- Knabenkräuter (Dactylorhiza)
-
Knabenkräuter 
Geflecktes Knabenkraut (Dactylorhiza maculata)
Systematik Ordnung: Spargelartige (Asparagales) Familie: Orchideen (Orchidaceae) Unterfamilie: Orchidoideae Tribus: Orchideae Untertribus: Orchidinae Gattung: Knabenkräuter Wissenschaftlicher Name Dactylorhiza Neck. ex Nevski Die Knabenkräuter (Dactylorhiza), auch Fingerwurzen oder Kuckucksblumen genannt, bilden eine etwa 40 Arten umfassende Gattung in der Familie der Orchideen (Orchidaceae). Die krautige Pflanzen überwintern mit Hilfe einer handförmigen Knolle, ihre Blätter sind oft rötlich gefleckt, die Blüten sind meist rosa oder gelb. Die farbigen Blütenstände werden von Insekten besucht, enthalten aber keinen Nektar. Das Verbreitungsgebiet der Gattung umfasst ganz Europa, den äußersten Norden Afrikas, das nördliche und zentrale Asien, eine Art erreicht den Nordwesten Amerikas. Die Knabenkräuter der Gattung Dactylorhiza wurden erst spät von der ebenfalls Knabenkraut genannten Gattung Orchis unterschieden. Systematische Fragen bleiben bei der Abgrenzung zur Gattung Coeloglossum und bei der Unterscheidung einzelner Arten offen. Die Verwendung der Knollen als Salep bereitet Probleme für den Artenschutz, während die Nutzung einzelner Arten als Zierpflanzen auf gärtnerischer Vermehrung beruht.
Inhaltsverzeichnis
Beschreibung
Vegetative Merkmale
Es sind ausdauernde krautige Pflanzen mit Knollen als Überdauerungsorganen. Die unterirdische Knolle wird 10 bis 20 cm lang. Sie setzt sich aus einem 0,5 bis 1,5 cm kurzen Spross und dem anschließenden Wurzelgewebe zusammen.[1] Seitlich ist sie zusammengedrückt, bei einigen Arten tief bis fünfteilig gefingert mit relativ dünnen, langen Teilstücken, bei anderen Arten weniger unterteilt und eher einer rundlich-spindelförmigen Knolle genähert. Da häufig mehrere Stelen vorhanden sind, könnten diese Knollen aus mehreren miteinander verwachsenen Wurzeln entstanden sein.[2] Jährlich wird eine neue Knolle gebildet, die den Winter überdauert, während die alte abstirbt. Oberhalb der Knolle entspringen am Spross die Wurzeln.[3] Nur beim Krim-Knabenkraut (Dactylorhiza iberica) sind unterirdische Ausläufer vorhanden.[2]
Der Spross ist aufrecht und unbehaart, er wird im Laufe des Sprosswachstums hohl. Der Spross ist im Querschnitt wie folgt aufgebaut: Unter der Kutikula und der Epidermis folgt ein drei bis sechs Zelllagen dickes Chlorenchym mit großen Zellzwischenräumen, dann ein drei bis fünf Zelllagen dickes Sklerenchym, das auch die Leitbündel umfasst, schließlich in der Mitte ein Parenchym, welches aufgelöst wird, wodurch sich der Hohlraum im Stängel bildet.[2] An der Basis der Sprossachse stehen zweizeilig angeordnet einige Niederblätter, drei davon enthalten eine Erneuerungsknospe, von denen die oberste austreibt. Die Laubblätter stehen in einer lockeren Rosette bzw. sind im unteren Bereich des Sprosses verteilt. Sie sind rein grün oder grün mit rötlichen Flecken. Die Blattspitze endet oft zu einer kleinen Haube verwachsen.[1][3] Die Blattbasis umfasst den Spross. Bei vielen Arten sind die Blätter breit-oval, allerdings haben besonders in Hochlagen und nördlichen Gebieten verbreitete Arten schmale, längs der Mittelrippe gefaltete oder sichelförmig gekrümmte Blätter. Eine weitere Anpassung der Blätter, die in Trockengebieten und Hochlagen vorkommt, ist eine reduzierte Anzahl der Blätter bei rosettenartiger Häufung an der Basis der Sprosse. Der Blattrand weist eine nur unter dem Mikroskop sichtbare gezähnte Struktur mit gefalteter Kutikula auf. Auf der Unterseite der Blätter und am Spross befinden sich die Stomata, diese besitzen keine Nebenzellen.[2]
-

Knolle, Geflecktes Knabenkraut (Dactylorhiza maculata)
-

Rötlich gefleckte, große Blätter an der Basis, Geflecktes Knabenkraut
-

Kleine Blätter am Stängel verteilt, Geflecktes Knabenkraut
-

Ungeflecktes Blatt, Fleischfarbenes Knabenkraut (Dactylorhiza incarnata)
Blüte und Frucht
