- Luziferin
-
Luciferine sind unterschiedliche Naturstoffe, die von verschiedenen Organismen synthetisiert werden. Sowohl die Luciferine als auch die Luciferasen sind art- oder gruppenspezifisch, also für jede Tiergruppe kennzeichnend. In Gegenwart des entsprechenden Luciferase-Enzyms reagieren sie mit Sauerstoff (Oxidation). Bei der Veränderung, meistens der Abspaltung von Teilgruppen an dem Luciferin, entsteht Energie, die als Lichtquant abgegeben wird.
Inhaltsverzeichnis
Geschichte und Allgemeines
Die ersten Arbeiten auf Luciferin-Luciferase-Systeme gehen auf den Franzosen Raphael Dubois zurück. Er entdeckte bei Arbeiten an Leuchtkäfern 1885, dass eine Substanz in einer lichtgebenden Reaktion verbraucht wird. Er bezeichnete diese als Luciferin, welche auch durch Hitze nicht zerstört wird. Die andere, hitzelabile Komponente hat der Wissenschaftler als Luciferase bezeichnet. Heutzutage wird Luciferase allgemein als das Enzym bezeichnet, dass das dazugehörende Luciferin umsetzt.
Die nächsten Untersuchungen wurden von Amerikaner Newton Harvey Anfang des 20. Jahrhunderts durchgeführt.[1] Er hat herausgefunden, dass es in jedem Luciferin-Luciferase-System eine Spezifität gibt. So können Luciferine der einen Spezies nicht durch die Luciferase einer artfremden Spezies umgesetzt werden. Schließlich benötigt jedes biolumineszente System Sauerstoff. Dies wurde aber bereits im 18. Jahrhundert von Robert Boyle beobachtet.
Biolumiszente Systeme sind nicht evolutionär konserviert, die Luciferasen teilen keine Sequenzhomologie. Luciferasen treten aber in 17 unterschiedlichen Stämmen und mindestens 700 Gattungen - meist marinen - auf.[2] Offenbar wurden sie öfters „erfunden“, phylogenetische Studien wiesen darauf hin, dass Luciferin-Luciferase-Systeme mehr als 30 unabhängige Ursprünge besitzen.[3]
Laut klassischer Definition ist das an der Luciferase gebundene Luciferin der Lichtemitter. Die Luciferase setzt unter Verbrauch von Sauerstoff das Luciferin hierbei um, manchmal werden dafür auch Cofaktoren wie ATP oder Ionen benötigt. Das oxidierte Luciferin befindet sich zunächst in einem Übergangszustand I und gelangt dann - häufig nach Decarboxylierung und weiteren Zwischenschritten - in einen elektronisch angeregten Zustand P*. Dieser fällt nach recht kurzer Zeit (wenige Nanosekunden) zurück in seinen Grundzustand P und emittiert währenddessen einen Lichtquant. Normalerweise sind die umgesetzten Luciferine auch Fluorophore, da sie durch Bestrahlen von Licht in einen angeregten Zustand gelangen können.
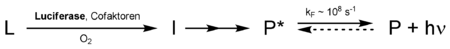 Luciferine werden durch Luciferasen unter Verbrauch von Sauerstoff umgesetzt, dabei entsteht ein Molekül in einem elektronisch angeregten Zustand P*. Nach einer kurzen Lebenszeit wird ein Photon emittiert und der Grundzustand P erreicht.
Luciferine werden durch Luciferasen unter Verbrauch von Sauerstoff umgesetzt, dabei entsteht ein Molekül in einem elektronisch angeregten Zustand P*. Nach einer kurzen Lebenszeit wird ein Photon emittiert und der Grundzustand P erreicht.Prinzipien
Um in den angeregten Zustand P* zu gelangen, ist biochemisch gesehen viel Energie nötig. Die Emission von Photonen mit einer Wellenlänge von 500 nm (grün, Energie etwa 2 eV/Photon) benötigt ca. 250 kJ/mol – zum Vergleich: die Hydrolyse von ATP zu ADP und Phosphat setzt ca. 30 kJ/mol frei. Außerdem kann die Energie nur in einem Schritt freigesetzt werden.
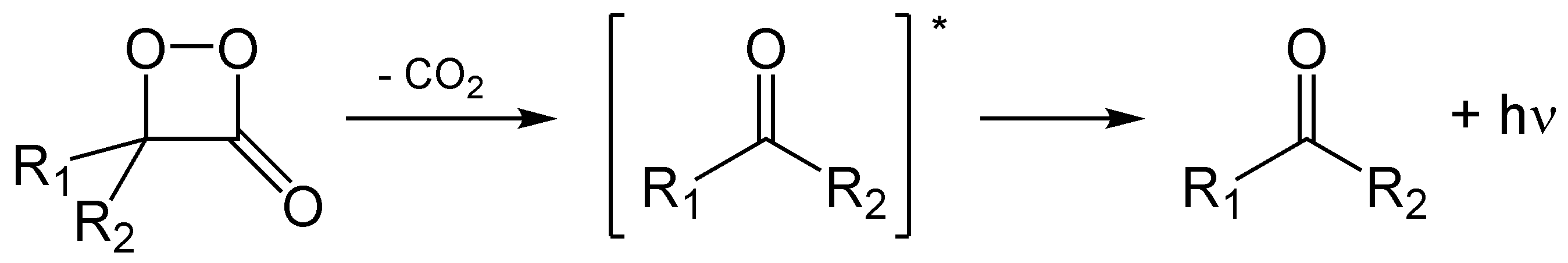
Das häufigste Prinzip ist die Generierung eines Vierrings, einem Dioxetan bzw. Dioxetanon. Nach erfolgter Decarboxylierung bildet sich der elektronisch angeregten Zustand.
 Ein Dioxetanon ist instabil und zerfällt unter Abspaltung von CO2. Dies erzeugt ein Keton im elektronisch angeregten Zustand.
Ein Dioxetanon ist instabil und zerfällt unter Abspaltung von CO2. Dies erzeugt ein Keton im elektronisch angeregten Zustand.Manchmal entspricht die Fluoreszenz nicht der erwarteten, zum Beispiel bei Studien in vitro (im Reagenzglas). Dafür gibt es verschiedene Ursachen. So emittieren an Enzyme gebundene Luciferine anders als freie Luciferine nach Anregung durch Licht. Manchmal wird die Energie an einen zweiten Fluorophor übertragen, so wie es beispielsweise bei Aequorin zu GFP in A. victoria geschieht.
Quantenausbeute Q
Ob die Umsetzung eines Luciferins durch die korrespondierende Luciferase effizient ist, wird durch die sogenannte Quantenausbeute Q (quantum yield) determiniert. Sie ist definiert als die Anzahl der emittierten Lichtquanten pro dafür umgesetzte Moleküle Luciferine.[4] Definitionsbedingt ist der Höchstwert von Q gleich 1, dies würde nämlich bedeuten, dass bei jedem umgesetzten Molekül Luciferin auch ein Lichtquant frei wird.
Luciferin-Arten
Luciferin-Luciferase-Systeme kommen in zahlreichen Arten vor. Es gibt vier Hauptklassen an Luciferin-Luciferase-Systemen, bei denen das Luciferin nach Umsetzung durch eine Luciferase in einen elektronisch angeregten Zustand überführt und damit der eigentliche Lichtemitter wird.
