- aDNA
-
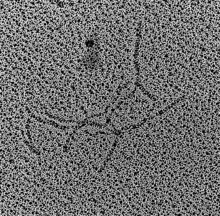 Quervernetzte aDNA aus einer 4000 Jahre alten Niere eines ägyptischen Priesters mit Namen Nekht-Ankh (vergrößert)
Quervernetzte aDNA aus einer 4000 Jahre alten Niere eines ägyptischen Priesters mit Namen Nekht-Ankh (vergrößert)
aDNA stammt aus dem englischen „ancient DNA“ (alte DNA) und ist die technische Bezeichnung für Reste von Erbgutmolekülen in toten Organismen.
Laut Konvention handelt es sich um aDNA, wenn die Probe von einem toten Organismus stammt und keine direkten Verwandten des beprobten Organismus mehr leben. Zum Beispiel wird menschliche DNA dann als aDNA angesprochen, wenn das Individuum mindestens 75 Jahre vor der Probenuntersuchung verstorben ist.
Außerdem wird auch derjenige Forschungszweig der Molekularbiologie mit „aDNA“ bezeichnet, der die alten Erbmoleküle – eben die aDNA – erforscht.
Die aDNA-Forschung ist in Zielen und Methoden eng mit der genetischen Rechtsmedizin und der forensischen Anthropologie (s. Anthropologie) verwandt.
Inhaltsverzeichnis
Geschichte
Die Geschichte der aDNA ist eng mit der Entwicklung der Polymerase-Kettenreaktion (engl. Polymerase Chain Reaction, PCR), einer speziellen molekularbiologischen Technik verwoben, die es ermöglicht, auch geringe Mengen Erbgut zu vervielfältigen und zu untersuchen. In der Anfangszeit der aDNA kam es zu einer Reihe von Berichten über aDNA, die sich später als falsch herausstellten und Material verwendeten, das aus heutiger Sicht keine Aussicht auf Erhaltung von amplifizierbarer DNA hat. Den Ergebnisse der Untersuchung von aDNA wird daher auch heute noch mit Vorbehalten begegnet.
Anwendungsgebiete
Der Erkenntnisgewinn anhand von aDNA ist für die genetische Biologie, die Paläozoologie, die Paläobotanik sowie die Anthropologie (besonders Paläoanthropologie) und mit letzterer im Schulterschluss auch für die Archäologie von Bedeutung. Aus letzterer entwickelte sich ein eigener Wissenschaftszweig, die Paläogenetik.
Artbestimmung
Jede biologische Art ist durch ein spezifisches genetisches Merkmalsmuster gekennzeichnet, deswegen ermöglicht die Untersuchung des Erbgutes schon einer einzigen Zelle die eindeutige artspezifische Zuweisung. aDNA nutzt man zur Artbestimmung, wenn die Erhaltungsbedingungen keine anderen eindeutigen Identifikationsmöglichkeiten mehr zulassen oder die Trennschärfe anderer Methoden zu ungenau ist. So lassen sich zum Beispiel Schafe und Ziegen aufgrund hoher Ähnlichkeit im Knochenbau allein anhand der Skelettmerkmale nicht auseinanderhalten.
Phylogenese
Die Evolution der Arten und ihre verwandtschaftlichen Beziehungen untereinander lassen sich statistisch anhand ihrer verschiedenen genetischen Merkmalsmuster darstellen; als Maßzahl dient hier der genetische Abstand. Zur Einordnung bereits ausgestorbener Arten behilft sich die phylogenetische Forschung der aDNA.
Individuelle Verwandtschaftsanalyse
Zwei Individuen sind im biologischen Sinne miteinander verwandt, wenn sie mindestens einen gemeinsamen Vorfahren aufweisen. Der Grad der biologischen Verwandtschaft zweier Individuen lässt sich ebenfalls anhand ihres Erbgutes ablesen. Verfahren ähnlich dem genetischen Vaterschaftstest finden auch in der aDNA-Analyse Anwendung. Einschränkend wirkt sich hier allerdings die sehr bruchstückhafte Erhaltung der DNA-Moleküle aus, die die Untersuchung jeweils nur kleiner Abschnitte ermöglicht. Besonderes Augenmerk gilt hier den Bereichen hoher Variabilität, d. h. Stellen, an denen häufig Mutationen auftreten. Im speziellen sind das STRs (Short Tandem Repeats) und in zunehmenden Maße auch SNPs (Single Nucleotide Polymorphisms).