Oberhalb der Blätter setzt sich die Sprossachse als traubiger Blütenstand fort. Die Blüten stehen meist dicht beieinander, sie sind rosa, purpurn, gelb oder selten weiß gefärbt. Die Tragblätter sind laubblattartig und oft länger als die Blüte. Der ungestielte Fruchtknoten ist zylindrisch bis spindelförmig und verdreht, so dass die Blüten resupiniert sind. Das nach oben weisende Sepal und die seitlichen Petalen haften aneinander an und formen eine Haube über der Blüte. Die seitlichen Sepalen sind abgespreizt bis nach hinten geschlagen. Die Lippe ist deutlich dreilappig bis fast ganzrandig, an der Basis mit einem Sporn, der jedoch keinen Nektar enthält. Die Lippe ist häufig auf hellem Grund mit dunkleren Streifen oder Flecken versehen. Die Säule ist kurz, seitlich mit zwei kaum sichtbaren Staminodien, das fruchtbare Staubblatt enthält zwei keulenförmige Pollinien. Die Pollinien hängen über ein Stielchen (Caudicel) an je einer Klebdrüse (Viscidium), die von einer dünnen Hülle (Bursicula) umgeben ist.[3] Die Pollinien setzen sich aus mehreren Teilstücken (Massulae) zusammen, die jeweils aus vielen miteinander verklebten Pollenkörnern bestehen und eine Länge von 140 bis 300 µm bei einer Breite von 80 bis 200 µm erreichen. Die einzelnen Pollenkörner haben einen Durchmesser von 10 bis 20 µm, sie sind fest miteinander verklebt, die äußere Schicht (Exine) der Pollenkörner ist nur auf der Außenseite der Massulae ausgebildet, sie fehlt im Innern zwischen den verklebten Pollenkörnern.[2] Das Trenngewebe zwischen Staubblatt und Narbe (Rostellum) reicht bis zwischen die beiden Pollinien. Die Kapselfrucht enthält 2000 bis 5000 Samen.[3] Sie sind spindelförmig, 350 bis 1100 µm lang bei einem Durchmesser von 150 bis 300 µm. Die Samenschale besteht im Vergleich zur Gattung Orchis aus vielen kleinen Zellen. Diese stoßen ohne Zwischenräume aneinander, sie weisen auf der Oberfläche artspezifische Muster auf.[2] Die Samen behalten ihre Keimfähigkeit mehrere Jahre, was für eine Orchidee relativ lange ist.[4]
Besucht ein Insekt die Blüte so haftet die Klebscheibe meist im Kopfbereich und die Pollinien werden von dem Bestäuber aus dem Staubblatt herausgezogen. Nach kurzer Zeit trocknen dann die Stielchen, wodurch sich die Pollinien nach vorne biegen und beim nächsten Blütenbesuch die Narbe berühren.[2] Die Knabenkräuter werden häufig von Hummel-Königinnen (Bombus) bestäubt, die noch unerfahren sind und in den Blüten nach Nektar suchen. Die variable Zeichnung und Färbung der Blüten innerhalb einer Art könnte dabei helfen, ein schnelles Lernen der Insekten zu vermeiden. Die Bestäuber sind nicht artspezifisch, so dass es zu häufigen Hybriden kommt. Bei Fuchs’ Knabenkraut (Dactylorhiza fuchsii) und dem Gefleckten Knabenkraut (Dactylorhiza maculata) wurden auch Bockkäfer (Cerambycidae) als Bestäuber beobachtet, diese fressen Papillen auf der Lippe. Honigbienen nehmen an Blüten von Fuchs’ Knabenkraut zuckerhaltige Narbenflüssigkeit auf.[3]
-

Blütenstand, Breitblättriges Knabenkraut (Dactylorhiza majalis)
-

Einzelne Blüten, Geflecktes Knabenkraut (Dactylorhiza maculata)
-

Fruchtstand, Geflecktes Knabenkraut
-

Zwei Farbmorphen, Holunder-Knabenkraut (Dactylorhiza sambucina)
Mykorrhiza und Protokorm
Die Samen keimen nur mit Hilfe von Nährstoffen, die sie von einem Pilz beziehen. Es bildet sich ein Protokorm, das länglich konisch geformt ist, etwa wie eine Rübe.[5] Im Stadium des Protokorms ernährt sich die Pflanze mykotroph. Auch später, bei Pflanzen die Laubblätter besitzen, können Pilze in den Wurzeln und gelegentlich in den Enden der fingerförmig geteilten Knollen gefunden werden. Aus Wurzeln erwachsener Pflanzen wurden Pilze der Gattung Ceratobasidium sowie Thanetephorus orchidicola, Thanatephorus cucumeris und Tulasnella calospora isoliert. Unter Laborbedingungen keimen die Samen der Orchideen mit einigen der Pilze, die aus erwachsenen Pflanzen isoliert wurden. Die Keimraten und die weitere Entwicklung des Protokorms ist jedoch bei verschiedenen Pilzstämmen sehr unterschiedlich.[4]
Inhaltsstoffe
Die Blätter einiger untersuchter Arten enthalten verschiedene Flavonoide. Bei Fuchs’ Knabenkraut (Dactylorhiza fuchsii) wurden Kaempferol und Quercetin gefunden (die in der Pflanze wahrscheinlich als Glycoside vorliegen). Beim Fleischfarbenen Knabenkraut (Dactylorhiza incarnata) sind in den Blättern Flavon-Glycoside vorhanden, vom Holunder-Knabenkraut (Dactylorhiza sambucina) kennt man verschiedene Quercetin-Glucoside wie Isoquercitrin.