Das Firefly-Luciferin, ein Benzothiazolen

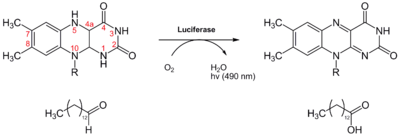
Die meisten biolumineszenten Insekten sind Käfer (Coleoptera) in den Familien Phengodidae (Federleuchtkäfer) und Elateridae (Schnellkäfer, Leuchtkäfer). Das Luciferin-Luciferase-System des nordamerikanischen Leuchtkäfers Photinus pyralis (engl. firefly) ist dabei am besten untersucht. Die Luciferase (EC 1.13.12.7 [6]), ein ca. 60 kD großes Enzym, katalysiert die oxidative Decarboxylierung von D-Luciferin (ein Benzothiazol) zu Oxyluciferin (vgl. untenstehende Abbildung, Kasten). Dafür benötigt sie neben Magnesiumionen auch ATP als Cofaktor. Die Reaktion findet in der Peroxisomen der Lichtorganzellen statt.[7]
Die Lichtausbeute dieser Reaktion ist außerordentlich hoch, da die Quantenausbeute Q bei 41 % liegt.[8] Je nach Art liegt das Emissionsmaximum λmax des freigesetzten Lichts zwischen 530 nm (grün) und 635 nm (rot).
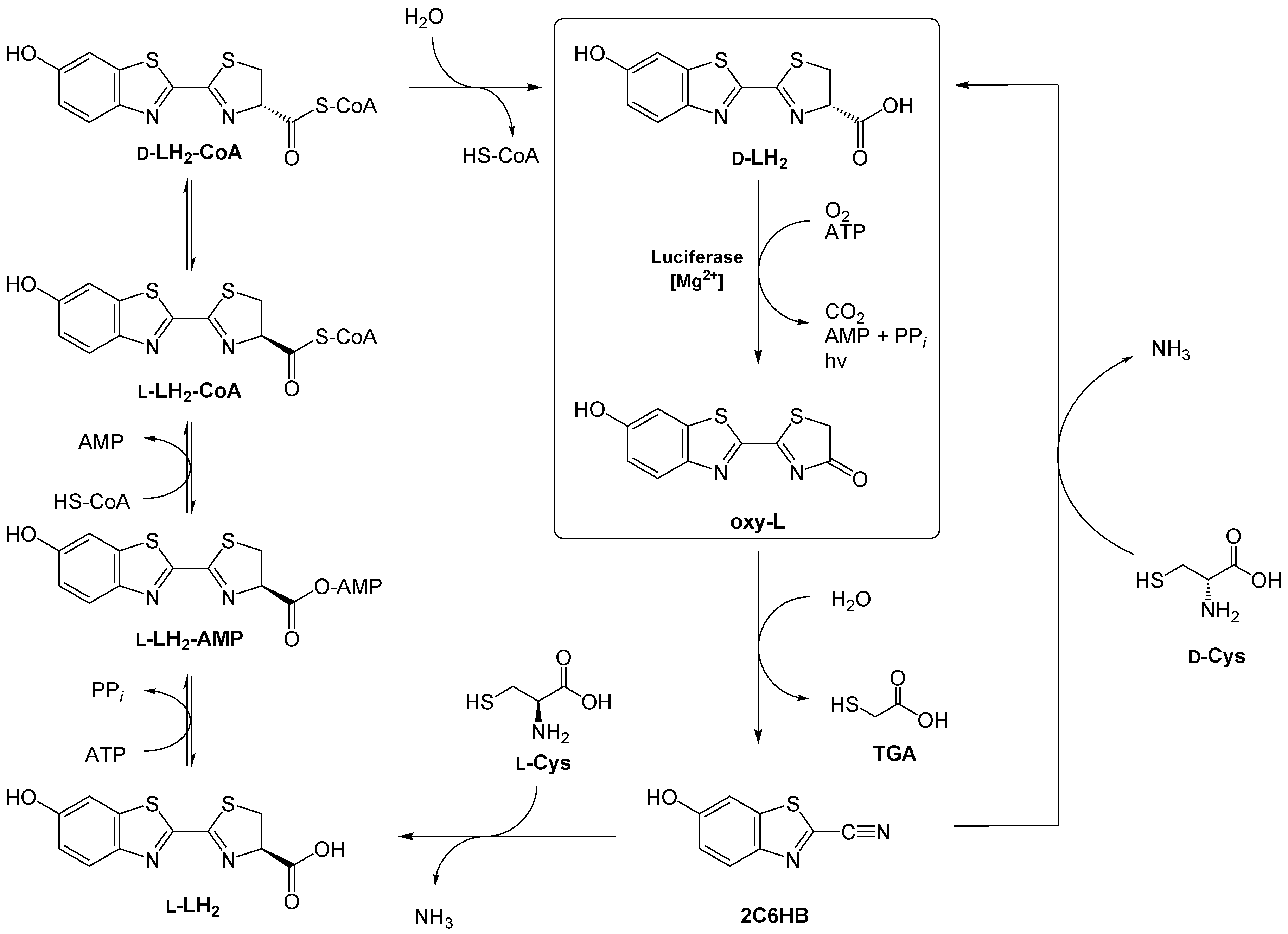
Wie die Insekten - oder mikrobielle Symbionten - des Luciferin herstellen, ist nicht ganz geklärt. Man weiß, dass D-Luciferin nicht direkt vom Käfer aufgenommen wird (es sei denn bei weiblichen Käfern der Art Photuris, die ihre männliche Artgenossen fressen).[9] Eine Möglichkeit besteht darin, das nach der Lichtreaktion entstandene Oxyluciferin zurück zu Luciferin zu recyclen. Hierbei soll zunächst das Oxyluciferin zu 2-Cyano-6-hydroxybenzothiazol (2C6HB) umgewandelt werden, was durch das Luciferin-regenerierende Enzym (LRE)[10] katalysiert wird. 2C6HB kondensiert dann mit einem D-Cystein zu D-Luciferin. Diese Kondensationsreaktion wird auch bei der chemischen Luciferinsynthese genutzt (vgl. Abbildung rechter Weg).
Eine seit kurzem diskutiere Möglichkeit geht indes davon aus, dass 2C6HB mit L-Cystein zunächst L-Luciferin bildet. Dieses wird dann über Zwischenschritte zu D-Luciferin racemisiert (vergleiche auch Abbildung linker Weg).[11]
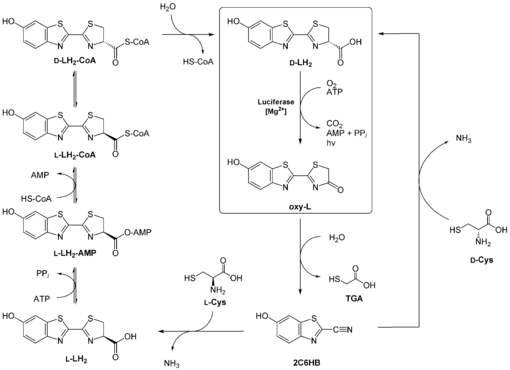 Zwei vorgeschlagene Mechanismen für die Regeneration D-Luciferins aus Oxyluciferin (siehe hierbei auch Text). Abkürzungen: LH2 Luciferin; HS-CoA Coenzym A; 2C6HB 2-Cyano-6-hydroxybenzothiazol; L Oxyluciferin; Cys Cystein; TGA Mercaptoessigsäure.