Vielversprechend ist hier v.a. die Auswertung mitochondrialer DNA, da diese im Vergleich zur DNA des Zellkerns in wesentlich größerer Kopienzahl vorliegt (etwa 1000 mitochondriale Kopien, aber nur 2 nukleare pro Zelle) und so Erhaltungsprobleme gelindert werden. Allerdings werden Mitochondrien nur von der Mutter, nicht aber vom Vater an die Kinder weitergegeben, d. h. es lässt sich nur die – im biologischen Sinne – matrilineare Abstammungslinie verfolgen.
Geschlechtsdiagnostik
Das genetische Geschlecht eines Individuums ist bei günstigen Erhaltungsbedingungen, d. h. bei der Überlieferung chromosomaler DNA, bestimmbar. Bei Arten, die wie der Mensch nur ein geschlechtsspezifisches Chromosom (Y-Chromosom) besitzen, sind sichere Differenzierungsmöglichkeiten jedoch eingeschränkt. Mit dem Nachweis von aDNA-Bruchstücken, die vom Y-Chromosom stammen (zum Beispiel aus der SRY-Region) ist das genetische Geschlecht eindeutig als männlich belegt, dagegen kann aus dem Fehlen Y-spezifischer Merkmale nicht sicher auf weibliches Geschlecht geschlossen werden, denn unwägbare Erhaltungsbedingungen könnten ebenso der Grund hierfür sein.
Trotz dieser Unsicherheit wird die molekulare Geschlechtsdiagnostik v.a. bei menschlichen Skelettresten angewendet. Insbesondere bei Individuen, an denen aufgrund des jungen Sterbealters oder unspezifischer Skelettmerkmale nur eine unsichere Einordnung mittels biologisch-anthropologischer Methoden möglich ist, hilft die aDNA weiter.
Identifizierung von Individuen und Gegenständen
Als Ausnahmen, die allerdings meist unter großer Anteilnahme der Öffentlichkeit stattfinden, können die Identifizierungen historisch bedeutender Personen mit sterblichen Überresten gelten. Der Wissenschaftler extrahiert dazu zunächst aDNA aus Gewebeproben des fraglichen Individuums (Knochen, Haare, Kleidung), vor allem um daraus die mt-Haplogruppe bzw. bei männlichen Individuen auch den Y-Haplotypen zu bestimmen. Anschließend vergleicht er diesen mit „authentischer“ DNA, die bestenfalls von noch lebenden mutmaßlichen Verwandten stammt. In manchen Fällen hilft auch die Rekonstruktion des genetischen Fingerabdrucks der Person, um diesen mit dem von eng verwandten Personen aus gesicherten Bestattungen zu vergleichen. Eine Extraktion aus persönlichen Gegenständen des fraglichen Individuums ist ebenfalls möglich, allerdings können hier aus Zweifeln an der Authentizität Probleme in der Beweisführung folgen. Bei signifikanter Übereinstimmung zwischen fraglichen und authentischen Proben gilt das Individuum als identifiziert.
Paläopathologie
Die wenigsten Krankheiten lassen sich eindeutig an Skeletten diagnostizieren, deswegen versucht die aDNA-Forschung seit zirka Mitte der 1990er Jahre Erreger von Infektionskrankheiten in menschlichen Überresten nachzuweisen. Im Mittelpunkt stehen dabei zunächst neben der Methodenentwicklung unter anderem der Nachweis eventuell ausgestorbener infektiöser Bakterienstämme und die Gegenüberstellung geschichtlich überlieferter Krankheitsverläufe und -symptome mit den heutigen Erkenntnissen über die jeweilige Krankheit. Mit wachsender Datenzahl kann dieser Zweig in der Zukunft außerdem einen wichtigen Beitrag zur historischen Epidemiologie liefern.
Überlieferungsbedingungen und Probleme
In trockener und kühler Umgebung kann DNA lange Zeit überdauern, allerdings lösen sich die empfindlichen Makromoleküle in kleine Kettenbruchstücke auf. Wärme, Feuchtigkeit, saure und basische pH-Werte begünstigen diesen Zerfallsprozess in immer kleinere Stückchen. Als Faustregel gilt, dass aDNA hauptsächlich Nukleotid-Ketten enthält, die kürzer als 200 Basenpaare sind – das ist im Vergleich zur theoretischen Gesamtlänge des zum Beispiel menschlichen Genoms von 3×109 Basenpaaren, sehr gering[1]. Für gegebene Umweltparameter lassen sich „Halbwertszeiten“ berechnen, mit denen die zu erwartende Qualität der Ergebnisse abgeschätzt werden kann[2].
