2010 veröffentlichte Studie derselben Forschergruppe, dass das Genom aus dem Zellkern der Neandertaler eine signifikant größere Ähnlichkeit mit dem Zellkern-Genom von Europäern und Asiaten hat als mit dem von Afrikanern. Die Studie kam zu dem Ergebnis „dass der Genfluss vom Neandertaler zu den Vorfahren der Nicht-Afrikaner erfolgte, bevor sich die eurasischen Gruppen voneinander trennten“. Das entsprechende Siedlungsgebiet, in dem Neandertaler und anatomisch moderner Mensch in der Zeitspanne von vor 110.000 Jahren bis vor rund 50.000 Jahren koexistierten, war der Nahe Osten.[54]
 Schädel des Homo floresiensis
Schädel des Homo floresiensisDer moderne Mensch muss den Neandertaler nicht aktiv ausgerottet haben, zumal die beiden Arten unterschiedliche ökologische Nischen besetzten. Schon ein geringfügiger Unterschied in den Reproduktionsraten würde ausreichen, das Aussterben der Neandertaler zu erklären. In Europa waren die letzten Neandertaler vor etwa 30.000 Jahren auf isolierte Inseln wie in Zafarraya in Spanien zurückgedrängt, vor 25.000 Jahren waren sie auch dort ausgestorben.
In China ist die dem Neandertaler entsprechende Form des Homo erectus, die bis heute noch nicht benannt worden ist, vor rund 100.000 Jahren ausgestorben. Für die Lücke zwischen 100.000 Jahren und dem Eintreffen des modernen Menschen vor rund 40.000 Jahren gibt es jedenfalls keine Funde. Dies wird von manchen chinesischen Wissenschaftlern anders interpretiert; sie leiten den modernen Chinesen unmittelbar vom asiatischen Homo erectus ab. Dafür liefern die genetischen Stammbäume keinen Hinweis, und die Fossilien geben keinen Hinweis auf eine Kreuzung zwischen Homo sapiens und Homo erectus in Asien.
2008 wurde der so genannte Denisova-Mensch aus der Denissowa-Höhle ⊙51.484.68 im Altai-Gebirge geborgen, der belegt, dass es vor rund 40.000 Jahren im Altai neben Homo sapiens und dem Neandertaler noch eine dritte, unabhängig von diesen beiden Arten dorthin eingewanderte Population der Gattung Homo gegeben hat, die bisher noch keinen Artnamen erhalten hat.[55][56] Einer Forschergruppe um Svante Pääbo war es 2010 gelungen, zunächst mtDNA und später Zellkern-DNA aus einem 48.000 bis 30.000 Jahre alten Fingerknochen zu gewinnen. Aus ihren Daten wurde abgeschätzt, dass sich die Entwicklungslinien des Neandertalers und des Denisova-Menschen vor 640.000 Jahren endgültig trennten und dass die endgültige Trennung ihrer gemeinsamen Vorfahren von den Vorfahren der heutigen Afrikaner vor rund 800.000 Jahren erfolgte. Genetische Befunde deuten darauf hin, dass einige Ethnien in Ost-Indonesien, Australien, Papua-Neuguinea, Fidschi und Polynesien DNA der Denisova-Menschen aufweisen, dass es also zu einer Paarung zwischen deren Vorfahren und Denisova-Menschen gekommen ist.[57]
Hinweise auf eine Vermischung von Homo sapiens mit archaischen Verwandten in Afrika ergab eine Analyse des Genoms von 500 Afrikanern.[58] Die beobachteten genetischen Besonderheiten „bei Pygmäen- und Nicht-Pygmäen-Populationen“ unter anderem in Kamerun und in der Demokratischen Republik Kongo könnten jedoch auch infolge eines früheren genetischen Engpasses (eines genetischen Flaschenhals') zustande gekommen sein.[59]
Beim Flores-Menschen ist noch umstritten, ob er eine eigene Art darstellt. Sollte das der Fall sein, koexistierte noch vor 13.000 Jahren eine weitere Art der Gattung Homo neben dem Homo sapiens.
Siehe auch
Literatur
- Luigi Luca Cavalli-Sforza: Gene, Völker und Sprachen. Die biologischen Grundlagen unserer Zivilisation. Hanser, München 1999, ISBN 3-446-19479-7.
- Steve Olson: Herkunft und Geschichte des Menschen. Was die Gene über unsere Vergangenheit verraten. Berliner Taschenbuch, Berlin 2003, ISBN 3-8333-0130-9.
- Gary Stix: Wie hat sich die Menschheit ausgebreitet? In: Spektrum der Wissenschaft. Spektrumverlag, Heidelberg September 2009, ISSN 0170-2971, S. 58-65 (populärwissenschaftliche Darstellung).
- Spencer Wells: The Journey of Man. A Genetic Odyssey. Penguin, London 2003, ISBN 0-141-00832-6.
Einzelnachweise
- ↑ Julia Voss: Darwins Bilder: Ansichten der Evolutionstheorie. Fischer, Frankfurt 2007.
- ↑ Ein prominentes Beispiel bietet der damalige Präsident der American Association of Physical Anthropology Carleton Coon: The Origin of Races. Alfred A. Knopf, New York 1962.
