- Innere Atmung
-
Als aerobe Atmung (Zellatmung, innere Atmung) werden Stoffwechselprozesse in Zellen von Lebewesen bezeichnet, die in einer Oxidation von Stoffen mit molekularem, elementarem Sauerstoff (O2) als Oxidationsmittel bestehen und dem Energiegewinn dienen. Insbesondere versteht man hierunter die biochemischen Vorgänge der Atmungskette in der inneren Membran der Mitochondrien, an deren Ende ATP synthetisiert wird. Andere Formen der Atmung – im Sinne des Gasaustausches von Organismen – werden unter dem Begriff der äußeren Atmung zusammengefasst.
Inhaltsverzeichnis
Übersicht
- Der Einheitlichkeit halber werden für Säuren als Zwischenstufen – wie in der Biochemie üblich – die Namen ihrer Anionen verwandt („Pyruvat“ statt „Brenztraubensäure“). Die Summenformeln beziehen sich jedoch auf die undissoziierten Verbindungen. Der Anhang „-P“ bei den Summenformeln steht für eine Phosphatgruppe.
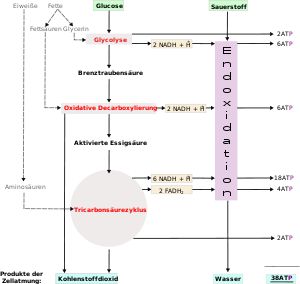 Übersicht über die Zellatmung.
Übersicht über die Zellatmung.Im Folgenden wird die Nutzung der Energie aus der Oxidation von Glucose (Traubenzucker) durch Zellen dargestellt. Zellen können Energie auch durch Oxidation anderer Stoffe gewinnen, die Oxidation von Glucose ist jedoch die am häufigsten genutzte Energiequelle.
Zellen nehmen zu ihrer Energieversorgung Glucose auf. Sie wird von Eukaryoten im Cytoplasma und in den Mitochondrien vollständig zu Kohlenstoffdioxid und Wasser oxidiert. Die Summengleichung lautet:
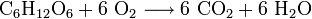
- Aus einem Molekül Glucose und sechs Molekülen Sauerstoff werden sechs Moleküle Kohlenstoffdioxid und sechs Moleküle Wasser.
Die Änderung der Freien Energie unter Standardbedingungen, jedoch pH = 7, beträgt bei dieser Reaktion ΔG0' = −2880 kJ je Mol Glucose[1]. Weichen die tatsächlichen Bedingungen von diesen Standardbedingungen ab, so ist auch der Betrag der Änderung der Freien Energie ein anderer, er kann erheblich vom Standardwert abweichen. In lebenden Systemen sind Standardbedingungen in der Regel nicht gegeben und ändern sich oft auch während der Stoffumsetzung. Der Betrag der Änderung der Freien Energie unter Standardbedingungen bietet also bei Lebewesen lediglich einen Anhaltspunkt für die bei einer chemischen Stoffumsetzung frei werdende Energie.
Bei dieser chemischen Stoffumsetzung werden in einer Reihe von komplizierten Reaktionsschritten – darunter viele Redoxreaktionen – Wasserstoffatome von den Abbauprodukten der Glucose-Moleküle abgetrennt und mit Hilfe von Wasserstoffüberträgern (Nicotinamid-Adenin-Dinucleotid, NAD) zu den Mitochondrien transportiert. Dort reagieren die Wasserstoffatome in der Atmungskette mit Sauerstoff zu Wasser (gelegentlich, nicht ganz zutreffend, als „biologische Knallgasreaktion“ bezeichnet); die Glucose-Moleküle werden letztlich vollständig oxidiert. Am Ende des Abbauprozesses gewinnt die Zelle mit Hilfe der bei der biologischen Wasserstoff-Oxidation frei werdenden Energie die energiereiche Verbindung Adenosintriphosphat (ATP). Sie dient als Energieüberträger und kurzfristiger Energiespeicher und ist für viele Stoffwechselvorgänge als universelle Energiequelle erforderlich.
Die zusammenfassende Reaktionsgleichung der Zellatmung entspricht von rechts nach links gelesen der Reaktionsgleichung für die oxygene Photosynthese.