Da aDNA meist in sehr geringen Mengen überliefert ist, bedient sich die Forschung der PCR, um die erhaltenen Stücken zunächst zu vervielfältigen. Aufgrund der ausgesprochen hohen Sensibilität der PCR sind Fehlamplifikationen sehr häufig, d. h. es werden anstatt der originären alten Ziel-DNA entweder das Erbgut anderer Organismen (häufig von Boden-Bakterien) oder moderne DNA des Ziel-Organismus vervielfältigt, die durch ungenügende Aufbereitung des Materials oder unsauberes Arbeiten in die Probe gelangt ist.
Ein weiteres, bisher kaum erforschtes Problem liegt in den so genannten „Hot Spots“, denn diese DNA-Stellen können nach dem Ableben des Organismus durch chemische Reaktionen derart verändert werden. So entstehen Pseudo-Mutationen, die es zu erkennen gilt.
Von geringerer Bedeutung sind sogenannte Inhibitoren, die aus dem Liegenmilieu der DNA, zum Beispiel dem Boden, stammen und die PCR durch Blockieren des Enzyms verhindern können. Häufig wird Bovines Serumalbumin, ein aus Rinderblut gewonnenes Eiweiß, zur Bindung von Eisen, dem häufigsten Inhibitor, dem Reaktionsgemisch zugefügt. Liegen dennoch Hinweise vor, dass PCR-Inhibition die Ursache für falsch negative Ergebnisse ist, hilft es, die aDNA-Probe verdünnt einzusetzen. Leider verringert sich durch die Verdünnung auch die DNA-Konzentration in der Probe, was die Chancen auf PCR-Erfolg wieder verringert.
Schließlich sagt die sichtbare Erhaltung eines Organismus wenig über den Zustand der enthaltenen aDNA aus. So ist zum Beispiel aus Moorleichen aufgrund des sauren Liegemilieus selten verwertbare DNA zu extrahieren. Auch Trockenmumien mit hervorragender Weichteilerhaltung enthalten oftmals nur noch sehr geringe aDNA Spuren.
Verfahren
Die Methodik ist auf die Gewinnung möglichst reiner, vieler und großer Oligonukleotid-Ketten ausgerichtet. Je nach Überlieferungsbedingungen stehen verschiedene Gewebe zu Verfügung, jedoch sind aufgrund der Abgeschlossenheit Hartgewebe (Knochen, Dentin) vorzuziehen, in denen die DNA von zum Beispiel Osteoklasten am besten vor Umwelteinflüssen geschützt erhalten bleibt.
Nach der Gewinnung und Reinigung der DNA-Reste aus dem Gewebe werden aus diesen zuerst die zu untersuchenden Sequenzen in einer PCR bis zur Nachweisgrenze vervielfältigt. Die PCR-Amplifikate können dann durch Fragmentgrößenbestimmung oder Sequenzierung untersucht werden. Vor der Sequenzierung wird, um Kontamination oder bestimmte PCR-Artefakte aufzudecken, häufig das PCR Amplifikat kloniert und selektiert.
Berühmte und wichtige Beispiele
Die vorgestellte Auswahl soll einen Überblick über die Spannweite der aDNA-Forschung geben. Da auch negative Ergebnisse von wissenschaftlicher Bedeutung sind, werden „berühmte und wichtige Misserfolge“ im Folgenden ebenfalls angeführt.
Aus der Erforschung von ausschließlich moderner DNA – also dem Erbgut noch lebender Individuen – können ebenfalls Rückschlüsse auf vergangene Zusammenhänge und Entwicklungen gezogen werden. Ein Beispiel dafür ist die mitochondriale Eva.Prokaryoten
Eine wichtige Frage in der Paläopathologie, dem Streit um die Herkunft der Syphilis-Erreger, wurde unter anderem mittels aDNA-Analysen zu beantworten versucht. Die gezielte Suche nach allen bekannten Treponema-Erregern in einer 46 Skelette umfassenden Studie blieb jedoch erfolglos. Die Entdeckung von alten Tuberkulose-Bakterien in einigen Individuen derselben Serie bestätigte dagegen die allgemeine Nachweismöglichkeit von Erregern bestimmter Krankheiten[3].
Pflanzen
Tiere
Ancient DNA wurde in einer Vielzahl von Arbeiten zur Klärung von Verwandtschaftsverhältnissen bei Tieren verwendet, im Folgenden findet sich eine Auswahl der bearbeiteten Taxa.