Die Blüten enthalten Anthocyane. Diese liegen als Cyanidin-Glucoside vor, Cyanin und Seranin als Diglucoside sowie weitere Farbstoffe – Orchicyanin I und II, Ophrysanin und Serapianin, die nach Orchideen benannt sind – die Oxalsäure über eine Acylgruppe enthalten.[3]
Zytologie
Die Chromosomenzahl bei Dactylorhiza beträgt 2n=40. Es gibt etliche polyploide Arten mit 2n=4x=80 und 2n=6x=120 Chromosomen. Hybriden zwischen diploiden und tetraploiden Individuen kommen vor (sie sind meist steril) und haben dann eine Chromosomenzahl von 2n=60, eine Zahl die auch beim Insel-Knabenkraut (Dactylorhiza insularis) vorkommt.[3] In einigen Populationen, etwa in der Verwandtschaft des Holunder-Knabenkrauts (Dactylorhiza sambucina) sowie bei Dactylorhiza aristata kommen regelmäßig Pflanzen mit einem Chromosomensatz von 2n=42 vor.[2] Beim Römischen Knabenkraut (Dactylorhiza romana) wurden Exemplare mit einem aneuploiden Chromosomensatz von 2n=40+1B festgestellt.[3] Weitere unregelmäßige Chromosomenzahlen, die nur selten vorkommen, sind 2n=100, 120, 122 (Dactylorhiza maculata, Dactylorhiza russowii) und 2n=41, 78 (Dactylorhiza maculata, Dactylorhiza fuchsii, Dactylorhiza umbrosa).[2] Die Chromosomen sind generell klein und ähneln in diesem Aspekt den Knabenkräutern der Gattung Orchis, die aber eine Zahl von 2n=42 besitzen.[3] Typisch für Dactylorhiza sind metazentrische, untereinander etwa gleich große Chromosomen.[2]
Lebenszyklus
Lebensablauf
Die Samen von Dactylorhiza keimen meist im Herbst, zur weiteren Differenzierung benötigen sie eine Kälteperiode[3] Junge Protokorme, die mykotroph leben, wurden bei mehreren Arten im Sommer gefunden. Im Herbst bildet sich die erste Wurzel, im nächsten Frühjahr kann der erste Spross mit Laubblättern gebildet werden. Dann geht die Pflanze zu sympodialem Wachstum über, das Rhizom wird frei von Pilzen, die dann auf die Wurzeln beschränkt sind.[4] Die Zeit, bis die ersten Laubblätter erscheinen, ist für verschiedene Arten unterschiedlich: dies kann schon in der ersten Vegetationsperiode passieren oder mehrere Jahre dauern. Eine Blüte erscheint bei den Jungpflanzen noch nicht (wie es bei der kurzlebigen Grünen Hohlzunge (Coeloglossum viride) der Fall ist), sondern erst nach weiteren Vegetationsperioden, die rein vegetativ verbracht werden. Die Zeitangaben von der Keimung bis zur ersten Blüte reichen von zwei bis 16 Jahren. Blühende Pflanzen blühen meist auch in der folgenden Vegetationsperiode wieder und sind über mehrere Jahre ausdauernd. Eine Population besteht so zum großen Teil (die Zahlenangaben reichen von 50 bis 85%) aus erwachsenen, blühenden Pflanzen. Besonders bei ungünstigen Umweltbedingungen können ausgewachsene Pflanzen wieder in das rein vegetative Stadium zurückfallen und die Blütenbildung einstellen. Selten kommt auch die sogenannte sekundäre Dormanz zum Tragen, bei der ausgewachsene Pflanzen ein oder mehrere Jahre nur mit ihren unterirdischen Organen überdauern. Viele Arten sind recht langlebig und vermehren sich in begrenztem Umfang auch vegetativ, insgesamt sind die Populationen für ihren Erhalt aber auf regelmäßige Vermehrung durch Samen angewiesen.[3]
Jahreszeitlicher Rhythmus
Der Austrieb der Blätter aus der Überdauerungsknolle erfolgt im Frühjahr, bei mediterranen Arten auch schon im Herbst. Gleichzeitig mit der oberirdischen Entwicklung der Blattrosette wird unterirdisch eine neue Knolle, selten zwei, angelegt, die während der Vegetationsperiode mit Reservestoffen gefüllt wird. Die Blüte erfolgt im Frühjahr oder Frühsommer, es folgt die Reife der Samen. Während im Süden die Wachstumsaktivität mit Einsetzen der Sommertrockenheit abgeschlossen wird, verschiebt sich weiter nördlich die Samenreife bis in den Herbst. Der alte Spross inklusive der Knolle, aus der er austrieb, sterben zum Ende der Vegetationsperiode ab. Die ungünstige Jahreszeit (trockener Sommer im mediterranen Bereich, kalter Winter weiter nördlich) wird mittels der neu gebildeten Knolle überdauert.[3]
Verbreitung
Areal
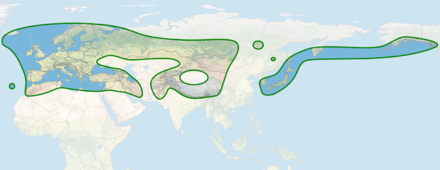 Verbreitungsgebiet der Gattung Dactylorhiza
Verbreitungsgebiet der Gattung Dactylorhiza
Die Dactylorhiza-Arten sind hauptsächlich in Europa in der gemäßigten Zone verbreitet, ebenso rund um das Mittelmeer einschließlich Nordafrikas und Kleinasiens. Dactylorhiza hatagirea als südöstlichste Art besitzt ein Verbreitungsgebiet am Südhang des Himalaya, Dactylorhiza aristata erreicht über den Osten Russlands die Aleuten, das Madeira-Knabenkraut (Dactylorhiza foliosa) markiert die südwestliche Verbreitungsgrenze, im Nordwesten wird noch Island erreicht. Dactylorhiza-Arten wachsen von Meereshöhe bis in Höhenlagen von 2500 Meter.[3]