Zwei vorgeschlagene Mechanismen für die Regeneration D-Luciferins aus Oxyluciferin (siehe hierbei auch Text). Abkürzungen: LH2 Luciferin; HS-CoA Coenzym A; 2C6HB 2-Cyano-6-hydroxybenzothiazol; L Oxyluciferin; Cys Cystein; TGA Mercaptoessigsäure.Beide Möglichkeiten dieser Biosynthese weisen noch einige Probleme auf:
So wird das Luciferin-regenerierende Enzym in den lichtproduzierenden Organen biolumineszenter Käfer nicht überproduziert. Da Oxyluciferin in wässrigen Lösungen instabil ist, müsste man gerade dort erwarten, LRE in größeren Mengen vorzufinden. Außerdem kann das reaktive 2C6HB nicht nur mit Cystein, sondern auch mit anderen Metaboliten reagieren. Es ist ferner nicht klar, woher beispielsweise das D-Cystein stammt und wie zwischen L-Cystein und D-Cystein diskriminiert werden könnte. So kann zwar die Luciferase L-Luciferin als Substrat verwenden, dieses hemmt aber die Lichtreaktion.[12] Außerdem konnte man noch nicht das Enzym verifizieren, das die Racemisierung katalysiert. Es ist nicht einmal sicher, wie die Käfer (oder Symbionten) Benzothiazolene herstellen können.
Tetrapyrrol, das Luciferin von Dinoflagellaten
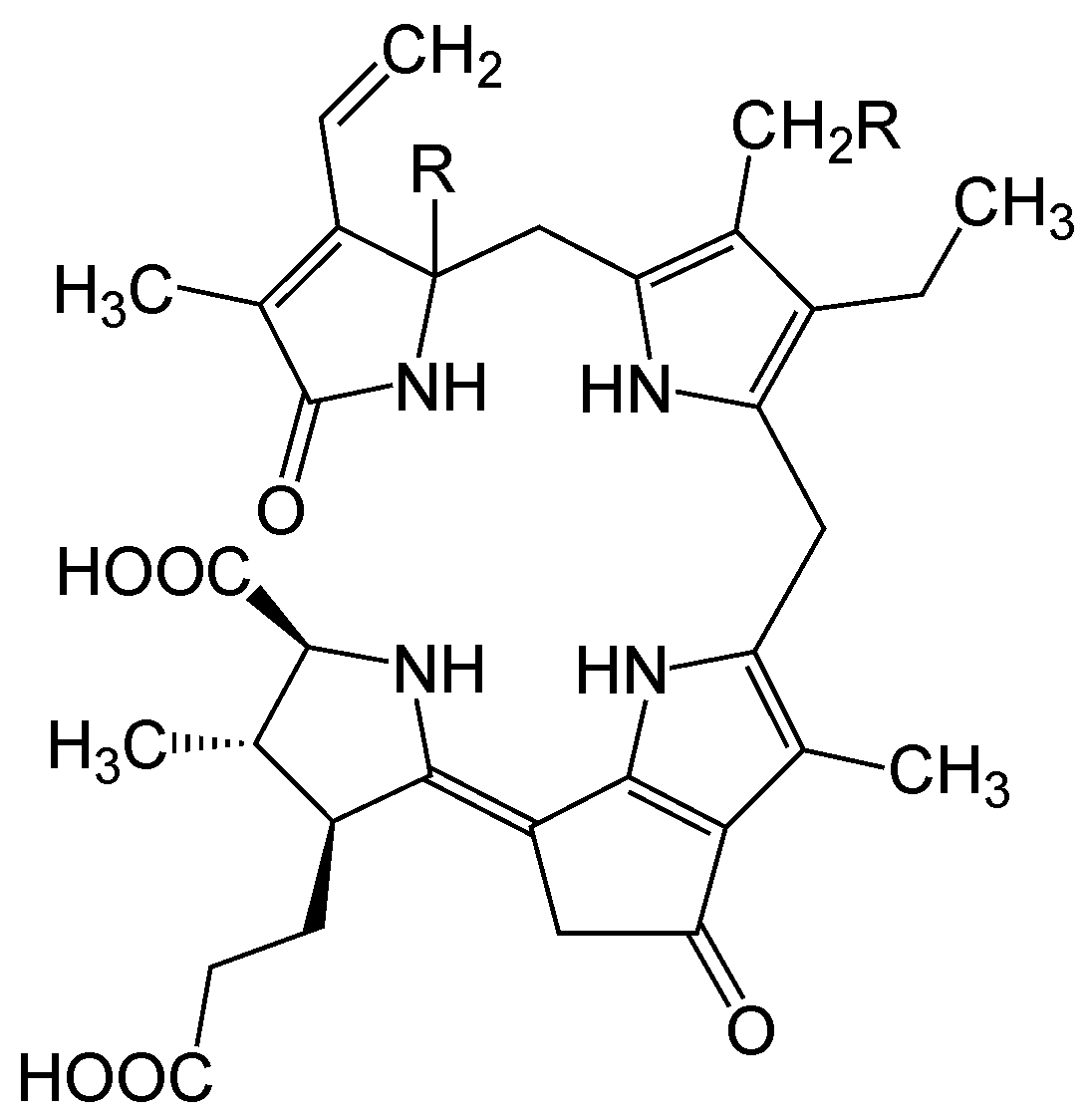
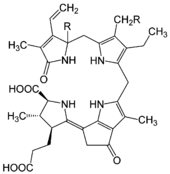 Das Luciferin aus Dinoflagellaten.
Das Luciferin aus Dinoflagellaten.Das Luciferin in dieser Gruppe entspricht dem chemischen Grundbau eines linearen Tetrapyrrols und findet sich bei Dinoflagellaten und bei Euphausiidae. Meeresleuchten, welches früher fälschlicherweise als Phosphoreszenz bezeichnet wurde, geht größtenteils auf diese einzelligen Algen zurück. Für die Lichtemission sind neben dem Luciferin und einer korrespondieren, ca. 137 kDa großen Luciferase (LCF) auch ein sogenanntes Luciferin-Bindeprotein (luciferin-binding protein (LBP)) notwendig.[13][14] Das Luciferin dieser Familie ist extrem instabil bei niedrigen pH-Werten, hohen Salz- und Sauerstoffkonzentrationen. Es konnte gezeigt werden, dass das LBD bei pH 8,0 an das Tetrapyrrol-Luciferin bindet, nicht aber bei pH 6,3.[15] Dadurch soll das Substrat bis zur Reaktion geschützt werden, die am besten bei pH 6,3 abläuft, zumal die Luciferase im leicht alkalischem Milieu (pH 8,0) inaktiv ist [16] Die Reaktion selbst findet dabei in speziellen Organellen statt, den sogenannten scintillons.[17] Das Licht, das bei dieser Reaktion entsteht, erscheint blau-grünlich (Emissionsmaximum von bei ca. λmax = 470 nm).