Ablauf des Prozesses
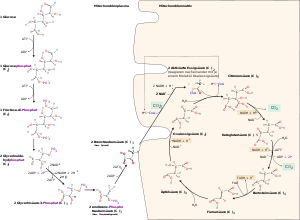 Glykolyse und Citratzyklus
Glykolyse und CitratzyklusDie Zellatmung ist ein Prozess, bei dem energiereiche in energiearme Stoffe umgesetzt werden. Im Fall der Zellatmung wird meistens das Glucosemolekül C6H12O6 in einer längeren Reihe von Schritten zu C1-Körpern (CO2) und Wasser (H2O) oxidiert. Dieser Abbau kann in vier Abschnitte unterteilt werden:
- die Glykolyse,
- die oxidative Decarboxylierung,
- den Citratzyklus (Citronensäurezyklus) und
- die Endoxidation in der Atmungskette.
Die Gesamtbilanz der Zellatmung kann folgendermaßen formuliert werden:
- Aus einem Molekül Glucose und sechs Molekülen Sauerstoff werden sechs Moleküle Kohlenstoffdioxid und sechs Moleküle Wasser.
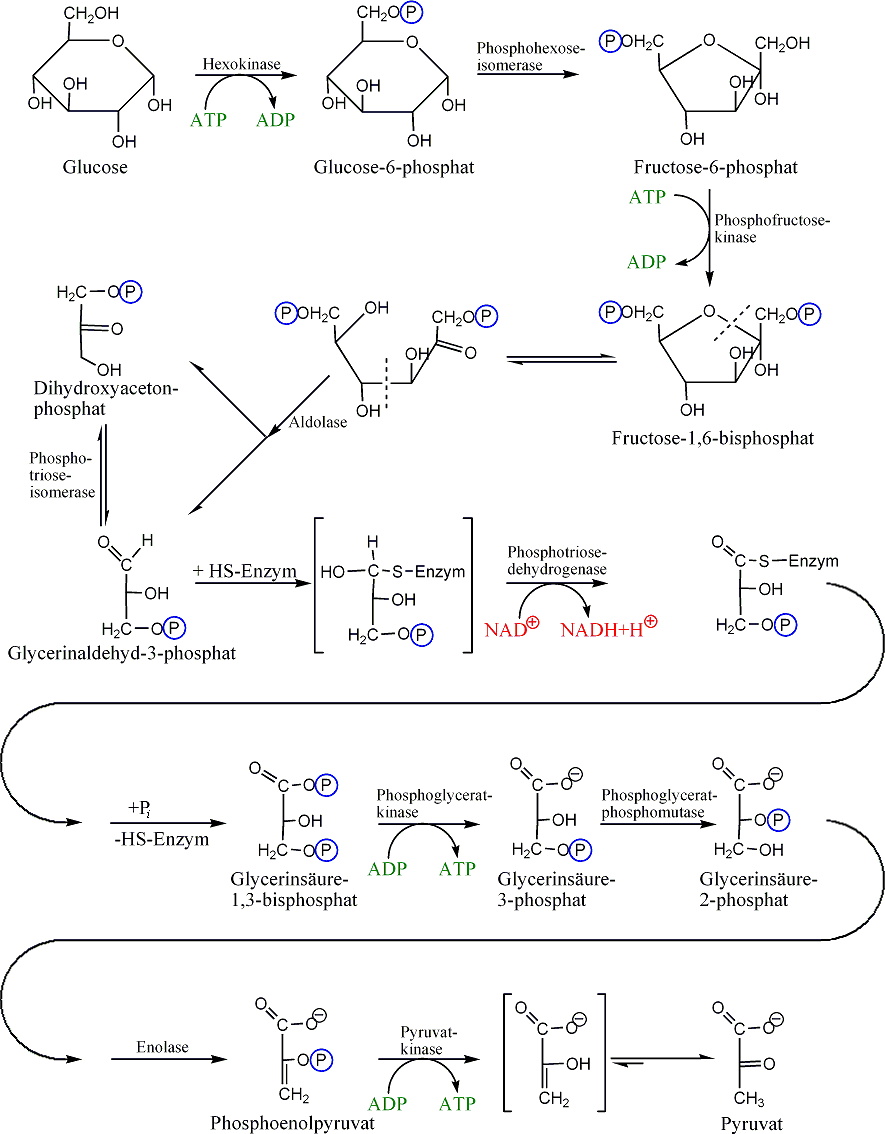
Glykolyse
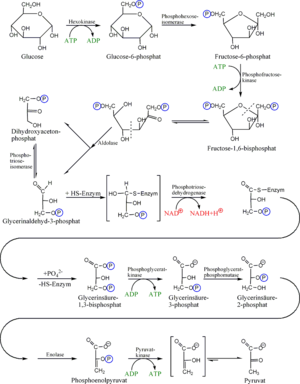 Glykolyse
Glykolyse- Hauptartikel: Glykolyse
Die Glykolyse (= Zuckerzerlegung) läuft im Cytoplasma ab. Bei diesem Vorgang wird das Glucose-Molekül gespalten. Dies geschieht durch zweifache Phosphorylierung, also der Abspaltung eines H unter Anlagerung eines Phosphatrestes (P) von ATP, so dass erst Glucose-6-phosphat (C6H11O6–P) entsteht (durch Strukturveränderung wird dieses in Fructose-6-phosphat umgewandelt) und dann Fructose-1,6-bisphosphat (C6H10O6–PP). Es werden also 2 Moleküle ATP zu 2 Molekülen ADP dephosphoryliert. Durch die Phosphorylierung der Glucose ist sie nun im aktivierten Zustand. Dieser C6-Körper wird dann in in zwei C3-Körper gespalten. Es entstehen ein Molekül Dihydroxyacetonphosphat (DHAP) und ein Molekül