Averyanov vermutet Vorfahren der Gattung Dactylorhiza in der Poltawa-Flora aus dem Paläogen. Diese Pflanzen hätten, aufgrund des wärmeren Klimas, noch keine unterirdischen Knollen besessen. Die Speicherorgane könnten sich dann während der Hebung der Alpen als Anpassung auf das Klima der Hochlagen entwickelt haben. Gegen Ende des Pliozän, als das Klima in Europa generell kühler wurde, besiedelten die Vorfahren von Dactylorhiza weiträumig tiefere Lagen. Auch der sommertrockene Mittelmeerraum konnte so erreicht werden, ermöglicht durch die in den Hochlagen erworbene Fähigkeit, kühlere und trockenere Bedingungen zu überstehen. Während der häufig wechselnden klimatischen Bedingungen der Eiszeiten schwankten die Areale einzelner Arten. Populationen wurden getrennt, es kam zu allopatrischer Artbildung. Während günstiger Perioden konnte die Gattung ihre Verbreitung quer durch Sibirien bis an den Pazifik ausdehnen; seither sind durch das zunehmend trockenere Klima im Innern Asiens Lücken im Areal entstanden.[2] Als Refugien während der Eiszeiten kommen Griechenland, die Iberische Halbinsel und Nordafrika in Frage. Die Besiedlung Mitteleuropas nach den Eiszeiten erfolgte von dort, daher ist die genetische Variabilität der nördlichen Pflanzen nur ein Ausschnitt der im Mittelmeerraum vorkommenden.[6]
Misst man die Diversität anhand der Anzahl beschriebener Arten, so bildet Nordwesteuropa ein Zentrum der Artenvielfalt. Etwa in einem Raum, der die Britischen Inseln sowie den Süden Skandinaviens umfasst und sich nach Süden bis zu den Alpen zieht, finden sich einerseits zahlreiche weit verbreitete Arten, andererseits auch Sippen mit eng umgrenztem Areal. Die hier vorhandenen endemischen Sippen sind jungen Ursprungs: entstanden einerseits durch die variable Vergletscherung während der Eiszeit, andererseits durch Hybridisierung. Die weiter verbreiteten Arten strahlen vor allem nach Osten aus, die osteuropäischen und westsibirischen Arten haben meist ihren Ursprung in Nordwesteuropa. Die Bezüge nach Süden zum Mittelmeerraum sind weniger ausgeprägt, dort kommen meist andere Arten als in Nordwesteuropa vor.
Eine hohe Artenvielfalt findet man auch in den Karpaten und auf dem Balkan. Hier gibt es eine Reihe von Arten, die ebenfalls in Nordwesteuropa vorkommen, aber auch etliche endemische Sippen. Ein drittes Mannigfaltigkeitszentrum liegt in Kleinasien: Die Arten, die hier vorkommen, haben kaum Bezüge zu den europäischen Dactylorhiza-Arten, sie kommen meist ausschließlich in Kleinasien und dem Kaukasus vor. Die Endemiten dieser beiden Gegenden sind älteren Ursprungs, die Arten haben ihre Areale nicht so verändert oder ausgedehnt wie einige europäische.
In Mittelasien finden sich noch zwei Regionen, in denen mehrere Dactylorhiza-Arten heimisch sind: einmal der Hindukusch sowie im Pamir. Hier kommen zu weit verbreiteten Arten mit europäischem Zentrum weitere, dort endemische Arten.[2]Die Verteilung der genetischen Diversität weicht allerdings von der Verteilung der Artenanzahl ab. Im östlichen Mittelmeerraum, im Kaukasus und auf der Krim herrscht die größte genetische Vielfalt, obwohl in diesem Areal nicht sonderlich viele Arten beschrieben sind. Im Nordwesten Europas, wo zahlreiche Arten bekannt wurden, gibt es dagegen auf der Ebene des Genoms nur eine geringe Vielfalt. In diesem Gebiet wurden viele nahe verwandte Sippen und Hybriden als eigenständige Arten beschrieben.[6]
Standorte
Die Standorte liegen meist in offenen, sonnigen Situationen oder im Halbschatten. Dabei kann es sich um unterschiedliche Habitate wie Trockenrasen, Dünen, Wiesen, Sümpfe, Moore, lichte Gebüsche und lichte Wälder handeln. Während einige Arten wie Dactylorhiza maculata und Dactylorhiza fuchsii sowohl an feuchten wie an trockenen Standorten gefunden wurden, sind andere Arten an bestimmte Biotope gebunden. Gleiches gilt etwa für den pH-Wert des Bodens.[3]
Gefährdung und Artenschutz
Bei vielen Populationen der Knabenkräuter ist ein Rückgang zu beobachten. Gründe dafür sind das Trockenlegen von Sümpfen und Mooren sowie eine Intensivierung der Landwirtschaft mit Düngung von Wiesen und Weiden. Auch geänderte Nutzung von Weideflächen, die zu Verbuschung und Beschattung führt, kann zum Rückgang der Knabenkräuter führen. In Asien ist das Sammeln der Knollen eine Gefährdung, besonders Dactylorhiza hatagirea aus dem Himalaya ist dadurch stark bedroht. Nur wenige Arten, etwa Fuchs’ Knabenkraut (Dactylorhiza fuchsii) und das Übersehene Knabenkraut (Dactylorhiza praetermissa), besiedeln gestörte Standorte und können sich dort eine gewisse Zeit halten.[6]
Systematik
Äußere Systematik