Es ist heute noch nicht geklärt, ob sich das Luciferin wegen seiner Verwandtschaft zu Chlorophyll a aus diesem ableitet oder erst aus mehreren Aminosäuren (Glycinen und Glutaminsäuren) schrittweise aufgebaut werden muss.[18] Außerdem ist es paradox, dass das bei der Lichtreaktion entsehende oxy-Luciferin kein Fluorophor ist.[19]
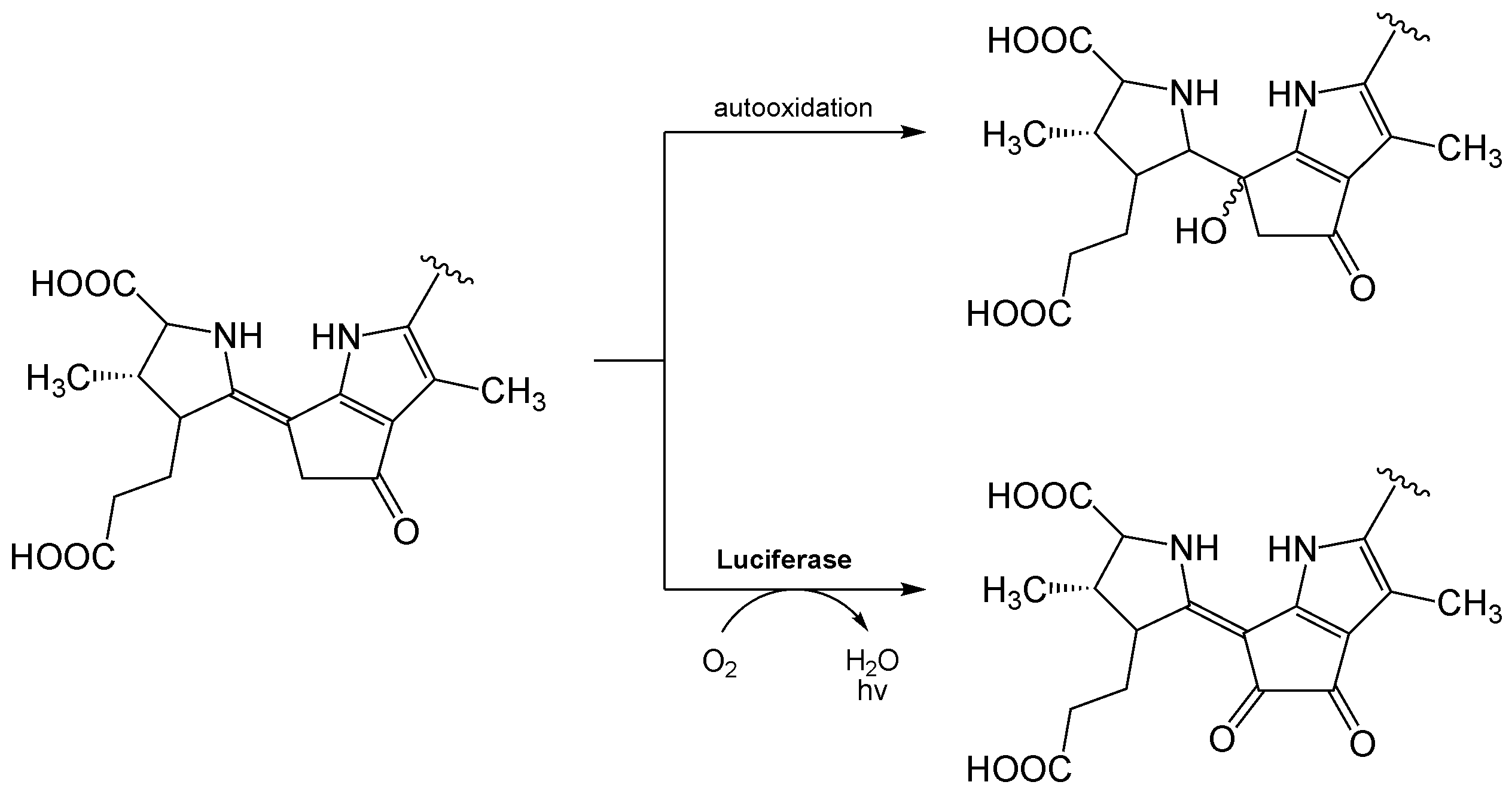
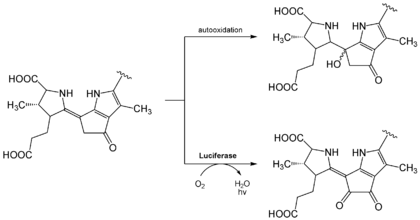 Biolumineszenzreaktion von Dinoflagellat-Luciferin. Bei der sog. „Lichtreaktion“ (unten) wird nach Oxidation Licht freigesetzt (λmax ≈ 470 nm). Durch Autooxidation kann jedoch auch ohne Luciferase eine sogenannte „Dunkelreaktion“ (oben) ablaufen, bei der kein Lichtquant emittiert wird.[19]
Biolumineszenzreaktion von Dinoflagellat-Luciferin. Bei der sog. „Lichtreaktion“ (unten) wird nach Oxidation Licht freigesetzt (λmax ≈ 470 nm). Durch Autooxidation kann jedoch auch ohne Luciferase eine sogenannte „Dunkelreaktion“ (oben) ablaufen, bei der kein Lichtquant emittiert wird.[19]Flavin, ein bakterielles Luciferin
 Bakterielles Luciferin, ein reduziertes Flavinmononukleotid (Riboflavin-5-phosphat).
Bakterielles Luciferin, ein reduziertes Flavinmononukleotid (Riboflavin-5-phosphat).Bakterielle Luciferasen nutzen reduzierte Flavinmononukleotide (FMNH2, was auch als Riboflavin-5-phosphat bezeichnet wird). FMNH2 ähnelt einem anderen Flavin, dem FADH2. Für die Lichtreaktion wird neben FMNH2 und molekularem Sauerstoff auch ein langkettiges Aldehyd benötigt. Es wird angenommen, dass hierfür Tetradekanal umgesetzt wird. Bei der durch die Flavin-abhängige Monooxygenase katalysierten Reaktion wird dabei blau-grünes Licht emittiert, dass ein Emissionsmaximum bei ca. λmax = 490 nm hat.
Alle Proteine, die etwas mit der Biolumineszenzreaktion zu tun haben, werden durch sogenannte lux-Gene kodiert (lat. lux Licht).
Das FMNH2 wird durch eine Riboflavinkinase unter ATP-Verbrauch aus Riboflavin (Vitamin B2) gewonnen. Nach Reaktion wird FMNH2 aus FMN regeneriert, was eine Flavinreduktase[20] unter NADPH-Verbrauch kataylisert. Auch das langkettige Aldehyd wird aus der bei der Reaktion entstandenen Fettsäure zurückgewonnen, was im sogenannten Fettsäure-Reduktase-Komplex[21] unter Verbrauch von ATP und NADPH katalysiert wird. Die Biolumineszenzreaktion verbraucht viel Energie, da für die Regenerierung der Komponenten bereits zwei Moleküle NADPH und ein Molekül ATP benötigt werden. Infolgedessen muss diese Lichtreaktion gut kontrolliert werden. Ein bekanntes Bakterium mit Biolumineszenzeigenschaft ist Vibrio fischeri.
 Umsetzung von bakteriellem Luziferin unter Verbrauch von Tetradecanal, einem langkettigen Aldehyd.
Umsetzung von bakteriellem Luziferin unter Verbrauch von Tetradecanal, einem langkettigen Aldehyd.Imidazolpryazin, die gemeinsame chemische Komponente des Coelentrazins und des Vargulins
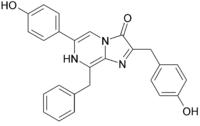
1962 wurde das Photoprotein Aequorin aus Aequorea victoria isoliert und dabei 1974 Coelenterazin als das Luciferin identifiziert.[23][24] Dieses Imidozolpyrazin ist in vielen anderen biolumineszenten marinen Organismen vertreten, z. B. in Tintenfischen, Garnelen (Mysida), Fischen, Ruderfußkrebsen (Copepoda) sowie Zehnfußkrebsen (Decapoda) und anderen Hohltieren (Cnidaria; Ctenophora), aber auch in nicht biolumineszenten Meeresbewohnern.[25][26][27] Auch Derivate von Coelenterazin werden von zahlreichen marinen Lebewesen genutzt. So ist beispielsweise das Luciferin von Watasenia scintillans[28] ein Disulfid.
