- Ribonucleotide Reductase
-
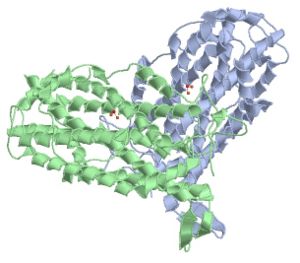 R2-Untereinheit der Class I-RNR
R2-Untereinheit der Class I-RNRDie Ribonukleotidreduktase (oft auch als RNR abgekürzt) ist ein Enzym, welches das letzte Glied in der Kette der Synthese der DNA-Bausteine bildet. Es reduziert die 2'-Hydroxylgruppe von Nukleotiden.
Inhaltsverzeichnis
Bedeutung und Vorkommen
Die Ribonukleotidreduktase ist ein unentbehrliches Enzym in der Biologie. Soweit bekannt, benutzt jeder Organismus die Ribonukleotidreduktase, um DNA-Bausteine herzustellen.
Da das Enzym die 2'-Hydroxy-Gruppe der Nukleotide reduziert, kann es auch als Bindeglied zwischen der DNA- und RNA-Welt gesehen werden. In diesem Zusammenhang wird auch spekuliert, ob RNR das Enzym war, welches den Weg aus einer primitiven RNA- zur heutigen DNA-Welt ebnete. (Siehe dazu auch: Chemische Evolution und Ribozym)
Wie viele Enzyme katalysiert die Ribonukleotidreduktase auch die entsprechende Rückreaktion (Oxidation von Desoxynukleotid zum Nukleotid). Diese Reaktion spielt für biologische Prozesse allerdings keine Rolle.Biologische Funktion
 Die katalysierte Reaktion der RNR
Die katalysierte Reaktion der RNRDie Ribonukleotidreduktase reduziert die Nukleotide in ihre jeweiligen Desoxynukleotide. Das Enzym unterscheidet hierbei nicht, um welches Nukleotid es sich handelt (Adenosin-, Guanosin-, Cytidin- oder Thymidin-Phosphate), sowohl Di- als auch die jeweiligen Triphosphate werden reduziert. Monophosphate und (phosphatfreie) Nukleoside werden nicht umgesetzt.
Ein herausragendes Merkmal der Ribonukleotidreduktase ist das radikalische Ablaufen des katalytischen Schrittes.[1] Dazu speichert das Enzym ein stabiles Radikal, welches bei jedem Turn-Over in die Bindungstasche transportiert werden muss. Nach der chemischen Reaktion wird das Radikal zurückgewonnen und wieder zur 'Speicherstelle' zurück transportiert. Dabei wird angenommen, dass das Elektron über mehrere Aminosäuren hüpft, die alle über Wasserstoffbrücken verbunden sind. Genauere Details sind hierzu aber noch nicht erforscht.
Es gibt mehrere Klassen der Ribonukleotidreduktase (Class I bis Class III). Obwohl alle Klassen die gleiche Reaktion katalysieren, sind sie strukturell sehr verschieden. Die Klassen sind nach der Art eingeteilt, wie das System das Radikal generiert. Jede Klasse teilt sich noch in weitere Unterklassen ein; dies soll aber der Einfachheit halber nicht ausgeführt werden.
Class I
Class I (EC 1.17.4.1) ist der Enzymtyp, welcher im Menschen vorkommt und am besten untersucht ist. Die Enzyme sind aerob.
Class I-RNR sind aus zwei verschiedenen Untereinheiten aufgebaut: R1 und R2.[2]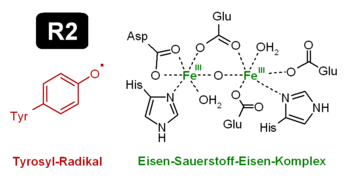 Molekulare Struktur des Tyrosylradikals mit dem benachbarten Eisen-Sauerstoff-Eisen-Komplexes, der das Radikal stabilisiert.
