- Ammonshorn (Fossil)
-
Ammoniten  Historische Rekonstruktion lebender Ammonitentiere von Heinrich Harder. Als überholt gilt die Deutung der Arme als Cirren und der Anaptychen als Gehäusedeckel.
Historische Rekonstruktion lebender Ammonitentiere von Heinrich Harder. Als überholt gilt die Deutung der Arme als Cirren und der Anaptychen als Gehäusedeckel.Zeitraum Unterdevon bis Oberkreide Fossilfundorte - Weltweit
Systematik Tiere (Animalia) Vielzellige Tiere (Metazoa) Urmünder (Protostomia) Weichtiere (Mollusca) Kopffüßer (Cephalopoden) Wissenschaftlicher Name Ammonoidea Zittel, 1884 Untertaxa - Palaeoammonoidea
- Mesoammonoidea
- Neoammonoidea
Die Ammoniten sind eine ausgestorbene Gruppe ausschließlich mariner Kopffüßer (Cephalopoda). Die Gruppe war sehr artenreich; über 1500 Gattungen sind bekannt, die Zahl der Arten dürfte bei etwa 30.000 bis 40.000 liegen. Die Größe der Schale ausgewachsener Tiere lag meist im Bereich von Zentimetern, Parapuzosia seppenradensis ist mit ca. 1,80 Meter Schalendurchmesser die größte bekannte Art.
Ammoniten stellen von ihrem ersten Auftreten im Unterdevon bis zu ihrem Aussterben am Ende der Kreide (Kreide-Tertiär-Grenze) eine große Zahl der Leitfossilien; zum Teil erfolgt die zeitliche Abgrenzung mariner Sedimente ausschließlich anhand von Ammoniten. Sie sind für die Geologie und die Paläontologie daher von großer Bedeutung. Aufgrund ihrer Schönheit, Vielfalt und Häufigkeit sind sie auch bei vielen Fossiliensammlern beliebt und entsprechend häufig im Fossilienhandel zu finden.
Inhaltsverzeichnis
Namensgebung
In die zoologische Systematik wurde die Unterklasse Ammonoidea 1884 von Karl Alfred von Zittel (1839–1904) eingeführt. Die Bezeichnung stammt aus der Antike, Plinius der Ältere bezeichnete Versteinerungen als Ammonshörner. Die ägyptische Gottheit Amun, auch Amon oder Ammon, wird auch als Widder mit entsprechenden Hörnern dargestellt. Die mit Wülsten versehenen und eingedrehten Hörner dieser Huftiere erinnern an Ammoniten. Die Bezeichnung ceras in vielen wissenschaftlichen Namen von Ammoniten leitet sich vom griechischen Wort κέρας für Horn ab. Einige europäische Ortschaften tragen Ammoniten im Stadtwappen, so etwa das französische Villers-sur-Mer in der Normandie, das britische Whitby, die Gemeinde Gosau in Österreich oder die deutschen Gemeinden Lüdinghausen (Westfalen) und Schernfeld (Bayern).
Biologie
Anatomie: Schale und Weichteile
 Discoscaphites conradi aus der Oberkreide in Schalenerhaltung
Discoscaphites conradi aus der Oberkreide in Schalenerhaltung Längsschnitt durch den gekammerten Teil.
Längsschnitt durch den gekammerten Teil.Das wichtigste und zugleich häufig das einzige erhaltene fossilierte Merkmal stellt die Schale der Tiere dar. Sie ist mehrschichtig und besteht aus dem dünnen Periostracum aus Conchiolin, einer aus Aragonit aufgebauten mittleren Prismenschicht sowie der inneren Perlmuttschicht (Hypostracum). Die Grundform des Gehäuses von Ammoniten ist eine in einer Ebene aufgerollte Spirale, wobei sich die Ränder der einzelnen Windungen mehr (involut) oder weniger (evolut) umfassen. Diese Schalenform wird als planspiral bezeichnet.
Die Schale aller Ammoniten ist in zwei Bereiche unterteilt, die Wohnkammer und den gekammerten Auftriebskörper (Phragmokon). Von außen sind, bei Steinkernen, die ebenfalls aus Perlmutt bestehenden Trennwände als Lobenlinien (Sutur) deutlich zu erkennen und stellen ein wichtiges Unterscheidungsmerkmal der verschiedenen Ammonitenarten und Gruppen dar. Die Kammern waren bei lebenden Tieren mit Gas gefüllt. Der Regulation des Gases diente wie bei den rezenten Perlbooten ein Sipho, der allerdings anders als bei den Nautiloiden nicht in der Mitte der Trennwände, sondern an der Außenkante verläuft. Durch das Ein- bzw, Auslassen des Gases in die Septen war es dem Ammoniten möglich, im Wasser auf und ab zu steigen.