Wie nun die biochemischen Mechanismen für das Luciferin-Luciferase-System mit Imidazolpryazinen ablaufen, wurde 2000 anhand von A. victoria gezeigt.[29] Hierbei spielt das Aequorin eine wesentliche Rolle, bei dem bereits das Luciferin (Coelenterazin) durch eine Peroxidbrücke mit dem Proteinteil verbunden ist. Infolgedessen führt das Photoprotein bereits das oxidierende Agens mit sich. Aequorin besitzt drei Bindestellen für Calciumionen. Wenn Calciumionen daran binden, ändert sich die Konformation des Proteins derart, dass eine intramolekulare Reaktion mit dem Coelenterazin ausgelöst wird. Dieses reagiert zunächst zu einem instabilen Dioxetan, so dass nach Abspaltung von CO2 schließlich das Anion von Coeleteramid entsteht. Nach Relaxation in den Grundzustand wird ein Lichtquant mit einer Wellenlänge von λmax = 465 nm emittiert. Wegen dieses blauen Leuchtens wird das Protein auch als das blaufluoreszierende Protein (blue fluorescent protein) (BFP)[30] bezeichnet. Das Photoprotein wird in Anwesenheit von Coelenterazin und molekularem Sauerstoff schließlich regeneriert.
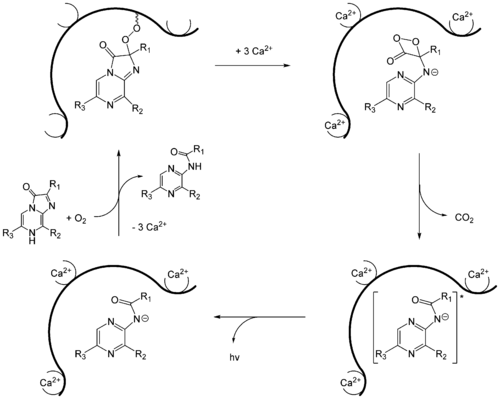 Mechanismus der Biolumineszenz von Aequorin.
Mechanismus der Biolumineszenz von Aequorin.Aequorea victoria fluoresziert aber nicht blau, sondern grün. Das liegt daran, dass das blau fluoreszierende Protein die Energie der Biolumineszenzreaktion strahlungslos auf das sogenannte grün fluoreszierende Protein (GFP) überträgt.
Manche marine Meeresorganismen nutzen Luciferine, die dem Coelenterazin ähneln. Jedoch ist bei diesen gemeinsam, dass sie im Zuge der Lichtreaktion das Anion des Coeleteramids erzeugen. Bei W. scintillans katalysiert eine membrangebundene Luciferase unter Verbrauch von ATP, Magesiumionen sowie Sauerstoff die Lichreaktion, dabei werden ATP, Magesiumionen sowie Sauerstoff für die Umsetzung von Coelenterazindisulfat benötigt. Es entsteht dabei Licht zwischen 400 bis 580 nm (λmax = 470 nm).[31]
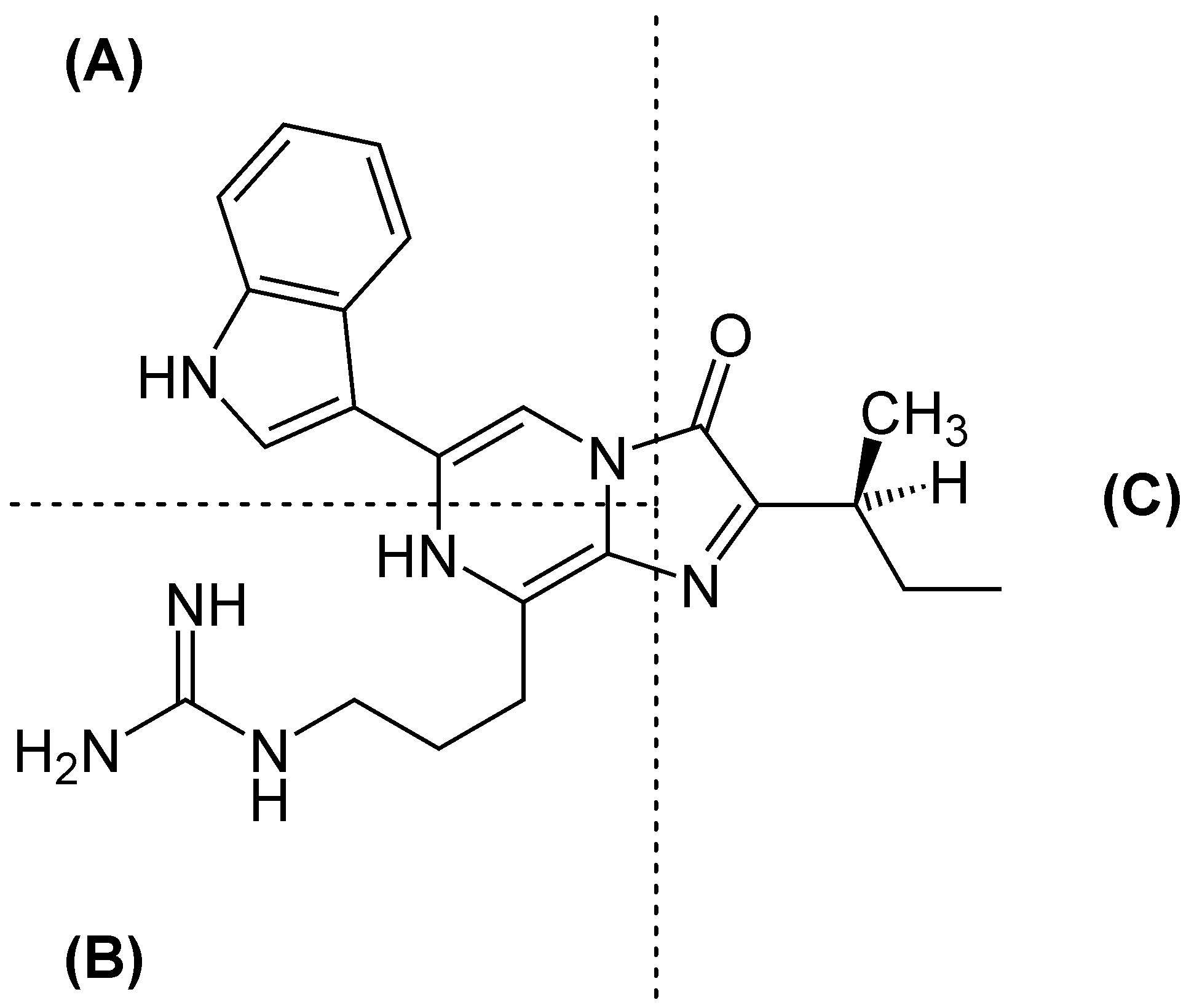
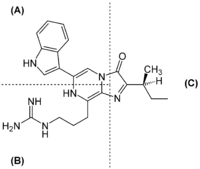 Das Luciferin aus Vargula hilgendorfii, auch als Vargula- bzw. Cypridina-Luciferin bekannt, besteht aus einer Tryptophan- (A), einer Arginin- (B) sowie einer Isoleucineinheit (C).
Das Luciferin aus Vargula hilgendorfii, auch als Vargula- bzw. Cypridina-Luciferin bekannt, besteht aus einer Tryptophan- (A), einer Arginin- (B) sowie einer Isoleucineinheit (C).Vargulin aus Muschelkrebsen (Vargula hilgendorfii oder auch Cypridina hilgendorfii) wurde 1957 isoliert und 1966 als Imidazolpryazinkomponente identifiziert. Die Quantenausbeute liegt bei Q = 0,3. [32] Auch hier entsteht nach Ablauf der Lichtreaktion das Coeleteramid der Komponente. Bereits 1966 wurde vermutete, dass jenes Luciferin aus L-Arginin, L-Isoleucin und L-Tryptophan aufgebaut wird, wofür es mittlerweile immer mehr Hinweise gibt.[33][34]
 Allgemeine Umsetzung eines Celentreazins durch eine korrespondierende Luciferase zu einem Coelenteramid.