- ↑ Rainer Grün, et al.: Direct dating of the Florisbad hominid. In: Nature. Bd. 382, 1996, S. 500–501.
- ↑ Ian McDougall et al.: Stratigraphic placement and age of modern humans from Kibish, Ethiopia. In: Nature, Band 433, 2005, S. 733–736; doi:10.1038/nature03258
- ↑ Friedemann Schrenk: Die Frühzeit des Menschen: Der Weg zum Homo sapiens. C. H. Beck, München 1997, S. 116f.
- ↑ Brenna M. Henn et al.: Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. In: Proceedings of the National Academy of Sciences, online-Vorabveröffentlichung vom 7. März 2011, doi:10.1073/pnas.1017511108
- ↑ Spencer Wells: The Journey of Man …, S. 98.
- ↑ Population Bottlenecks and Volcanic Winter
- ↑ Humans came 'close to extinction'
- ↑ Spencer Wells: The Journey of Man …, S. 64 und 66.
- ↑ Spencer Wells: The Journey of Man …, S. 15f.
- ↑ Arthur Mourant: The Distribution of the Human Blood Groups. Blackwell, Oxford 1954.
- ↑ Originalarbeit in Richard Lewontin: The Apportionment of Human Diversity. In: Evolutionary Biology. Bd. 6, 1972, S. 381–398. Ausführlich dazu Richard Lewontin: Menschen: genetische, kulturelle und soziale Gemeinsamkeiten. Spektrum der Wissenschaft, Heidelberg 1986.
- ↑ A. W. F. Edwards und L. L. Cavalli-Sforza: Reconstruction of evolutionary trees. In: V. E. Heywood und J. McNeill (Hrsg.): Phenetic und Phylogenetic Classification. The Systematics Association, London 1964, S. 67–76.
- ↑ Luigi Luca Cavalli-Sforza und Walter Bodmer: The Genetics of Human Populations. W. H. Freeman, San Francisco 1971.
- ↑ Rebecca L. Cann, Mark Stoneking und Allan C. Wilson: Mitochondrial DNA and human evolution. In: Nature. Bd. 325, 1987, S. 31–36.
- ↑ “All these mitochondrial DNAs stem from one woman who is postulated to have lived about 200,000 years ago, probably in Africa.“
- ↑ Spencer Wells: The Journey of Man …, S. 33.
- ↑ H. C. Harpending et al.: Mismatch distributions of mtDNA reveal recent human population expansions. In: Human Biology. Bd. 66, 1994, S. 761–775.
- ↑ a b Peter A. Underhill et al.: Y chromosome sequence variation and the history of human populations. In: Nature Genetics, Bd. 26, 2000, S. 358–361.
- ↑ P. C. Sabeti et al.: Positive Natural Selection in the Human Lineage. In: Science, Band 312, Nr. 5780, 2006, S. 1614–1620, doi:10.1126/science.1124309
- ↑ Jonathan K. Pritchard: Evolution auf der Kriechspur. In: Spektrum der Wissenschaft. Nr. 3, 2011, S. 28 ff..
- ↑ Zur Einführung: Luigi Luca Cavalli-Sforza: Gene, Völker und Sprachen. Die biologischen Grundlagen unserer Zivilisation. dtv, 2001, ISBN 978-3423330619
- ↑ Quentin D. Atkinson: Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa. In: Science, Band 332, Nr. 6027, 2011, S. 346–349, doi:10.1126/science.1199295
- ↑ Doron M. Behar et al.: The Dawn of Human Matrilineal Diversity. In: American Journal of Human Genetics, Bd. 82, 2008, S. 1130–1140.
- ↑ Gary Stix: Wie hat sich die Menschheit ausgebreitet? In: Spektrum der Wissenschaft. September 2009, S. 58-65, hier S. 64
- ↑ Vincent Macaulay u. a.: Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes. In: Science, Band 308, Nr. 5724, 2005, S. 1034–1036, doi:10.1126/science.1109792
- ↑ Curtis W. Marean et al.: Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. In: Nature. Bd. 449, 2007, S. 905-908 doi:10.1038/nature06204
- ↑ Spencer Wells: The Journey of Man …, S. 68.
- ↑ a b Michael Balter: New Work May Complicate History Of Neandertals and H. sapiens. In: Science, Bd. 326, Nr. 5950, 2009, S. 224f, doi:10.1126/science.326_224
- ↑ Simon J. Armitage u. a.: The Southern Route ‚Out of Africa‘: Evidence for an Early Expansion of Modern Humans into Arabia. In: Science, Band 331, 2011, S. 453–456, doi:10.1126/science.1199113
- ↑ idw-online vom 27. Januar 2011: Out of Africa – früher als bisher angenommen.
- ↑ Michael Petraglia et al.: Middle Paleolithic Assemblages from the Indian Subcontinent Before and After the Toba Super-Eruption. In: Science, Bd. 317, Nr. 5834, 2007, S. 114-116.