Um ein Wachstum des Tieres zu ermöglichen, wurden zum einen der Schalenrand an der Mündung vergrößert und zum anderen im hinteren Teil der Wohnkammer neue Kammern abgetrennt. Der Weichteilkörper befand sich hauptsächlich in der Wohnkammer und war wie bei Nautilus mittels großer Muskeln mit ihr verwachsen. Außerhalb der Wohnkammer befanden sich die Arme sowie der Kopf mit den Augen und dem Kiefer und die Mündung des Trichters.
 Nachbildung des größten bislang aufgefundenen Ammoniten, aufgestellt in Seppenrade
Nachbildung des größten bislang aufgefundenen Ammoniten, aufgestellt in SeppenradeDie Größen der Schalen variieren stark. Die größten Schalen von Ammoniten mit einem Durchmesser von rund 1,80 Meter wurden bislang in der Westfälischen Bucht gefunden. Sie gehören zur Art Parapuzosia seppenradensis. Die ersten Exemplare wurden 1887 und 1895 in einem Steinbruch bei Seppenrade entdeckt, wonach die Art benannt wurde. Das größte erhaltene Schalengehäuse kam beim U-Bahn-Bau in Dortmund ans Tageslicht. Die Funde stammen aus kreidezeitlichen Mergel-Schichten.
Arme
Alle rezenten Cephalopoden besitzen Arme, entweder 8 oder 10 wie etwa Oktopus oder Sepien, oder zahlreiche Cirren wie Nautilus. Die Arme von Oktopus oder Sepia sind mit Saugnäpfen besetzt, die von Belemniten mit kleinen Haken, Nautilus besitzt weder Haken noch Saugnäpfe sondern ein klebriges Sekret. Hieraus ergibt sich für Ammoniten eine ganze Reihe an Möglichkeiten. Am Wahrscheinlichsten scheint eine relativ kleine Anzahl an Armen mit Saugnäpfen zu sein.
Radula, Aptychen und Anaptychen (Nahrungsaufnahme)
 Aptychus aus dem Malm (Solnhofen)
Aptychus aus dem Malm (Solnhofen)Der Bau von Mund und „Kiefern“ war bei Ammoniten dem anderer Weichtiere wie Schnecken oder den enger verwandten lebenden Kopffüßern ähnlich. Fossil erhaltungsfähig sind insbesondere Hartteile. Nur in Ausnahmefällen wurden die Zähnchen der Raspelzunge (Radula) gefunden, relativ häufiger sind isolierte Teile des Kiefers wie Aptychen und Anaptychen. Da die Radula von Cephalopoden funktional nur geringem evolutionärem Druck unterliegt, sie dient im wesentlichen dem Weitertransport von Nahrung beim Schlingvorgang, kann sie nur wenig zur Deutung beitragen. Die Radula der Ammoniten war schmal und lang, die Zahl, Größe und Gestalt der Zähnchen der Radula variierte besonders mit der Körpergröße.
Aptychen, die man lange in Analogie zu Bildungen bei Nautilus, als Gehäusedeckel deutete, sind durch neuere Untersuchungen als Teil des Unterkiefers identifiziert, die anders als die Kiefer rezenter Kopffüßer allerdings nicht zum Schneiden oder Beißen geeignet waren. Die in ihrer Gestalt an Muscheln erinnernden Aptychen bestehen aus zwei spiegelsymmetrischen Schalen, die z. T. sehr solide sind und Wachstumsstreifen aufweisen. Anhand von Serienschliffen ließen sie sich als Stützstrukturen des verhornten Unterkiefers erkennen, der mit einem ebenfalls verhornten Oberkiefer zusammenwirkt. Die Radula befindet sich zwischen den Kieferteilen. Aptychen finden sich vom Lias bis zum Aussterben der Ammoniten in der Kreide. Die nur hornig ausgebildeten Anaptychen gehören zum Unterkiefer anderen Ammonitenarten, die vom Oberdevon bis zum oberen Lias lebten. U. a. anhand der Gestaltung von Radula und Aptychen sowie von in Ausnahmefällen fossilisierten Resten des Mageninhaltes lässt sich die Nahrungsbiologie der Ammoniten erschließen. Die Aptychen dienten vermutlich zum Hineinschaufeln von Nahrung. Aufgrund der Physik der Ammonitenschale (siehe unten) waren Ammoniten sicher keine schnellen Jäger. Allenfalls Lauerjagd kommt für die langsamen Ammoniten in Frage. Die oft kleinen Wohnkammern bieten nicht genug Platz für einen langen Pflanzenfresserdarm. Gegen eine vegetabile Ernährung spricht auch die Verbreitung in oberflächenfernen Meeresteilen, in denen Pflanzen fehlen.