Allgemeine Umsetzung eines Celentreazins durch eine korrespondierende Luciferase zu einem Coelenteramid.Nicht klassische Luciferin-Luciferase-Systeme
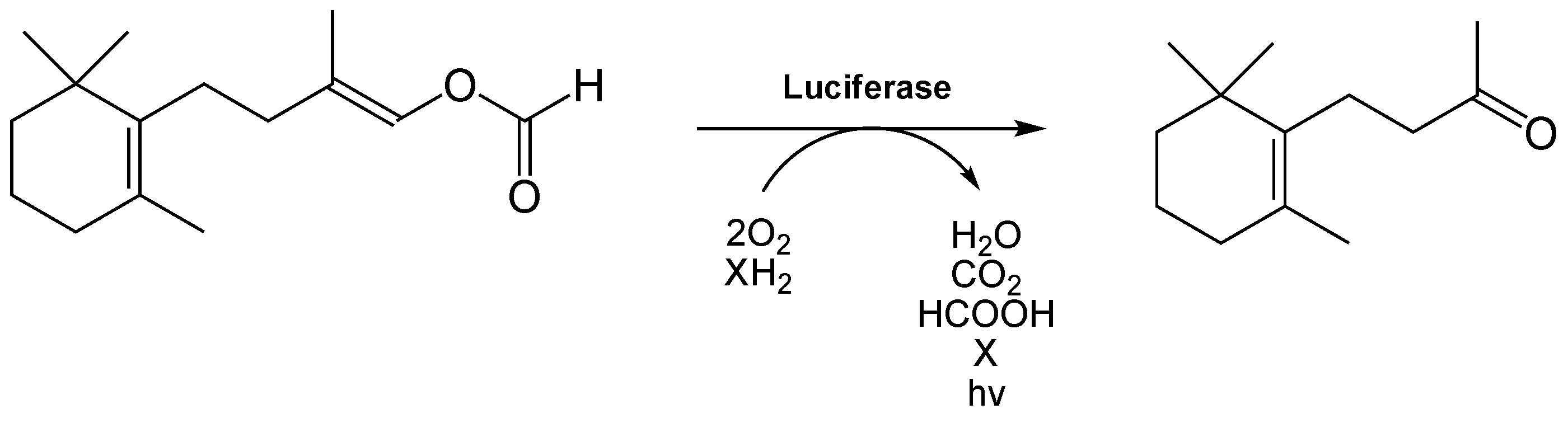
Das bei der Süßwasserschnecke Neuseelands (Latia neritoides) vorkommende Luciferin[35] ist ein terpenoides Aldehyd und wird als Latia-Luciferin bezeichnet.[36][37]
Es wird katalytisch zu einem Keton (oxy-Luciferin) ungesetzt, was durch eine 173 kDa große Luciferase (EC-Nummer 1.14.99.21), ein Homohexamer, katalysiert wird.[38] Für die Reaktion ist als Cofaktor das 39 kDa große purple protein[36][37] aber nicht unbedingt nötig.[38] Bei der Reaktion werden neben CO2, H2O und Ameisensäure auch Licht freigesetzt, dessen Emissionsmaximum bei λmax = 536 nm liegt.[39] Daher erscheint der Schleim der Schnecke, der z. B. nach mechanischen Reizen abgesondert wird, grün zu lumineszieren.
Das entstehende oxy-Luciferin ist aber im Gegensatz zu oxy-Luciferin des Leuchtkäfers kein Fluorophor. Es wird vermutet, dass die bei dieser Reaktion freiwerdende Energie auf den eigentlichen Emitter übertragen wird, einem proteingebundenen Flavin.[40][38] Die Reaktionseffizienz dieser Biolumineszenzreaktion ist sehr gering, da die Quantenausbeute Q bei ca. 0,003 liegt.[38]
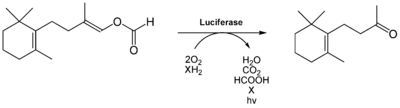 Luciferinreaktion des Luciferins von Latia neritoides. Die reduzierende Komponente X sowie der eigentliche Lichtemitter der Reaktion müssen noch identifiziert werden.
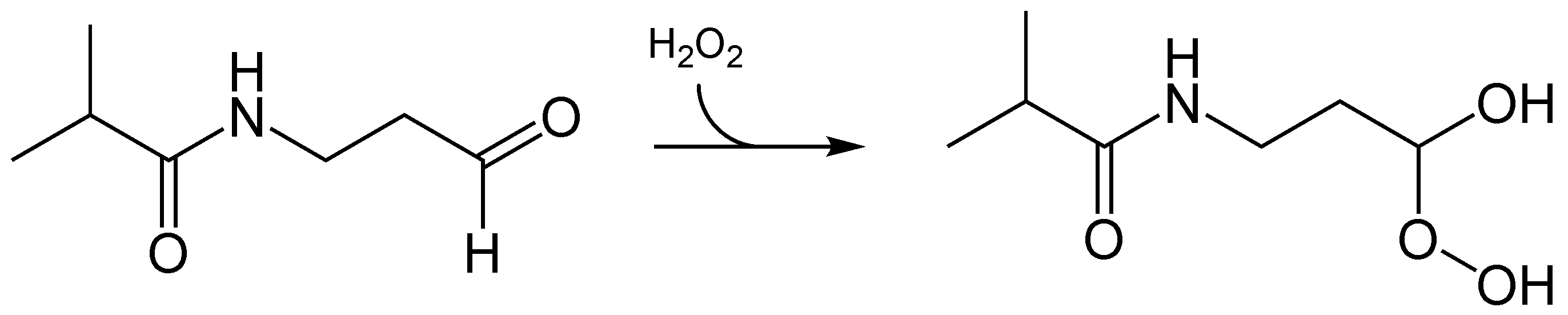
Luciferinreaktion des Luciferins von Latia neritoides. Die reduzierende Komponente X sowie der eigentliche Lichtemitter der Reaktion müssen noch identifiziert werden.Das Luciferin des Wurmes Diplocardia longa ist ein einfaches Aldehyd, das N-Isovaleryl-3-aminopropanal. Das besondere bei der Biolumineszenzreaktion ist die Tatsache, dass Wasserstoffperoxid an Stelle von molekularen Sauerstoff benötigt wird. Die korrespondierende Luciferase, eine 300 kD großes Enzym, setzt dann die aktivierte Form, ein Peroxid-Addukt, um. Die Luciferase benötigt Kupfer, es wird blau-grünes Licht emittiert. Man weiß jedoch nicht, welchen Zweck die Biolumineszenz bei Würmern generell haben kann.[41][42] Auch hier muss der eigentliche Emittter der Biolumineszenzreaktion noch identifiziert werden.
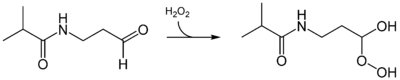 Im Gegensatz zu den meisten Luciferinen wird das Luciferin des Wurmes Diplocardia longa mittels Wasserstoffperoxid oxidiert und dann umgesetzt.
Im Gegensatz zu den meisten Luciferinen wird das Luciferin des Wurmes Diplocardia longa mittels Wasserstoffperoxid oxidiert und dann umgesetzt.Anwendungen
Diagnostik
Mit Hilfe des Luciferin-Luciferase-System aus Leuchtkäfern kann die Anwesenheit von ATP in Proben schnell überprüft werden.[43]
Da die Lichtreaktion Aequorins von Calciumionen abhängig ist, kann durch dieses System die Konzentration an Calciumionen gemessen werden. Die erste Anwendung datiert sich auf 1967, als man mit Hilfe von Aequorins intrazelluläre Änderungen der Calciumkonzentrationen in Muskelzellen detektiert hat. Nach Klonierung von Aequorins in Bakterien konnte man die Calciumkonzentraiton bateriellen Cytosols messen.[44] Außerdem ist es möglich, Aequorin in eukaryontische Zellen klonieren.[45] So konnte man beispielsweise bei transgenen Pflanzen die Änderung der cytosolischen Calciumkonzentration nach Berühren der Pflanze oder nach einem Kälteschock messen.[46]
Gentechnik
Luciferasen werden in der Molekularbiologie oft als Marker eingesetzt: Organismen, die das Gen erhalten und in ihr Genom eingebaut haben, leuchten bei Zufuhr von Luciferin. So könnte man beispielsweise nachweisen, ob Gene, die man in Organismen einbringen möchte, auch wirklich exprimiert werden. Dafür wird einfach das zu exprmierende Gen mit dem einem Gen gekoppelt, das für eine Luciferase kodiert.