Molekulare Struktur des Tyrosylradikals mit dem benachbarten Eisen-Sauerstoff-Eisen-Komplexes, der das Radikal stabilisiert.R1 beinhaltet mehrere Bindungsstellen für Effektoren, sowie die Bindungstasche (active site), in der das Nukleotid reduziert wird. In R2 wird ein Radikal auf einer Tyrosinseitenkette gespeichert. Dieses Radikal hat eine erstaunlich hohe Halbwertszeit von etwa vier Tagen. R1 und R2 haben eine sehr geringe Affinität, so dass das Enzym selten als Gesamtkomplex vorliegt. Dies ist auch der Grund, warum es bisher noch nie gelang, die Ribonukleotidreduktase als Ganzes zu kristallisieren.
Für den katalytischen Schritt muss das Radikal vom Tyrosylradikal in R2 auf ein Cystein in R1 transportiert werden.[3] Der Abstand wird dabei auf 35 Ångström geschätzt. Es wird vermutet, dass das Elektron dabei über mehrere Aminosäuren 'hüpft', um die (für molekulare Verhältnisse) enorm weite Strecke zurück zu legen. Vergleicht man diese Strecke mit unseren Grössenverhältnissen, müsste ein Langstrecken-Läufer eine Distanz von 1050 Kilometer zurücklegen. Das entspricht dem 25fachen eines Marathons.
Class II
Class II-RNR (EC 1.17.4.2) kommen unter anderem im Organismus Lactobacillus leichmannii vor. Hier wird das Radikal in situ generiert, indem im Coenzym B12 die Cobalt-Kohlenstoff-Bindung gebrochen wird. Enzyme dieser Klasse sind fakultativ aerob. Sie können sowohl in An- als auch in Abwesenheit von Sauerstoff arbeiten.
Class III
Hier wird das Radikal auf einer Glycyl-Seitenkette gespeichert. Bakteriophage T4 zum Beispiel arbeitet mit dieser Klasse des Enzyms. Class III-RNR sind anaerob, das heißt sie arbeiten nur in einer sauerstofffreien Umgebung.
Mechanismus der Reduktion
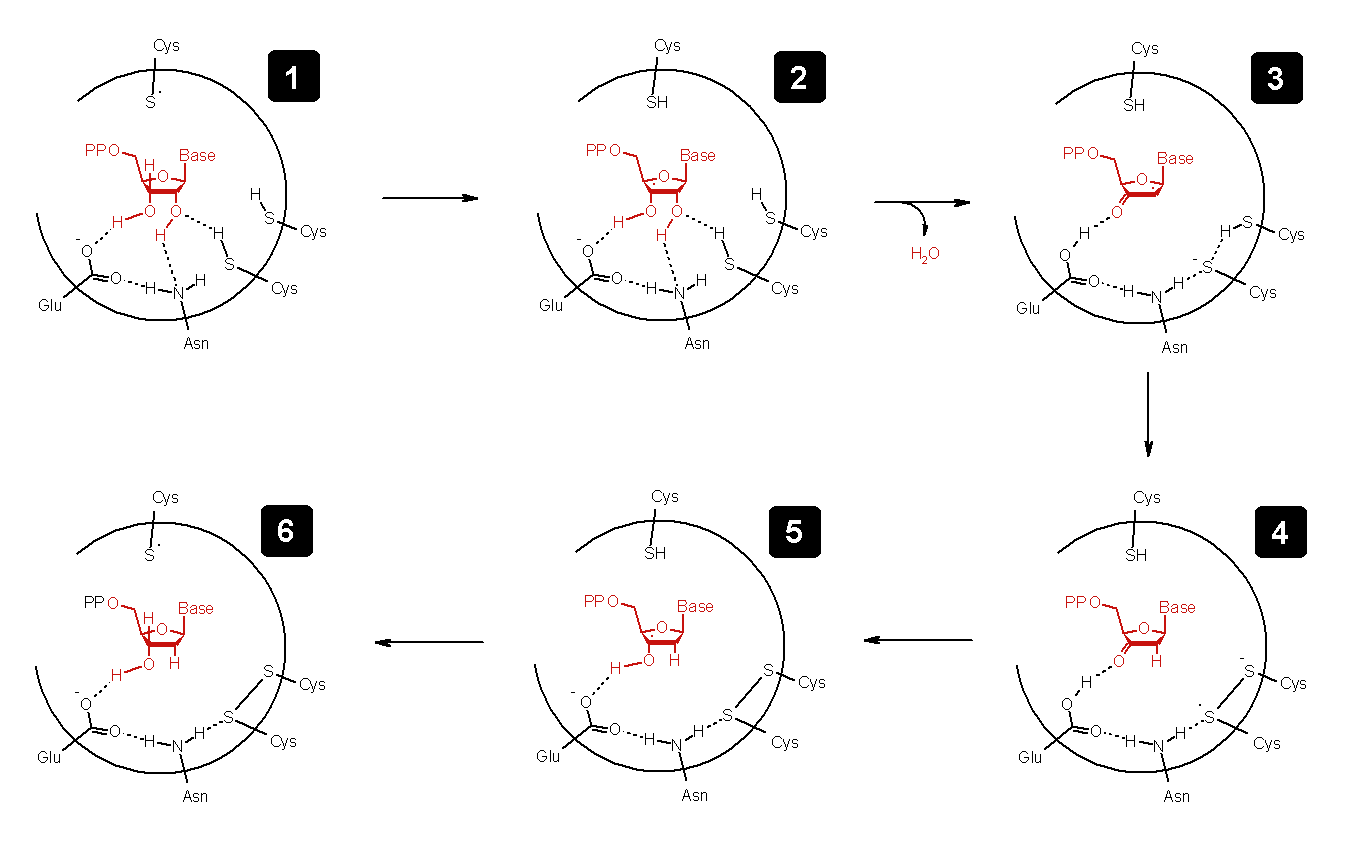
Der genaue Mechanismus der Nukleotid-Reduktion ist ziemlich kompliziert und bisher auch noch nicht vollständig aufgeklärt. Im folgenden Schema ist der Ablauf mit den vermuteten Zwischenschritten aufgeführt, wie er zur Zeit in der Wissenschaft diskutiert wird.