Als Nahrung kommen daher Plankton, Kammerlinge (Foraminifera), kleinere Ammoniten (bis rund 1/10 der Größe des Prädators), Krebse, Muschelkrebse, Stachelhäuter (z. B. Armglieder von Seelilien), Brachiopodenbrut oder Aas und eventuell auch Korallen und Bryozooen in Frage. Auffällig ist, dass nach dem Erlöschen der Ammoniten in der Oberkreide die Blütezeit der marinen Schnecken beginnt.
Weitere Weichteile (Tintenbeutel, Kiemen und Augen)
Nachweise von Tintenbeuteln bei Ammoniten sind nicht eindeutig. Höchstwahrscheinlich liegen Verwechslungen mit Magen und Ösophagus der Ammoniten vor. Kiemen sind fossil nicht überliefert. Nautiloiden haben 4 Kiemen, die anderen rezenten Kopffüßer nur 2 Kiemen. Aufgrund der Verwandtschaftsverhältnisse wird vermutet, dass Ammoniten 2 Kiemen besaßen. Augen sind fossil nicht überliefert, man vermutet aber, dass es sie gab. Die Frage ist, ob sie mit ihnen nur schwarz-weiß oder farbig sehen konnten. Nautiloiden besitzen Augen ohne Linsen, die nach dem Prinzip der Lochkamera funktionieren. Rezente Kopffüßer besitzen verschiedene Formen von geschlossenen Linsenaugen.
Rhyncholithen
Rhyncholithen, die aufgrund ihrer Ähnlichkeit mit den Kiefern von Nautilus früher als Kieferteile von Ammoniten gedeutet wurden, sind nach heutiger Auffassung sicher keine Ammonitenkiefer, sondern vermutlich Kiefer von anderen Cephalopoden, eine Zuordnung zu einzelnen Gruppen ist aber noch nicht möglich.
Fortbewegung
Aufgrund des Baues und der Gestaltung der planspiralen Schale, sowie aus dem Vergleich mit lebenden Kopffüßern, kann das Schwimmvermögen der Ammoniten grob erschlossen werden. Aus der Form der Ammonitenschale ergeben sich drei wichtige Größen: Die Lage des Auftriebszentrums, der Schwerpunkt und die Gestaltung der Wohnkammer. Je geringer der Abstand zwischen Auftriebszentrum und Schwerpunkt ist, desto einfacher gerät das Tier bei kräftigem Vortrieb in Rotation. Dies bedeutet, dass Ammoniten nur dann relativ schnell sein konnten, wenn der Abstand groß war. Lebende Nautilusse besitzen einen relativ großen Abstand und zeigen allenfalls eine – verglichen mit Kalmaren – langsame und taumelnde Schwimmbewegung. Die Gestaltung der Wohnkammer ist entscheidend, da das in ihr befindliche Wasser mit großer Geschwindigkeit und Druck durch den Trichter ausgepresst werden muss, um eine hohe Beschleunigung zu ermöglichen. Lange Wohnkammern ermöglichen allenfalls einen langsamen Wasseraustritt, schmale Wohnkammern besitzen nicht genügend Raum für die Aufnahme einer großen Wassermenge und der notwendigen Muskulatur für einen schnellen, kräftigen Wasseraustritt. Die alte Idee, dass Ammoniten mit hochmündigen, glatten Schalen aufgrund ihres geringen Wasserwiderstandes als schnelle Schwimmer zu betrachten sind, ist daher unzutreffend. Auch die vertikale Bewegung der Ammoniten war sicher nicht schnell. Die Änderung der Gas- bzw. Flüssigkeitsmenge im gekammerten Teil der Schale zur Regulation des statischen Auftriebs war langsam, Nautilus benötigt dafür Stunden. Moderne Rekonstruktionen von Ammoniten sehen in ihnen daher weniger schnelle Jäger, als eher schneckenartig langsame Tiere die Bewohner des Bodens oder des offenen Wassers waren und starke Wasserbewegungen, wie sie in der Brandung auftreten, mieden.