Einzelnachweise
- ↑ Harvey, EN. Bioluminescence. Academic, New York 1952.
- ↑ Hastings, JW. Bioluminescence. (N. Sperelakis, ed.)., in: Cell Physiology, Dritte Edition, Academic Press, NY., 1115–1131.
- ↑ Herring PJ. (1987): Systematic distribution of bioluminescence in living organisms, in: J Biolumin Chemilumin. 1 (3); 147–163; PMID 3503524.
- ↑ White, EH. et al. (1971): The chemi- and bioluminescence of firefly luciferin: an efficient chemical production of electronically excited states, in: Bioorg. Chem. 1 (1–2); 92–122; doi:10.1016/0045-2068(71)90009-5.
- ↑ PubChem 5459882 (Firefly-Luciferin).
- ↑ NiceZyme View of ENZYME: EC 1.13.12.7.
- ↑ Keller, GA. et al. (1987): Firefly luciferase is targeted to peroxisomes in mammalian cells, in: Proc Natl Acad Sci USA. 84 (10): 3264–3268; PMID 3554235; PDF (freier Volltextzugriff, engl.)
- ↑ Ando, Y. et al. (2008): Firefly bioluminescence quantum yield and colour change by pH-sensitive green emission, in: Nat. Photonics 2; 44–47; doi:10.1038/nphoton.2007.251.
- ↑ Eisner T. et al. (1997): Firefly »femmes fatales« acquire defensive steroids (lucibufagins) from their firefly prey, in: Proc Natl Acad Sci USA 94 (18), 9723–9728; PMID 9275191; PDF (freier Volltextzugriff, engl.)
- ↑ Gomi K. und Kajiyama N. (2001): Oxyluciferin, a luminescence product of firefly luciferase, is enzymatically regenerated into luciferin, in: J Biol Chem. 276 (39); 36508–36513; PMID 11457857; PDF (freier Volltextzugriff, engl.)
- ↑ Niwa, K. et al. (2006): Stereoisomeric bio-inversion key to biosynthesis of firefly D-luciferin, in: FEBS Lett. 580 (22); 5283−5287; PMID 16979628; PDF (freier Volltextzugriff, engl.)
- ↑ Lembert N. (1996): Firefly luciferase can use L-luciferin to produce light, in: Biochem J. 317 (Pt 1); 273–277; PMID 8694774;PDF (freier Volltextzugriff, engl.)
- ↑ Morse, D. et al. (1989): Role of a luciferin-binding protein in the circadian bioluminescent reaction of Gonyaulax polyedra, in: J Biol Chem. 264 (20), 11822–11826; PMID 2745419; PDF (freier Volltextzugriff, engl.)
- ↑ Morse, D. und Mittag, M. (2000): Dinoflagellate luciferin-binding protein, in: Methods Enzymol. 305, 258–276; PMID 10812606.
- ↑ Fogel, M. und Hastings, JW. (1971): A substrate-binding protein in the Gonyaulax bioluminescence reaction, in: Arch Biochem Biophys. 142(1): 310–321; PMID 5545485.
- ↑ Schultz, LW. et al. (2005): Crystal structure of a pH-regulated luciferase catalyzing the bioluminescent oxidation of an open tetrapyrrole, in: Proc Natl Acad Sci USA 102(5): 1378–1383; PMID 15665092; PDF (freier Volltextzugriff, engl.)
- ↑ Fogel M und Hastings JW. (1972): Bioluminescence: mechanism and mode of control of scintillon activity, in: Proc Natl Acad Sci USA 69(3):690–369; PMID 4501583; PDF (freier Volltextzugriff, engl.)
- ↑ Wu, C. et al. (2003): Tracer studies on dinoflagellate luciferin with [15N]-glycine and [15N]-l-glutamic acid in the dinoflagellate Pyrocystis lunula, in: Tetrahedron Letters 44(6); 1263–1266; doi:doi:10.1016/S0040-4039(02)02815-0.
- ↑ a b T. Wilson and JW. Hastings (1998), S. 208.
- ↑ Duane, W. und Hastings, JW. (1975): Flavin mononucleotide reductase of luminous bacteria, in: Mol Cell Biochem. 6(1); 53–64; PMID 47604.
- ↑ Rodriguez A. et al. (1983): Purification of the acyl coenzyme A reductase component from a complex responsible for the reduction of fatty acids in bioluminescent bacteria. Properties and acyltransferase activity, in: J Biol Chem. 258(8); 5233–5237; PMID 6833298; PDF (freier Volltextzugriff, engl.)
- ↑ PubChem 2830 (Coelenterazin-Luciferin).
- ↑ Shimomura, O. et al. (1963): Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea, in: J Cell Comp Physiol. 59; 223–239; PMID 13911999.
- ↑ Shimomura, O. et al. (1974): Mechanism of the luminescent intramolecular reaction of aequorin, in: Biochemistry 13(16); 3278–3286; PMID 4152180.
- ↑ Shimomura, O. et al. (1980): Widespread occurrence of coelenterazine in marine bioluminescence, in: Comp. Biochem. Physiol. 65B, 435–437.
- ↑ Campbell, AK. und Herring, PJ. (1990): Imidazolopyrazine bioluminescence in copepods and other marine organisms, in: Mar. Biol. 104(2); 219–225; doi:10.1007/BF01313261.
- ↑ Shimomura, O. (1987): Presence of coelenterazine in non-bioluminescent marine organisms, in: Comp Biochem Physiol 86B (1987); 361–363.
- ↑ S. Watasé (1905): The luminous organ of firefly squid, in: Dobutsugaku Zasshi 17, 119–123 (Jap.)
- ↑ Head JF. et al. (2000): The crystal structure of the photoprotein aequorin at 2.3 Å resolution, in: Nature. 405(6784); 291–293; PMID 10830969; PDF (freier Volltextzugriff, engl.)
- ↑ Shimomura, O. und Johnson, FH. (1970): Calcium binding, quantum yield, and emitting molecule in aequorin bioluminescence, in: Nature 227(5265); 1356–1357; PMID 4393938.
- ↑ Tsuji FI. (2005): Role of molecular oxygen in the bioluminescence of the firefly squid, Watasenia scintillans, in: Biochem Biophys Res Commun. 338(1): 250–253; PMID 16165097.
- ↑ Johnson, FH. und Shimomura, O. (1972): Enzymatic and nonenzymatic bioluminescence, in: Photophysiology (7); 275–334; PMID 4376836.
- ↑ Kato, S. et al. (2004): Identification of the biosynthetic units of Cypridina luciferin in Cypridina (Vargula) hilgendorfii by LC/ESI-TOF-MS, in: Tetrahedron 60(50); 11427–11434; doi:10.1016/j.tet.2004.09.080.
- ↑ Kato S. et al. (2006): Stereoselective incorporation of isoleucine into Cypridina luciferin in Cypridina hilgendorfii (Vargula hilgendorfii), in: Biosci Biotechnol Biochem. 70(6); 1528–1532; PMID 16794342; PDF (freier Volltextzugriff, engl.)