Glycerinaldehyd-3-phosphat (GAP, siehe Glycerinaldehyd), beide haben die gleiche Summenformel C3H5O3–P. Nur das Glycerinaldehyd-3-phosphat wird weiter abgebaut, deswegen wird das DHAP in dieses umgewandelt. Es gehen also immer zwei GAP-Moleküle in den weiteren Abbau. Ein weiteres Phosphat (freies Phosphat, nicht aus ATP) wird angelagert und GAP wird oxidiert, wodurch 1,3-Bisphosphoglycerat (C3H4O4–PP, siehe Phosphoglycerinsäure) entsteht. Die Elektronen werden auf den Wasserstoffüberträger NAD+ (Nicotinamidadenindinukleotid in der oxidierten Form) übertragen. Im weiteren Schritt wird ein Phosphatrest (P) auf ADP übertragen, so dass ATP und 3-Phosphoglycerat (PGS, C3H5O4–P) entstehen. Durch Abspaltung von Wasser entsteht nun Phosphoenolpyruvat (PEP, C3H3O3–P). Im letzten Schritt wird auch der letzte Phosphatrest (P) auf ADP übertragen, so dass Pyruvat (C3H4O3) und ATP entstehen. Auf dem Weg von 1,3-Bisphosphoglycerat zum Pyruvat werden also je Molekül 1,3-Bisphosphoglycerat 2 Moleküle ATP durch Phosphorylierung von ADP gebildet.
Bilanz der Glykolyse
Oxidative Decarboxylierung
- Hauptartikel: Oxidative Decarboxylierung
Die Oxidative Decarboxylierung ist ein kurzer Schritt, der allerdings unabdingbar für den darauf folgenden Schritt ist. Sie läuft bei Eukaryoten in der Mitochondrienmatrix ab. Aus dem Pyruvat wird durch einen komplizierten Reaktionsmechanismus ein CO2 abgespalten (Decarboxylierung) und 2 H-Atome auf NAD+ übertragen (Redoxreaktion) sowie die dadurch entstehende Essigsäure (Acetat) an das Coenzym A (CoA) gebunden, so dass Acetyl-CoA entsteht.