Geschlechtsdimorphismus: Mikrokonche und Makrokonche
Bei einigen Ammoniten lag vermutlich ein starker baulicher Unterschied bei Männchen und Weibchen vor, bei dem sich die kleineren Männchen (Mikrokonch) sogenannte Zwergmännchen von den größeren Weibchen (Makrokonch) unterscheiden lassen. Um dieses zu belegen, müssen ausgewachsene Ammoniten der gleichen Art verglichen werden. Dies geschieht über die Bildung einer morphologischen Reihe. Dass eine Ammonitenschale einem ausgewachsenem Tier und nicht nur einem kleinen noch nicht erwachsenen Tier gehörte, lässt sich an den Lobenlinien erkennen. Bei erwachsenen Tieren verringert sich der Abstand zwischen den letzten Lobenlinien, es kommt zur Lobendrängung.
Auch in weiteren Schalenmerkmalen wird ein Geschlechtsdimorphismus vermutet. An der Gehäusemündung besitzen einige Arten Schalenverlängerungen die „Visier“ genannt werden, diese könnten kleinere Männchen vor Kannibalismus der Weibchen geschützt haben.
Verwandtschaft und Stammesgeschichte
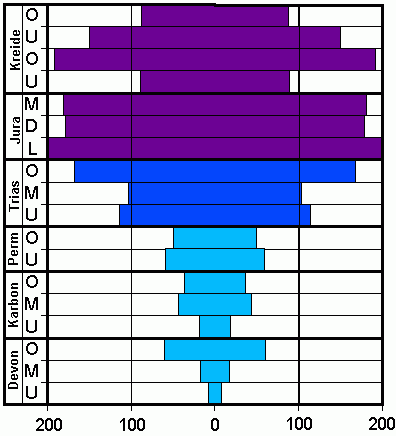 Verteilung von rund 1500 Ammonitengattungen von Devon bis Kreide, hellblau: vor allem Goniatiten, blau: vor allem Ceratiten, violett: vor allem Ammoniten
Verteilung von rund 1500 Ammonitengattungen von Devon bis Kreide, hellblau: vor allem Goniatiten, blau: vor allem Ceratiten, violett: vor allem AmmonitenDie Ammoniten gehen ebenso wie die Belemniten aus der Gruppe der Bactriten hervor. Drei große, artenreiche Gruppen, die Goniatiten, die Ceratiten und die Ammoniten, folgen aufeinander. Am Ende des Perm und der Trias starben jeweils bis auf wenige Gattungen alle Vertreter der Ammoniten aus. Dem Aussterben folgte rasch eine schnelle Neubildung von Arten und Gattungen. In der Oberkreide starben alle Ammoniten ohne Nachfahren aus.
In der Frühphase der Evolution der Ammoniten veränderte sich die grundlegende Gestalt der Schale von einer primär gestreckten Form, die bereits gekammert war, in die planspirale Form, der der überwiegende Teil der Gattungen und Arten angehört. Aber auch diese Form war nicht frei von evolutionären Entwicklungen.
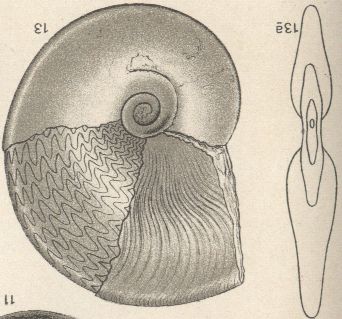
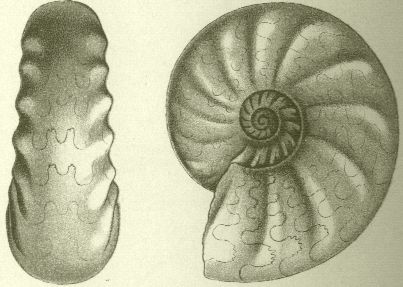
So nahm etwa die Gestaltung der Lobenlinie an Komplexität zu, wie z. B. Vertreter der großen Gruppen „Goniatiten“ (Devon-Perm), „Ceratiten“ (Trias) und eigentliche „Ammoniten“ (Jura-Kreide) zeigen. Die Abbildungen wurden in die Lebendstellung der Ammonitentiere gedreht. Die Komplexität der Lobenlinie nimmt auch im Laufe der Ontogenese zu.