- ↑ PubChem 5280505 (Latia-Luciferin).
- ↑ a b Shimomura, O. und Johnson, FH. (1968): The structure of Latia luciferin, in: Biochemistry 7(5): 1734–1738; PMID 5650377.
- ↑ a b Shimomura, O. et al. (1972): Reactions Involved in Bioluminescence Systems of Limpet (Latia neritoides) and Luminous Bacteria, in: Proc Natl Acad Sci USA 69 (8), 2086–2089; PMID 4506078; PDF (freier Volltextzugriff, engl.)
- ↑ a b c d Ohmiya, Y. et al. (2005): Bioluminescence in the Limpet-Like Snail, Latia neritoides, in: Bull. Chem. Soc. Jpn. 78 (7); 1197–1205; doi:10.1246/bcsj.78.1197.
- ↑ Nakamura, M. et al. (2004): Synthesis of Latia luciferin benzoate analogues and their bioluminescent activity, in: Tetrahedron Letters 45 (10), 2203–2205, doi:10.1016/j.tetlet.2004.01.027.
- ↑ Vadim R. Viviani: The Biological and Biochemical Diversity of Terrestrial Bioluminescence.
- ↑ Rudie, NG. et al. (1976): Purification and properties of luciferin from the bioluminescent earthworm, Diplocardia longa, in: Photochem Photobiol. 23 (1): 71–75; PMID 1265130.
- ↑ Rudie, NG. et al. (1981): Earthworm bioluminescence: characterization of high specific activity Diplocardia longa luciferase and the reaction it catalyzes, in: Biochemistry 20 (2): 344–350; PMID 6258637.
- ↑ Neufeld HA. et al. (1975): A rapid method for determining ATP by the firefly luciferin-luciferase system, in: Experientia 31 (3), 391–392; PMID 1116561.
- ↑ Knight, MR. et al. (1991): Recombinant aequorin as a probe for cytosolic free Ca2+ in Escherichia coli, in: FEBS Lett. 282 (2); 405–408; PMID 2037058; PDF (freier Volltextzugriff, engl.)
- ↑ Kendall, JM et al. (1992): Targeting aequorin to the endoplasmic reticulum of living cells., in: Biochem. Biophys. Res. Commun. 189 (2); 1008–1016; PMID 1472014.
- ↑ Knight, MR. et al. (1991): Transgenic plant aequorin reports the effects of touch and cold-shock and elicitors on cytoplasmic calcium, in: Nature 352(6335); 524–526; PMID 1865907.
Literatur
- Osamu Shimomura: Bioluminescence: Chemical Principles and Methods. Word Scientific Publishing Company 2006; ISBN 9-81256801-8
- T. Wilson and JW. Hastings (1998): Bioluminscence, in: Annu. Rev. Cell Dev. 14; 197–230; PMID 9891783
- Greer III., LF. und Szalay, AA. (2002): Imaging of light emission from the expression of luciferases in living cells and organisms: a review, in: Luminescence 17, 43–74; PMID 11816060 doi:10.1002/bio.676
- K. Teranishi (2007): Luminescence of imidazo[1,2-a]pyrazin-3(7H)-one compounds, in: Bioorg Chem. 35(1); 82–111; PMID 17007903
Weblinks
- Luciferin-Luciferase Systeme (engl.)
- Lumineszenz - Chemilumineszenz - Biolumineszenz von Dieter Weiß (Uni Jena)
- Übersichtsseite (American Society for Photobiology, engl.)
- Firefly Bioluminescence von Bruce R. Branchini
- Dinoflagellate Bioluminescence von J. Woodland Hastings, Wayne Schultz und Liyun Liu
- Bacterial Bioluminescence von Leo Yen-Cheng Lin und Edward A. Meighen
- Terrestrial Bioluminescence von Vadim R. Viviani


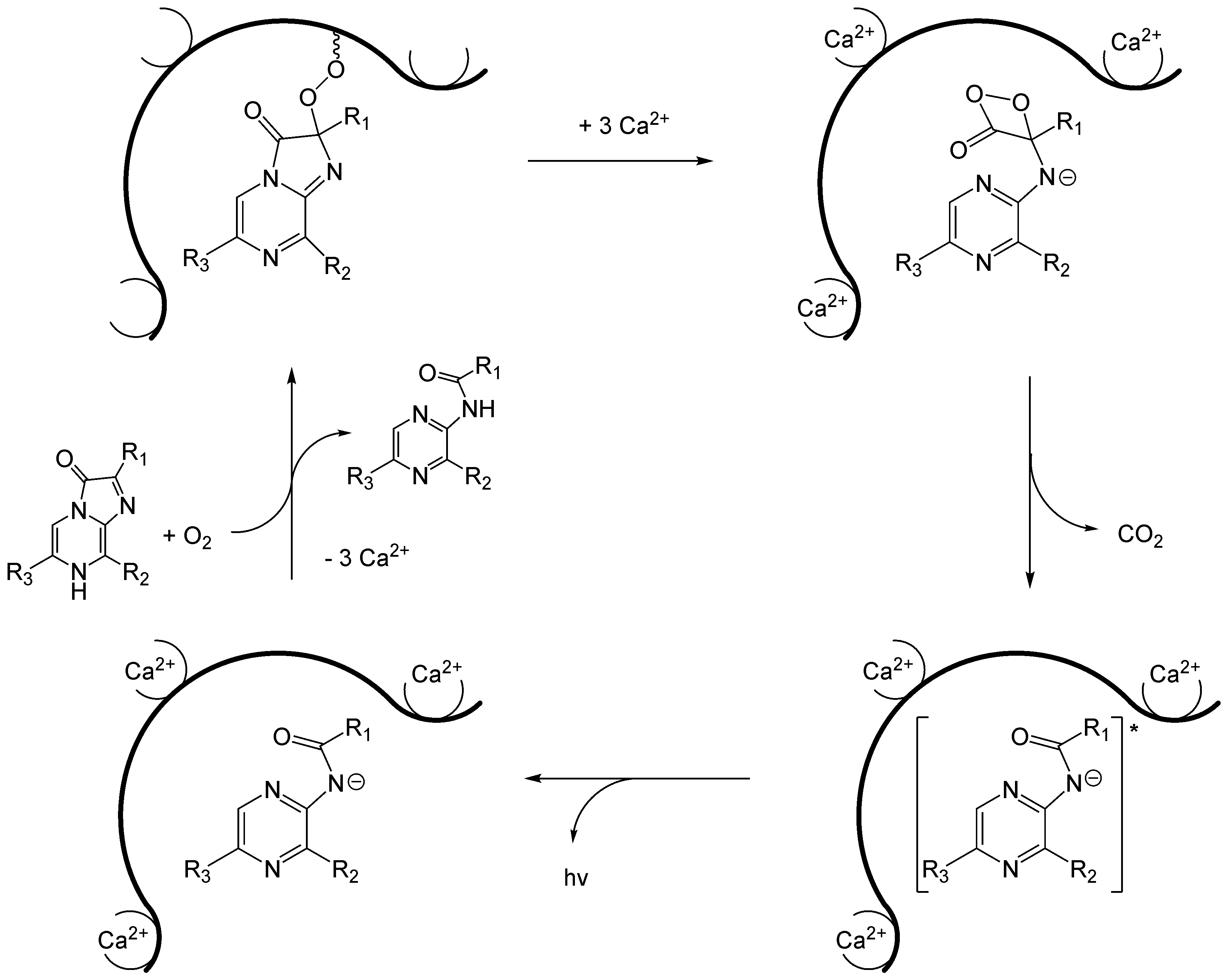

Wikimedia Foundation.