Innerhalb der Orchideen wird die Gattung Dactylorhiza in die Unterfamilie Orchidoideae und dort in die Tribus Orchideae eingeordnet. Die nächste Verwandtschaft besteht zur Gattung der Händelwurzen (Gymnadenia) und zur Grünen Hohlzunge (Coeloglossum viride). Die meisten verwandten Gattungen kommen ebenfalls schwerpunktmäßig in Europa vor, mit Ausnahme der Klade aus Waldhyazinthen (Platanthera) und Galearis, die Verbreitungszentren in Ostasien und Nordamerika besitzt. Die Verwandtschaftsbeziehungen lassen sich mit einem Kladogramm wie folgt darstellen:[3]
Knabenkräuter (Dactylorhiza) + Grüne Hohlzunge (Coeloglossum viride)
Händelwurzen (Gymnadenia)
Höswurzen (Pseudorchis)
Waldhyazinthen (Platanthera)
Galearis
Kugelorchis (Traunsteinera)
Zwergstendel (Chamorchis alpina)
Verwandtschaft zur Grünen Hohlzunge
Besonders nah mit Dactylorhiza verwandt ist die Grüne Hohlzunge, für die eine eigene Gattung, Coeloglossum, aufgestellt wurde. Erste Untersuchungen der ribosomalen DNA platzierten die Grüne Hohlzunge innerhalb der Gattung Dactylorhiza. Folgerichtig wurde sie von Coeloglossum viride in Dactylorhiza viridis umbenannt. Der Gattungsname Dactylorhiza, der jüngeren Datums ist als Coeloglossum, wurde als nomen conservandum vorgeschlagen.[3] Eine Untersuchung der DNA der Chloroplasten erbrachte keine Auflösung der nah verwandten Arten. Das Ergebnis einer neueren Studie, die weitere DNA-Abschnitte einbezog, sieht die Grüne Hohlzunge als Schwestertaxon zur Gattung Dactylorhiza und weiterhin als eigenständige Gattung.[7]
Coeloglossum viride als Schwestergruppe zu Dactylorhiza Coeloglossum viride innerhalb Dactylorhiza Dactylorhiza incarnata - Gruppe
restliche Dactylorhiza
Coeloglossum viride
Dactylorhiza incarnata - Gruppe
Coeloglossum viride
restliche Dactylorhiza
Innere Systematik
In der Gattung Dactylorhiza gibt es diploide und tetraploide Pflanzen, wobei die Zuordnung der diploiden zu einzelnen Arten nicht so strittig ist wie die Beurteilung der tetraploiden Sippen. Die Verdoppelung des Chromosomensatzes kann als Autopolyploidie vorliegen, das heißt, dass nur eine Ausgangsart an der Bildung der tetraploiden Art beteiligt war, oder als Allopolyploidie, bei der die tetraploiden Pflanzen durch Hybridisierung zweier diploider Elternarten entstanden. Aus derselben Kombination zweier Elternarten sind zu verschiedenen Zeiten und an verschiedenen Orten wiederholt Hybriden hervorgegangen, die sich äußerlich und in ihren ökologischen Ansprüchen unterscheiden können, genetisch aber wenige Unterschiede aufweisen.[8] Die Abgrenzung insbesondere der tetraploiden Knabenkraut-Arten ist schwierig, da sie eine große Variationsbreite, die zwischen den Ausgangsarten vermittelt, zeigen. Neben alten, stabilisierten Hybridschwärmen gibt es häufig spontane Hybriden, die stellenweise zahlreicher als die Elternarten sind.[3] Die Entschlüsselung der Verwandtschaftsverhältnisse durch molekulargenetische Methoden wird durch die wiederholte Hybridisierung und durch die geringen genetischen Unterschiede innerhalb der Gattung erschwert.[9]
Leonid Averyanov legte eine Einteilung der Gattung in vier Sektionen vor. Zwei davon enthielten nur je eine Art, das Krim-Knabenkraut (Dactylorhiza iberica) in der Sektion Iberanthus sowie Dactylorhiza aristata in der Sektion Aristatae. Die Sektion Sambucinae war ebenfalls artenarm, die Mehrzahl der Arten stand in der Sektion Dactylorhiza, die durch Subsektionen noch feiner unterteilt wurde.[2] Molekulargenetische Untersuchungen konnten die von Averyanov vorgeschlagene Unterteilung großteils nicht stützen. Neben den tatsächlich relativ isolierten drei artenarmen Sektionen Averyanovs (Dactylorhiza iberica, Dactylorhiza aristata sowie Dactylorhiza romana und Dactylorhiza sambucina) gibt es drei weitere große Gruppen: zwei hauptsächlich diploide Gruppen um Dactylorhiza incarnata sowie Dactylorhiza maculata und die Gruppe der tetraploiden Hybriden zwischen diesen beiden.[8] In der Darstellungsweise eines klassischen Kladogramms lassen sich die Beziehungen durch Hybridisierung nicht darstellen, die beiden nachfolgenden enthalten deshalb nur eine Auswahl diploider Arten:
Pillon et al 2006[6] Pillon et al 2007[8] Dactylorhiza foliosa
Dactylorhiza maculata
Dactylorhiza fuchsii
Dactylorhiza saccifera
Dactylorhiza aristata
Dactylorhiza romana
Dactylorhiza sambucina
Dactylorhiza iberica
Coeloglossum viride
Dactylorhiza incarnata
Dactylorhiza euxina
Dactylorhiza foliosa
Dactylorhiza maculata
Dactylorhiza fuchsii
Dactylorhiza saccifera
Dactylorhiza sambucina
Dactylorhiza romana
Dactylorhiza aristata
Coeloglossum viride
Dactylorhiza incarnata
Dactylorhiza euxina
Dactylorhiza iberica
Liste der Dactylorhiza-Arten
Alphabetische Liste der Dactylorhiza-Arten:[10]
- Dactylorhiza alpestris (Pugsley) Aver.