Bilanz der Oxidativen Decarboxylierung:
Citratzyklus (oder Citronensäurezyklus)
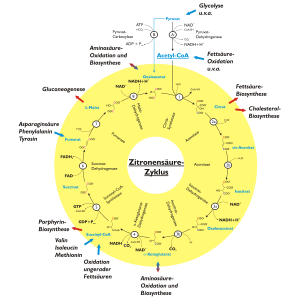 Citronensäuresäurezyklus
Citronensäuresäurezyklus- Hauptartikel: Citratzyklus
Der Citratzyklus, auch als Citronensäurezyklus oder Tricarbonsäurezyklus bezeichnet, ist nach dem ersten Zwischenprodukt benannt, dem Citrat, dem Anion der Citronensäure, einer dreiwertigen Carbonsäure. Im letzten Schritt des Citratzyklus entsteht Oxalacetat (C4H4O5). Nur sie ist fähig, sich mit Acetyl-CoA zu verbinden und – durch Aufnahme von Wasser und Abspaltung von Coenzym A – Citrat (C6H8O7) zu bilden. Dabei wird also das Coenzym A wieder regeneriert. Erneut wird CO2 abgespalten und ein Reduktionsäquivalent NADH und das α-Ketoglutarat (C5H6O5) gebildet. Im nächsten Schritt erfolgt erneut eine Abspaltung von CO2 und die Bildung von NADH mit Hilfe des Coenzyms A. Der C3-Körper, der in die Reaktion eingegangen ist, ist erst an dieser Stelle komplett gespalten worden. Die nächsten Schritte dienen nur noch der Bildung von Oxalacetat, damit der Zyklus wieder von vorn beginnen kann. Dies geschieht über die Moleküle Succinyl-CoA (C4H5O3–CoA), Succinat (C4H6O4), Fumarat (C4H4O4), Malat (C4H6O5) und dann durch Oxidation (Bildung eines Reduktionsäquivalentes) zum Oxalacetat.
Bilanz des Citratzyklus (läuft zweimal ab, da aus 1 Mol Glucose 2 Mol Pyruvat und somit auch 2 Mol Acetyl-Coenzym A gebildet werden):
Endoxidation in der Atmungskette
Durch den bisherigen Prozess sind 4 ATP entstanden. Den größten Teil der ATP-Ausbeute liefert jedoch die Atmungskette mit Hilfe der Reduktionsäquivalente. Insgesamt stehen 10 NADH (zwei aus der Glykolyse und acht (2 mal 4) aus dem Citratzyklus) und 2 FADH2 (Flavinadenindinukleotid) zur Verfügung.
Ein NADH kann 2 Elektronen (e−) abgeben, wodurch der am NAD gebundene Wasserstoff als Proton (H+) frei wird und das verbleibende NAD-Molekül positiv geladen wird: NAD+. Weil die so abgegebenen 2 Elektronen auf einem recht hohen Energieniveau liegen (sehr niedriges Redoxpotential des Redoxpaares NADH/NAD+), können mit ihrer Hilfe 10 Protonen aus der Matrix in den Intermembranraum transportiert werden. Dies geschieht folgendermaßen: Die 2 Elektronen des NADH reduzieren den ersten Komplex (Komplex I) von mehreren Enzym-Komplexen der Atmungskette, die sich zwischen Matrix und Intermembranraum des Mitochondriums befinden. Jedes Elektron wird nun über Redoxreaktionen von einem Enzym-Komplex zum nächsten weitergegeben. Aufgrund der Weitergabe von Elektronen von Komplex zu Komplex wird dieser Vorgang auch als Elektronentransportkette bezeichnet. Durch den Komplex I, den Komplex III und den Komplex IV werden H+-Ionen (Protonen) aus der Matrix in den Intermembranraum transportiert. Im Intermembranraum entsteht auf diese Weise eine hohe Wasserstoffionen-Konzentration, wodurch ein pH-Wert unter 7 entsteht, und es entsteht ein osmotisches Potential. Die Redoxreaktionen und die Entstehung des osmotischen Potentials zusammen werden Chemiosmose genannt: Die Redoxreaktionen sind chemische Umsetzungen, der Unterschied der H+-Konzentrationen von Matrix und Intermembranraum stellt ein osmotisches Potential dar.
Die Protonen fließen schließlich durch die membrangebundene ATP-Synthase vom Intermembranraum in den Matrixraum zurück. Dieses Enzym katalysiert die Synthese von ATP aus einem Phosphatrest und ADP. Die in der protonenmotorischen Kraft steckende Durchflussenergie wird dazu genutzt, dass entstandenes ATP von der ATP-Synthase freigesetzt wird. Der Transport eines Moleküls ADP aus dem Cytoplasma in die Matrix bzw. umgekehrt der Transport eines Moleküls ATP in das Cytoplasma wird durch eine ATP/ADP-Translokase katalysiert. Für diesen Transport wird jedoch auch der Protonengradient angezapft, so dass für die Verfügbarkeit von ATP bzw. ADP ein Proton verbraucht wird. Damit müssen mindestens 4 Protonen für die Erzeugung eines Moleküls ATP berechnet werden.