- ↑ Online-Artikel: Newly Discovered Archaeological Sites In India Reveals Ancient Life
- ↑ M. L. K. Murty: Recent Research on the Upper Palaeolithic Phase in India. In: Journal of Field Archaeology, Band 6, Nr. 3 (Herbst 1979), S. 301-320
- ↑ Peter Hiscock: Kolonisierung des 5. Kontinents. In: Archäologie in Deutschland 2/2002 S. 14–17
- ↑ Lluís Quintana-Murci et al.: Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa. In: Nature Genetics, Bd. 23, 1999, S. 437–441, doi:10.1038/70550
- ↑ Yoshan Moodley et al.: The Peopling of the Pacific from a Bacterial Perspective. In: Science, Bd. 323, Nr. 5913, 2009, S. 527–530, doi:10.1126/science.1166083
- ↑ Satish Kumar et al.: The earliest settlers' antiquity and evolutionary history of Indian populations: evidence from M2 mtDNA lineage. In: BMC Evolutionary Biology. Bd. 8, 2008, S. 230. doi:10.1186/1471-2148-8-230
- ↑ HUGO Pan-Asian SNP Consortium: Mapping Human Genetic Diversity in Asia. In: Science. Bd. 326, Nr. 5959, 2009, S. 1541-1545. doi:10.1126/science.1177074
- ↑ Hélène Rougier et al.: Peștera cu Oase 2 and the cranial morphology of early modern Europeans. In: PNAS, Band 104, Nr. 4, 2007, S. 1165-1170.
- ↑ Stefano Benazzi et al.: Early dispersal of modern humans in Europe and implications for Neanderthal behaviour. In: Nature, Online-Vorabveröffentlichung vom 2. November 2011, doi:10.1038/nature10617
- ↑ Sandrine Prat et al.: The Oldest Anatomically Modern Humans from Far Southeast Europe: Direct Dating, Culture and Behavior. In: PLoS ONE 6(6): e20834; doi:10.1371/journal.pone.0020834
- ↑ Eiszeit. Kunst und Kultur. Thorbecke, Ostfildern 2009, S. 93.
- ↑ V.V. Pitulko et al.: The Yana RHS Site: Humans in the Arctic Before the Last Glacial Maximum. In: Science, Band 303, Nr, 5654, 2004, S. 52-56.
- ↑ Michael R. Waters et al.: The Buttermilk Creek Complex and the Origins of Clovis at the Debra L. Friedkin Site, Texas. In: Science, Band 331, Nr. 6024, 2011, S. 1599–1603, doi:10.1126/science.1201855
eurekalert.org vom 24. März 2011: „Artifacts in Texas predate Clovis culture by 2,500 years, new study shows.“ (mit Abb.) - ↑ Guidon, Niède y G. Delibrias, Carbon-14 dates point to man in the Americas 32.000 years ago, Nature vol. 321 (1986): 769-771.
- ↑ John Noble Wilford, Findings Plunge Archaeology of the Americas Into Turmoil Estimates of first Asian migration are 33,000 years apart., New York Times vom 30. Mai 1989 (zur Auseinandersetzung unter den Archäologen)
- ↑ Jared Diamond: Collapse. How Societies Choose to Fail or Survive. Penguin, London 2005, S. 86f.
- ↑ Robert K. Headland: Die Erkundung der Antarktis. In: Arktis – Antarktis. Kunst- und Ausstellungshalle der Bundesrepublik Deutschland, Bonn 1997, S. 48–54.
- ↑ Jun Z. Li et al.: Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation. In: Science, Bd. 319, 2008, S. 1100–1104.
- ↑ Jared Diamond: Arm und Reich: Die Schicksale menschlicher Gesellschaften. S. Fischer, Frankfurt 2000.
- ↑ Die erste Veröffentlichung der Arbeitsgruppe Pääbo dazu ist M. Krings et al.: Neandertal DNA sequences and the origin of modern humans. In: Cell. Bd. 90, 1997, S. 19–30
- ↑ Richard E. Green et al.: A draft sequence of the Neandertal Genome. In: Science, Band 328, 2010, S.710–722, doi:10.1126/science.1188021
- ↑ Johannes Krause, Qiaomei Fu, Jeffrey M. Good, Bence Viola, Michael V. Shunkov, Anatoli P. Derevianko und Svante Pääbo: The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. In: Nature. Band 464, 2010, doi:10.1038/nature08976
- ↑ David Reich et al.: Genetic history of an archaic hominin group from Denisova Cave in Siberia. In: Nature, Band 468, Nr. 7327, 2010, S. 1053–1060, doi:10.1038/nature09710
- ↑ David Reich et al.: Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania. In: The American Journal of Human Genetics, Band 89, Nr. 4, 2011, S. 516–528, doi:10.1016/j.ajhg.2011.09.005
- ↑ Michael F. Hammer et al.: Genetic evidence for archaic admixture in Africa. In: PNAS, Band 108, Nr. 37. 2011, S. 15123–15128, doi:10.1073/pnas.1109300108
- ↑ Ann Gibbons: African Data Bolster New View of Modern Human Origin. In: Science, Band 334, Nr. 6053, 2011, S. 167, DOI:10.1126/science.334.6053.167








Wikimedia Foundation.