- Dactylorhiza aristata (Fisch. ex Lindl.) Soó
- Dactylorhiza atlantica Kreutz & Vlaciha
- Dactylorhiza baldshuanica Chernyak.
- Dactylorhiza baumanniana J.Hölzinger & Künkele
- Dactylorhiza cordigera (Fr.) Soó
- Dactylorhiza cordigera subsp. bosniaca (Beck) Soó
- Dactylorhiza cordigera subsp. cordigera
- Dactylorhiza cordigera var. graeca (H.Baumann) Presser
- Dactylorhiza cordigera subsp. pindica (B.Willing & E.Willing) H.Baumann & R.Lorenz
- Dactylorhiza cordigera var. rhodopeia Presser
- Dactylorhiza czerniakowskae Aver.
- Dactylorhiza durandii (Boiss. & Reut.) M.Laínz
- Dactylorhiza elata (Poir.) Soó
- Dactylorhiza elata subsp. elata
- Dactylorhiza elata subsp. sesquipedalis (Willd.) Soó
- Dactylorhiza euxina (Nevski) Czerep.
- Dactylorhiza euxina subsp. armeniaca (Hedrén) Kreutz
- Dactylorhiza euxina subsp. euxina
- Dactylorhiza foliosa (Rchb.f.) Soó
- Dactylorhiza fuchsii (Druce) Soó
- Dactylorhiza fuchsii subsp. carpatica (Batoušek & Kreutz) Kreutz
- Dactylorhiza fuchsii subsp. fuchsii
- Dactylorhiza fuchsii subsp. hebridensis (Wilmott) Soó
- Dactylorhiza fuchsii subsp. okellyi (Druce) Soó
- Dactylorhiza fuchsii subsp. psychrophila (Schltr.) Holub
- Dactylorhiza fuchsii subsp. sooana (Borsos) Borsos
- Dactylorhiza graggeriana (Soó) Soó
- Dactylorhiza hatagirea (D.Don) Soó
- Dactylorhiza iberica (M.Bieb. ex Willd.) Soó
- Dactylorhiza incarnata (L.) Soó
- Dactylorhiza incarnata subsp. coccinea (Pugsley) Soó
- Dactylorhiza incarnata subsp. cruenta (O.F.Müll.) P.D.Sell
- Dactylorhiza incarnata subsp. gemmana (Pugsley) P.D.Sell
- Dactylorhiza incarnata subsp. incarnata
- Dactylorhiza incarnata subsp. lobelii (Verm.) H.A.Pedersen
- Dactylorhiza incarnata subsp. ochroleuca (Wüstnei ex Boll) P.F.Hunt & Summerh.
- Dactylorhiza incarnata subsp. pulchella (Druce) Soó
- Dactylorhiza insularis (Sommier) Ó.Sánchez & Herrero
- Dactylorhiza irenica F.M.Vázquez
- Dactylorhiza isculana Seiser
- Dactylorhiza kafiriana Renz
- Dactylorhiza kafiriana subsp. baumgartneriana (B.Baumann & al.) Sczep. & Kreutz
- Dactylorhiza kafiriana subsp. kafiriana
- Dactylorhiza kalopissii E.Nelson
- Dactylorhiza kalopissii subsp. kalopissii
- Dactylorhiza kalopissii subsp. macedonica (J.Hölzinger & Künkele) Kreutz
- Dactylorhiza kalopissii subsp. pythagorae (Gölz & H.R.Reinhard) Kreutz
- Dactylorhiza kulikalonica Chernyak.
- Dactylorhiza lapponica (Laest. ex Hartm.) Soó
- Dactylorhiza lapponica subsp. angustata (Arv.-Touv.) Kreutz
- Dactylorhiza lapponica subsp. lapponica
- Dactylorhiza lapponica subsp. rhaetica H.Baumann & R.Lorenz
- Dactylorhiza maculata (L.) Soó
- Dactylorhiza maculata subsp. battandieri (Raynaud) H.Baumann & Künkele
- Dactylorhiza maculata subsp. caramulensis Verm.