Vertreter der Goniatiten
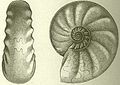
Vertreter der Ceratiten

Vertreter der Ammoniten
Die Lobenlinie ist bei als Steinkernen erhaltenen Ammoniten zu erkennen (bei Exemplaren in Schalenerhaltung muss die Schale abpräpariert werden). Neben dieser Tendenz finden sich eine Unzahl an Variationen der Proportion der Schale, von „Verzierungen“ wie Rippen, Spaltrippen, Wülste, Rillen, Dornen oder Knoten, z. T. als Folgen von Konvergenz.
Typogenese
Ammoniten waren das Musterbeispiel der überholten Typostrophenlehre, wie sie Otto Heinrich Schindewolf vertrat. Eine Typostrophe beginnt mit der Entstehung einer neuen Form (Typogenese), die dann im Laufe der Zeit im Rahmen ihrer Entwicklungspotenzen ausgestaltet wird (Typostase). Schließlich gelangt die Form an die Grenzen ihrer Möglichkeiten und stirbt aus (Typolyse). Die Evolution der Ammoniten folgt scheinbar diesem Schema. Beispielsweise ist in dieser Deutung das Auftreten von heteromorphen Ammonitenarten, die keine planspiralen Gehäuse besitzen, in der Oberkreide eine Typolyse, d. h. eine stammesgeschichtliche Degeneration. Spätere Funde belegen das Auftreten solcher Formen auch in anderen Epochen.
Aussterben
Vor dem endgültigen Aussterben der Ammoniten an der Kreide-Tertiär-Grenze gab es zwei weitere Massenaussterben: Das erste in der rund 10 Millionen Jahre dauernden Permkrise, bei der rund 75–90 % aller Tierarten ausstarben. Das zweite Massensterben der Ammoniten liegt an der Trias-Jura Grenze. Bei beiden Massensterben haben nur wenige Ammonitenarten überlebt, die danach schnell eine Vielzahl neuer Familien, Gattungen und Arten entwickelten.
Die Ursachen für diese Massenaussterben sind umstritten, klimatische und astronomische Ursachen (Meteoriteneinschläge, KT-Impakt) werden ebenso diskutiert wie u. a. aufgrund von Kontinentaldrift veränderte Meeresströmungen mit tiefgreifenden Wechsel im Nahrungsangebot, Temperaturverteilung im Meer und der Wassertiefe – also rapiden Änderungen der Ammonitenhabitate.
Schmuck aus Ammoniten
 Ammolit als Schmuckstein
Ammolit als SchmucksteinAus fossilen Überresten von Ammoniten kann sich der als Ammolit bezeichnete opaleszierende Edelstein bilden, der im Schmuckhandel auch unter den Namen Calcentin oder Korit angeboten wird. Komplett pyritisierte Ammoniten werden im Volksmund als Goldschnecken bezeichnet und ebenso wie auch geschnittene und polierte Exemplare, werden in Schmuckstücke (z. B. Amulette) eingearbeitet.
Das System der Ammoniten
Die Bezeichnung Ammonoidea umfasst neben den eigentlichen Ammoniten des Jura und der Kreide eine Reihe weiterer Formen, klassischerweise als Ordnungen geführt (Gattungslisten unvollständig):
Ammonoidea
 Perisphinctes im Berliner Museum für Naturkunde
Perisphinctes im Berliner Museum für Naturkunde-
- Ordnung Agoniatitida (Gattungen Agoniatites, Anarcestes, Maenioceras, Prolobites, Manticoceras, Beloceras)
- Ordnung Clymeniida ? frühe Goniatiten (Gattungen Acanthoclymenia, Gonioclymenia, Hexaclymenia, Wocklumeria, Platyclymenia, Clymenia, Parawocklumeria)
- Ordnung Goniatitida ? Echte Goniatiten (Gattungen Tornoceras, Cheiloceras, Sporadoceras, Gattendorfia, Ammonellipsites, Goniatites, Gastrioceras, Schistoceras, Perrinites, Cyclolobus)
- Ordnung Prolecanitida ? frühe Ceratiten (Gattungen Prolecanites, Medlicottia, Sageceras)
- Ordnung Ceratida ? Echte Ceratiten (Gattungen Xenodiscus, Otoceras, Beneckeia, Ceratites, Cloristoceras, Tropites, Cladiscites, Ptychides, Pinacoceras)
 Ein Exemplar von Hoploscaphites aus Nordamerika. Die Gehäuseform weicht von der einfachen Spirale ab.