- Die DNA aus den Zellen eines fossilen Höhlenbären aus der Vindija-Höhle gehörte 1999 zu den ersten in Teilen rekonstruierten Fragmenten ausgestorbener Lebewesen.[4]
- Der DNA-Code für das Hämoglobin eines 43.000 Jahre alten Wollhaarmammuts weist drei vom Hämoglobin eines Asiatischen Elefanten abweichende Sequenzen auf. Diese wurden 2010 in die DNA-Sequenz für das Hämoglobin eines Asiatischen Elefanten eingebaut, um Erkenntnisse über die Kälteanpassung der Mammuts zu gewinnen.[5]
- Beutelwolf
- Harpagornis (Haast-Adler)
- Mammut
- Mastodon
- Megaloceros (Riesenhirsch)
- Moas
- Quagga
- Fossilien in Bernstein
- Film und Roman des Titels „Jurassic Park“ haben Anfang der 1990er Jahre stark zur öffentlichen und sogar wissenschaftlichen Euphorie in Sachen aDNA beigetragen. In der Geschichte wird aus Fossilien in Bernstein (sog. Inklusen) altes Erbgut gewonnen, das zur Neuzüchtung bereits ausgestorbener Arten verwendet wird. Diese aufgrund dessen wohl bekannteste Methode der aDNA-Gewinnung ist allerdings nicht möglich – die Fossilien scheinen schlichtweg zu alt, um intakte Nucleotidketten, egal ob tierischen oder pflanzlichen Ursprung, zu enthalten. Bis heute ist kein reproduzierbarer positiver Nachweis von aDNA aus Bernstein gelungen.
- Riesenschildkröten auf den Maskarenen
- Rinder
- Säbelzahnkatzen
- Zwergelefanten auf griechischen Inseln
Menschen
aDNA-Forschung ist aufgrund ihres jungen Alters und der ungewohnt tiefen Einblicke, die sie in vergangene Zeiten zu liefern vermag, sehr medienwirksam. Vor allem Ergebnisse, die an menschlichen Überresten gewonnen wurden, tauchen hin und wieder als „Sensationsmeldungen“ in Presse und Rundfunk auf, zum Teil auch in die Pietät verletzender Form. Als Beispiel sei hier die Erforschung von Krankheiten historisch bekannter Personen genannt. Zu den Ergebnissen, die einer breiten Öffentlichkeit bekannt wurden, zählen die folgenden Beispiele:
- Ägyptische Mumien
-
- Aufgrund hervorragender Weichteilerhaltung sind die ägyptischen Mumien häufiger Gegenstand von aDNA-Untersuchung, denn man erhofft sich auch auf molekularer Ebene gute Überlieferungsbedingungen. Dagegen sprechen allerdings die hohen, den DNA-Zerfall fördernden, Temperaturen.
- Der bekannte Evolutionsgenetiker Svante Pääbo publizierte 1985 als erster die Entdeckung alter DNA in Proben einer ägyptischen Mumie.[6] Aufgrund damals noch fehlender Kontrollmöglichkeiten räumte er später jedoch, dass seine aDNA-Proben möglicherweise durch moderne DNA kontaminiert gewesen sein könnten. [7][8]
- russische Zarenfamilie (siehe Nikolaus II.)
- Neandertaler
-
- In der Arbeitsgruppe von Svante Pääbo wurden erstmals 1997 Teile der DNA [9], 2008 die vollständige mitochondriale DNA [10] und 2010 mehr als drei Milliarden Basenpaare (= 60 Prozent) der DNA analysiert.[11]
-
- Der Tod Bormanns (der sich später bei Untersuchungen des Skeletts als Freitod herausstellte) im Frühjahr 1945 ist mehrfach angezweifelt worden. 1972 wurden am Lehrter Bahnhof in Berlin zwei Skelette entdeckt, von denen eines laut gerichtsmedizinischem Gutachten (Gebissmerkmale) als Bormann identifiziert werden konnte. Erneute Zweifel führten schließlich zu einer DNA-Analyse Ende der 1990er – obwohl es sich der Definition nach noch nicht um eine aDNA-Untersuchung handelte (<75 Jahre), konnten nur noch verhältnismäßig geringe Reste DNA nachgewiesen werden. Die Übereinstimmung mitochondrialer Muster zwischen dem beprobten Skelett und einer noch lebenden Cousine Bormanns macht eine verwandtschaftliche Beziehung in mütterlicher Linie und damit die Identifizierung Bormanns wahrscheinlich.[12]
-
- Der Schwedenkönig fiel 1632 während der Schlacht bei Lützen. Seine Leiche wurde einbalsamiert nach Stockholm überführt und dort beigesetzt. Teile seiner Kleidung verblieben jedoch in Lützen und werden dort heute im Museum ausgestellt. Die Untersuchung von Blutresten im Stoff erbrachte ausreichende Mengen alter DNA, um diese mit dem Erbgut seiner Nachfahren im heutigen schwedischen Herrscherhaus vergleichen zu können. Die Echtheit wurde bestätigt.[13]
- Thomas Jefferson
- Friedrich Schiller
- Verwandtschaftsanalyse der spätbronzezeitlichen Skelette aus der Lichtensteinhöhle im Harz
- Verwandtschaftsanalyse der schnurkeramischen Skelette aus Eulau in Sachsen-Anhalt (Haak et al 2008)
Siehe auch
Literatur
Zur Einführung
- M. Jones: The Molecule Hunt. How Archaeologists are Bringing the Past Back to Life. Penguin Books 2001–2, ISBN 1-55970-679-1. – Guter Abriss der Forschungsgeschichte und nebenbei allgemein verständliche Erklärung der Methoden; zuweilen etwas zu ungenau. Der Autor ist einer der Vorreiter auf dem Gebiet der aDNA.