Durch die Oxidation von einem NADH entstehen somit 2,5 ATP. Ausnahme sind die zwei NADH aus der Glykolyse. Diese befinden sich noch im Cytoplasma und müssen erst in die Mitochondrien transportiert werden. Erfolgt dies mit Hilfe des Glycerin-3-Phosphat-Shuttles, gewinnt man aus diesen nur je 1,5 ATP. Da 8 + 2 NADH oxidiert werden, entstehen insgesamt 8 × 2,5 + 2 × 1,5 = 23 ATP.[2] Wird jedoch cytosolisches NADH durch den Malat-Aspartat-Shuttle in die Matrix gebracht, können aus diesen Reduktionsäquivalenten analog je 2,5 mol ATP erzeugt werden. Damit können maximal 10 × 2,5 = 25 ATP erzeugt werden.[3]
Mit dem FADH2 verläuft der Vorgang im Prinzip genauso, nur gibt FADH2 auf einem höheren Redoxpotential und damit niedrigeren Energieniveau Elektronen ab. Dessen Elektronen können also erst auf einer energetisch niedriger stehenden Stufe in die Atmungskette eingeschleust werden. Deshalb können mit Hilfe der Elektronen des FADH2 nur 4 Protonen aus der Matrix in den Intermembranraum gepumpt werden. Mit einem FADH2 werden infolgedessen nur 1,5 ATP gebildet. Da zwei FADH2 oxidiert werden, entstehen dabei 3 ATP.[2]
Die Protonen und die Elektronen des NADH und des FADH2 (insgesamt 24) werden zusammen mit 6 Molekülen O2, die durch die Membran in die Mitochondrienmatrix transportiert werden, zu 12 H2O oxidiert. Die Elektronen- bzw. Wasserstoffüberträger NAD+ und FAD können durch Aufnahme von je 2 e– und 2 H+ wieder zu NADH bzw. FADH2 reduziert werden.
Bilanz der Atmungskette:
Energiebilanz
Schritt Coenzym-Ausbeute ATP-Ausbeute ATP-Quelle Glykolyse Vorbereitungsstufe −2 für die Zerlegung der Glucose in 2 Moleküle Glycerinaldehyd-3-phosphat verbrauchte Energie Glykolyse Ertragsstufe 4 Substratkettenphosphorylierung 2 NADH 3 Oxidative Phosphorylierung. Oxidative Decarboxylierung 2 NADH 5 Oxidative Phosphorylierung Citratzyklus 2 Substratkettenphosphorylierung (in Form von GTP) 6 NADH 15 Oxidative Phosphorylierung 2 FADH2 3 Oxidative Phosphorylierung Gesamtausbeute 30 ATP[3][2]–32 ATP[3] pro Molekül Glucose (Glycerin-3-phosphat-Shuttle bzw. Malat-Aspartat-Shuttle) Da Prokaryoten keine Zellkompartimente besitzen, müssen sie nicht Energie für intrazelluläre Transportvorgänge ausgeben. Daher können sie maximal 38 Moleküle ATP aus einem Molekül Glucose erzeugen.[3]
Quellen
- ↑ R. Wehner, W. Gehring: Zoologie. 24. Auflage. Georg Thieme Verlag, Stuttgart, New York 2007, ISBN 978-3-13-367424-9.
- ↑ a b c d e Berg, Stryer, Tymoczko: Biochemie. Spektrum Akademischer Verlag, 2007, ISBN 978-3827418005
- ↑ a b c d Reginald Garrett und Charles M. Grisham: Biochemistry. (International Student Edition). Thomsom Learning Inc.; 3. Auflage 2005; ISBN 0-534-41020-0; S. 669f.
Literatur
- Reginald Garrett und Charles M. Grisham: Biochemistry. (International Student Edition). Thomsom Learning Inc.; 3. Auflage 2005; ISBN 0-534-41020-0; S. 640–674


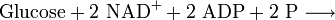
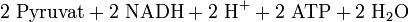
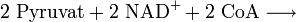
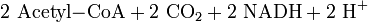

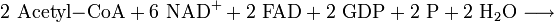
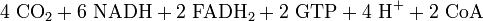
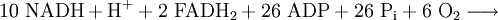
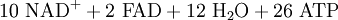
Wikimedia Foundation.