- Dactylorhiza maculata subsp. elodes (Griseb.) Soó
- Dactylorhiza maculata subsp. ericetorum (E.F.Linton) P.F.Hunt & Summerh.
- Dactylorhiza maculata subsp. islandica (Á.Löve & D.Löve) Soó
- Dactylorhiza maculata subsp. maculata
- Dactylorhiza maculata subsp. podesta (Landwehr) Kreutz
- Dactylorhiza maculata subsp. savogiensis (D.Tyteca & Gathoye) Kreutz
- Dactylorhiza maculata subsp. schurii (Klinge) Soó
- Dactylorhiza maculata subsp. transsilvanica (Schur) Soó
- Dactylorhiza magna (Czerniak.) Ikonn.
- Dactylorhiza majalis (Rchb.) P.F.Hunt & Summerh.
- Dactylorhiza majalis subsp. baltica (Klinge) H.Sund.
- Dactylorhiza majalis subsp. majalis
- Dactylorhiza majalis subsp. sphagnicola (Höppner) H.A.Pedersen & Hedrén
- Dactylorhiza nieschalkiorum H.Baumann & Künkele
- Dactylorhiza osmanica (Klinge) P.F.Hunt & Summerh.
- Dactylorhiza osmanica var. anatolica (E.Nelson) Renz & Taubenheim
- Dactylorhiza osmanica var. osmanica
- Dactylorhiza praetermissa (Druce) Soó
- Dactylorhiza praetermissa subsp. osiliensis (Pikner) Kreutz
- Dactylorhiza praetermissa subsp. praetermissa
- Dactylorhiza purpurella (T.Stephenson & T.A.Stephenson) Soó
- Dactylorhiza purpurella var. cambrensis (R.H.Roberts) R.M.Bateman & Denholm
- Dactylorhiza purpurella var. purpurella
- Dactylorhiza romana (Sebast.) Soó
- Dactylorhiza romana subsp. georgica (Klinge) Soó ex Renz & Taubenheim
- Dactylorhiza romana subsp. guimaraesii (E.G.Camus) H.A.Pedersen
- Dactylorhiza romana subsp. romana
- Dactylorhiza russowii (Klinge) Holub
- Dactylorhiza saccifera (Brongn.) Soó
- Dactylorhiza saccifera subsp. gervasiana (Tod.) Kreutz
- Dactylorhiza saccifera subsp. saccifera
- Dactylorhiza salina (Turcz. ex Lindl.) Soó
- Dactylorhiza sambucina (L.) Soó
- Dactylorhiza sudetica (Poech ex Rchb.f.) Aver.
- Dactylorhiza traunsteineri (Saut. ex Rchb.) Soó
- Dactylorhiza traunsteineri subsp. curvifolia (F.Nyl.) Soó
- Dactylorhiza traunsteineri subsp. traunsteineri
- Dactylorhiza traunsteineri subsp. turfosa (F.Proch.) Kreutz
- Dactylorhiza umbrosa (Kar. & Kir.) Nevski
- Dactylorhiza urvilleana (Steud.) H.Baumann & Künkele
- Dactylorhiza viridis (L.) R.M.Bateman, Pridgeon & M.W.Chase
Botanische Geschichte
Die Knabenkräuter der Gattung Dactylorhiza wurden erst spät von denen der Gattung Orchis unterschieden. Obwohl der heutige wissenschaftliche Name schon 1790 von Necker geprägt wurde, geriet er in Vergessenheit und gilt nicht als gültige Erstbeschreibung (Neckers Elementa botanica ist als opus utique opressum, als nicht zu verwendendes Werk, im ICBN gelistet). Auch die Arbeit von Nevski, der 1937 die Erstbeschreibung in der Flora der UdSSR veröffentlichte, blieb noch ohne Resonanz. So verwendete Vermeulen 1947 in seiner Dissertation Studies on Dactylorchids, die ein grundlegendes Werk für die Erforschung der Gattung war, den neuen Namen Dactylorchis. Die Trennung von der Gattung Orchis wurde damit weithin anerkannt, der Name Dactylorhiza fand in den folgenden Jahren Verbreitung, etwa in dem von Karlheinz Senghas 1968 herausgegebenen Werk Probleme der Gattung Dactylorhiza.
Der Name leitet sich von griech. δάκτυλος dactylos = Finger und ρίζα rhiza = Wurzel her und beschreibt die Knollen, welche zusammengedrückt und mehrteilig-handförmig sind. Der gebräuchlichste deutsche Name „Knabenkraut“ entstand, als die heutigen Gattungen Orchis und Dactylorhiza noch nicht unterschieden wurden. Er wird auch weiterhin für die Arten der Gattung Dactylorhiza verwandt. Die Bezeichnung „Kuckucksblumen“, eine weitere alte Bezeichnung für die Knabenkräuter, wurde von Rothmaler 1958 als Name für die Gattung Dactylorhiza vorgeschlagen. Gelegentlich wird zur Abgrenzung von der Gattung Orchis die Verwendung des Namens „Fingerwurzen“ empfohlen, der die wörtliche Übersetzung des botanischen Namens wiedergibt.[11]
Aberglaube und Verwendung
Im Aberglauben galten die am Johannistag ausgegrabenen Knollen („Johannishändchen“) als Glücksbringer.