- S. Hummel: Ancient DNA Typing. Methods, Strategies and Applications. Springer, 2003, ISBN 3-540-43037-7.
- Michael Hofreiter: Spurensuche in alter DNA. Biologie in unserer Zeit 39(3), S. 176 – 184 (2009), ISSN 0045-205X pdf online
Fachartikel
- Jermey J. Austin, Andrew J. Ross, Andrew B. Smith, Richard A. Fortey, Richard H. Thomas: Problems of reproducibility – does geologically ancient DNA survive in amber-preserved insects?. In: Proceedings of the Royal Society London. 264/1997. S. 467–474. (PDF)
- Abigail S. Bouwman, Terence A. Brown: The limits of biomolecular palaeopathology: ancient DNA cannot be used to study venereal syphilis. In: Journal of Archaeological Science. 32/2005. S. 703–713.
- W. Haak, G. Brandt, H. de Jong et al.: Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age. doi:10.1073/pnas.0807592105
- Matthias Krings, Anne Stone, Ralf W. Schmitz, Helke Krainitzki, Mark Stoneking, Svante Pääbo: Neandertal DNA Sequences and the Origin of Modern Humans. In: Cell. 90/1997. S. 19–30. (PDF)
- Isolina Marota, Corrado Basile, Massimo Ubaldi, Franco Rollo: DNA decay rate in papyri and human remains from Egyptian archaeological sites. In: American Journal of Physical Anthropology. 117/2002. S. 310–318.
Einzelnachweise
- ↑ Hummel 2003
- ↑ zum Beispiel Marota et al 2002
- ↑ Bouwman & Brown 2004
- ↑ A. D. Greenwood u. a.: Nuclear DNA sequences from late Pleistocene megafauna. In: Molecular Biology and Evolution, Band 16, 1999, S. 1466–1473, Volltext (PDF)
- ↑ Kevin L. Campbell u. a.: Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance. In: Nature Genetics, Band 42, 2010, S. 536–540, doi:10.1038/ng.574
- ↑ Svante Pääbo: Molecular cloning of ancient Egyptian mummy DNA. In: Nature, Band 314, Nr. 6012, 1985, S. 644–645, doi:10.1038/314644a0
- ↑ plosgenetics.org vom 28. März 2008: J. Gitschier (2008): Imagine: An Interview with Svante Pääbo. In: PLoS Genet 4(3): e1000035. doi:10.1371/journal.pgen.1000035
- ↑ Nick Zagorski: Profile of Svante Pääbo. In: PNAS, Band 103, Nr. 37, 2006, S. 13575–13577, doi:10.1073/pnas.0606596103 und als [www.ncbi.nlm.nih.gov/pmc/articles/PMC1564240/pdf/zpq13575.pdf Volltext (PDF)]
- ↑ Matthias Krings u. a.: Neandertal DNA Sequences and the Origin of Modern Humans. In: Cell, Band 90, Nr. 1, 1997, S. 19–30, doi:10.1016/S0092-8674(00)80310-4
- ↑ Richard E. Green u. a.: A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. In: Cell, Band 134, Nr. 3, 2008, S. 416–426, doi:10.1016/j.cell.2008.06.021
- ↑ Richard E. Green u. a.: A draft sequence of the Neandertal Genome. In: Science, Band 328, 2010, S.710–722, doi:10.1126/science.1188021
- ↑ K. Anslinger, G. Weichhold, W. Keil, B. Bayer, W. Eisenmenger: Identification of the skeletal remains of Martin Bormann by mtDNA analysis. In: International Journal of Legal Medicine. 114/2001. S. 194–196.
- ↑ H. Ellegren: Gamla gener guldgruva för historiker. In: Forskning & Framsteg. 6/1994. S. 21–26
Wikimedia Foundation.