 Biochemischer Mechanismus der Reduktion eines Nukleotids
Biochemischer Mechanismus der Reduktion eines Nukleotids- Es wird zuerst ein Radikal auf die 3'-Position der Ribose übertragen (Schritt vom 1. zum 2. Bild).
- Nach Abspaltung von Wasser an der 2'-Position (Bild 2 -> 3)
- wird das Radikal auf zwei Cysteinketten des Enzyms übertragen (Bild 3 -> 4).
- Danach wird das Radikal über Ribose
- zurück auf das ursprüngliche Cystein übertragen (Bild 4 -> 6).
Insgesamt wird unter Oxidation von zwei Cystein-Seitenketten die Ribose zur Desoxyribose reduziert. Aus einem RNA- wurde ein DNA-Baustein hergestellt. Dieser Baustein ist nun bereit, um von der DNA-Polymerase in die DNA-Doppelhelix eingebaut zu werden.
Regulation
Da die RNR nur in speziellen Phasen der Zellteilung aktiv sein muss, gibt es wie bei den meisten Enzymen Mechanismen, um die RNR an- bzw. abzuschalten. Zudem wird in einem komplizierten System genau geregelt, welches der vier Nukleotide reduziert werden soll. Hier sollen nur kurz die Aktivatoren und Effektoren aufgezählt werden, ohne genauer auf die Details einzugehen:
Aktivatoren:
- ATP aktiviert das Enzym
- dATP deaktiviert es
Effektoren:
- ATP und dATP -> CDP/CTP und UDP/UTP werden reduziert
- dGTP -> ADP/ATP
- dTTP -> GDP/GTP
Entdeckung der Ribonukleotidreduktase
Spätestens seit der Entdeckung der Struktur der DNA durch James Watson und Francis Crick im Jahr 1953 stellte sich die Frage, wie die Zelle die einzelnen Bausteine für das DNA-Polymer herstellt. Acht Jahre später wurde zum ersten mal aus verschiedenen Zellen eine Substanz isoliert, die fähig war, Nukleotide in ihre entsprechenden Desoxynukleotide zu reduzieren.[4] Nach und nach konnte ein immer vollständigeres Bild des neuentdeckten Enzyms gezeichnet werden. Man fand heraus, dass sich die Ribonukleotidreduktase aus zwei verschiedenen Untereinheiten zusammensetzt, und entdeckte die Effektoren und Aktivatoren der RNR.