Ein Exemplar von Hoploscaphites aus Nordamerika. Die Gehäuseform weicht von der einfachen Spirale ab.- Neoammonoidea
- Ordnung Phylloceratida (Gattungen Phylloceras, Leiophyllites)
- "Ordnung" Lytoceratida (Gattungen Lytoceras u. a.)
- "Ordnung" Ancyloceratida (Gattungen Ancyloceras, Macroscaphites, Crioceratites, Baculites, Turrilites, Bostrychceras, Scaphites, Hoploscaphites, Douvilleiceiras, Parahoplites, Deshayesites)
- Ordnung Ammonitida Echte Ammoniten (Mehrere Überfamilien)
- Psilocerataceae (Gattungen Psiloceras, Schlotheimia, Arietites, Echioceras, Oxynoticeras)
- Eoderocerataceae? ?Ringripper? (Gattungen Eoderoceras, Androgynoceras, Amaltheus, Pleuroceras, Dactylioceras)
- Hildocerataceae? ?Sichelripper? (Gattungen Harpoceras, Hildoceras, Leioceras, Ludwigia, Sonninia, Oppelia)
- Stephanocertaceae (Gattungen Stephanoceras, Macrocephalites, Kosmoceras, Quenstedtoceras)
- Perisphinctaceae (Gattungen Perisphinctes, Ataxioceras, Rasenia, Gravesia, Aulacostaphanus, Virgatites, Aspidoceras, Polyptychides)
- Weitere Gattungen (ehemals Überfamilien Desmocerataceae und Hoplitaceae): Callizoniceras, Pachydiscus, Leymeriella, Schloenbachia, Tissotia, Flickia.
Literatur
- U: Lehmann: Ammoniten. Ihr Leben und ihre Umwelt 2. Auflage. Verlag Enke, Stuttgart 1987. ISBN 3-432-88532-6. (gut geeignete Einführung, allerdings ohne systematischen Teil)
- A. H. Müller: Lehrbuch der Paläozoologie, Band II/2 (Invertebraten: Teil 2: Mollusca 2 – Arthropoda 1). 4. Auflage. Verlag Dr. Friedrich Pfeil, München 1994. ISBN 3-899-37017-1. (Ein Lehrbuch mit systematischem Teil, wer genaueres zu Arten erfahren möchte, ist auf speziellere Literatur angewiesen.)
- A. E. Richter: Ammoniten. Überlieferung, Formen, Entwicklung, Lebensweise, Systematik, Bestimmung Verlag Franckh-Kosmos, Stuttgart 1982. ISBN 3-811-21166-8 (Auch ein Überblick, eher für Sammler; der Band von Lehmann bietet mehr und genaueres zur Biologie.)
- R. Schlegelmilch: Die Ammoniten des süddeutschen Lias. Gustav Fischer Verlag, Stuttgart 1976. ISBN 3-437-30238-8
- R. Schlegelmilch: Die Ammoniten des süddeutschen Dogger. Gustav Fischer Verlag, Stuttgart 1985. ISBN 3-437-30488-7
- R. Schlegelmilch: Die Ammoniten des süddeutschen Malms. Gustav Fischer Verlag, Stuttgart 1994. ISBN 3437306103 (Drei Beispiele für Monografien der Ammoniten in einem geographischen Raum, in dem klassische Arbeiten zu Ammoniten entstanden. Hervorragende Zeichnungen und umfassende Beschreibungen, zahlreiche Abbildungen von Typusexemplaren. Standardwerke für die Bestimmung.)
- Zum Ober-Bathonium (Mittlerer Jura) im Raum Hildesheim, Nordwestdeutschland - Mega- und Mikropaläontologie, Biostratigraphie. In: Geologisches Jahrbuch Reihe A Heft 121. Hannover 1990. Seite 21–63.
- H. Keupp: Ammoniten. Paläobiologische Erfolgsspiralen Jan Thorbecke Verlag, Stuttgart 2000. ISBN 3-7995-9086-2. (Ein umfassendes Nachschlagewerk, welches zum Thema Ammoniten kaum eine Frage offen lässt.)
Weblinks








Wikimedia Foundation.