Die Knollen der Orchis- und Dactylorhiza-Arten und einiger anderer Orchideen wurden früher als Schleimdroge („Tubera Salep“) in der Kinderheilkunde bei Reizerscheinungen des Magen-Darmtraktes angewandt, als es noch keine besseren Arznei- und Heilmittel gab. Heute werden Orchideenarten nicht mehr als wirksame Heilkräuter angesehen. Zudem stehen alle Arten der Gattungen Orchis und Dactyorhiza unter strengstem Naturschutz, was insbesondere für die unterirdischen Pflanzenteile gilt.
Literatur
- Leonid V. Averyanov: A Review of the Genus Dactylorhiza. In: Orchid Biology. Reviews and Perspectives. Bd. 5, Timber Press, Portland, Oregon 1990, ISBN 0-88192-170-X, S. 159–206.
- Fritz Füller: Die Gattungen Orchis und Dactylorhiza. Orchideen Mitteleuropas, 3. Teil. In: Die Neue Brehm-Bücherei. Bd. 286, A. Ziemsen Verlag, Wittenberg 1983, ISSN 0138-1423.
- Alec M. Pridgeon, Phillip Cribb, Mark W. Chase, Finn Rasmussen (Hrsg.): Genera Orchidacearum. Orchidoideae (Part one). Oxford University Press, New York und Oxford 2001, ISBN 0-19-850710-0, S. 279–284.
- Karlheinz Senghas, Hans Sundermann (Hrsg.): Probleme der Gattung Dactylorhiza. In: Jahresberichte des Naturwissenschaftlichen Vereins in Wuppertal. Heft 21/22, Wuppertal 1968.
Einzelnachweise
- ↑ a b M. G. Vakhrameeva, I. V. Tatarenko, T. I. Varlygina, G. K. Torosyan, M. N. Zagulskii: Orchids of Russia and Adjacent Countries. A.R.G. Gantner, Ruggell 2008, ISBN 978-3-906166-61-2, S. 58ff, 211ff.
- ↑ a b c d e f g h i j k l m Leonid V. Averyanov: A Review of the Genus Dactylorhiza. In: Orchid Biology. Reviews and Perspectives. Bd. 5, Timber Press, Portland, Oregon 1990, ISBN 0-88192-170-X, S. 159–206.
- ↑ a b c d e f g h i j k l m n o p q Alec M. Pridgeon, Phillip Cribb, Mark W. Chase, Finn Rasmussen (Hrsg.): Genera Orchidacearum. Orchidoideae (Part one). Oxford University Press, New York und Oxford 2001, ISBN 0-19-850710-0, S. 279–284.
- ↑ a b c Hanne N. Rasmussen: Terrestrial orchids from seed to mycotrophic plant. Cambridge University Press, Cambridge 1995, ISBN 0-521-04881-8, S. 295–303.
- ↑ Hanne N. Rasmussen: Terrestrial orchids from seed to mycotrophic plant. Cambridge University Press, Cambridge 1995, ISBN 0-521-04881-8, S. 121.
- ↑ a b c d Yohan Pillon, Michael F. Fay, Alexey B. Shipunov, Mark W. Chase: Species diversity versus phylogenetic diversity: A practical study in the taxonomically difficult genus Dactylorhiza (Orchidaceae). In: Biological Conservation. Bd 129, Nr. 1, 2006, S. 4-13, doi:10.1016/j.biocon.2005.06.036.
- ↑ Nicolas Devos, Olivier Raspé, Anne-Laure Jacquemart, Daniel Tyteca: On the monophyly of Dactylorhiza Necker ex Nevski (Orchidaceae): is Coeloglossum viride (L.) Hartman a Dactylorhiza?. In: Botanical Journal of the Linnean Society. Bd. 152, 2006, S. 261–269.
- ↑ a b c Yohan Pillon, Michael F. Fay, Mikael Hedrén, Richard M. Bateman, Dion S. Devey, Alexey B. Shipunov, Michelle van der Bank, Mark W. Chase: Evolution and temporal diversification of western European polyploid species complexes in Dactylorhiza (Orchidaceae). In: Taxon. Bd 56, Nr. 4, 2007, S. 1185-1208.
- ↑ Richard M. Bateman, Peter M. Hollingsworth, Jillian Preston, Luo Yi-Bo, Alec M. Pridgeon, Mark W. Chase: Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae). In: Botanical Journal of the Linnean Society. Bd 142, 2003, S. 1–40, doi:10.1046/j.1095-8339.2003.00157.x.
- ↑ World Checklist of Dactylorhiza. In: The Board of Trustees of the Royal Botanic Gardens, Kew.
- ↑ Fritz Füller: Die Gattungen Orchis und Dactylorhiza. Orchideen Mitteleuropas, 3. Teil. In: Die Neue Brehm-Bücherei. Bd. 286, A. Ziemsen Verlag, Wittenberg 1983, ISSN 0138-1423.
Weblinks
 Commons: Dactylorhiza – Album mit Bildern und/oder Videos und Audiodateien
Commons: Dactylorhiza – Album mit Bildern und/oder Videos und Audiodateien- Die Orchideen der Rhön
- Orchideen der Südsteiermark Orchis und Dactylorhiza Knabenkräuter
- Dactylorhiza Bilddatenbank
siehe:
-









Wikimedia Foundation.