Es dauerte jedoch noch bis in die 1990er Jahren, bis man die beiden Untereinheiten des Enzyms einzeln kristallisieren konnte und somit die molekulare Struktur herausgefunden hatte.[5][6] Nach Aufklärung der Struktur stellen sich nun weitere Fragen, so etwa nach dem Mechanismus der Andockung der beiden Untereinheiten R1 und R2 aneinander, dem Radikaltransport durch das Enzym über eine verhältnismäßig große Distanz hinweg bzw. nach der Generierung des Radikals bei der Herstellung des Enzyms.
Man ist in den letzten Jahren in diesen Fragen nicht sehr viel weiter gekommen. Es gibt zwar etliche plausible Modelle, wie die RNR auf atomarer Ebene arbeitet, aber wenig gesicherte Tatsachen.
Die RNR in der Krebsforschung und Krebstherapie
Die Ribonukleotidreduktase steht auch im Fokus der Krebsforschung. Weil das Enzym immer dann gebraucht wird, wenn die Zelle sich teilt oder DNA-Schäden reparieren muss, ist die Zelle beim Wachstum auf die RNR angewiesen. Das Enzym ist mit einer Stoffwechsel-Rate von ca. 10 s-1 sehr langsam, sprich ineffizient. Das ist aufgrund der Teilungsrate der normalen Zelle auch nicht problematisch, für Krebszellen ist dies jedoch ein Hindernis, weil diese möglichst schnell zu wuchern versuchen. Es gibt deshalb Krebszellen, die die RNR derart modifizieren, dass es schneller arbeiten kann. Ein Wirkstoff, der genau diese modifizierten Enzyme blockiert, würde den Krebs im Wachstum stark verlangsamen oder im optimalen Fall sogar stoppen.[7]
In der Behandlung von myeloischen Leukämien (vor allem bei Anzeichen einer Leukostase) und anderen myeloproliferativen Erkrankungen wie essentielle Thrombozythämie und Polycythaemia vera (rubra) wird Hydroxycarbamid (z.B. Syrea®, Litalir®), ein Hemmstoff der Ribonukleotid-Reduktase, als Chemotherapeutikum eingesetzt.
Quellen und weiterführende Informationen
Einzelnachweise
- ↑ JoAnne Stubbe, Daniel G. Nocera et al., Radical Initiation in the Class I Ribonucleotide Reductase: Long-Range Proton-Coupled Electron Transfer?, Chemical Reviews, 2003, 103, p. 2167-2201.
- ↑ Ulla Uhlin, Hans Eklund, Structure of ribonucleotide reductase protein R1, Nature, vol. 370, 18. August 1994, p.533-539.
- ↑ Britt-Marie Sjöberg et al., Two Conserved Tyrosine Residues in R1 Participate in an Intermolecular Electron Transfer in Ribonucleotide Reductase, J. of Biol. Chem., 1996, vol. 271, No. 34, p. 20655-20659.
- ↑ Peter Reichard, Astor Baldesten and Lars Rutberg, Formation of Deoxycytidine Phosphates from Cytidine Phosphates in Extracts from Escherichia coli, J. of Biol. Chem., 1961, vol. 236, No. 4, p. 1150-1157.
- ↑ Nordlund, P., Sjöberg B.-M., Eklund, H., Three-dimensional structure of the free radical protein of ribonucleotide reductase, Nature, 1990, vol. 345, p. 593-598.
- ↑ Ulla Uhlin, Hans Eklund, Structure of ribonucleotide reductase protein R1, Nature, 1994, vol. 370, p. 533-539.
- ↑ Yun Yen, Ribonucleotide Reductase Subunit One as Gene Therapy Target, Clinical Cancer Research, vol. 9, 2003, p.4304-4308.
Literatur
- P. J. Sadler: Metal Sites in Proteins and Models - Iron Centres, Structure and Bonding Vol. 88, Springer Verlag, ISBN 3-540-62870-3.
Weblinks


Wikimedia Foundation.