- Rüsseltiere
-
Rüsseltiere 
Zeitraum Seelandium (Paläozän) bis heute 60 bis 0 Mio. Jahre Fundorte - Afrika, Asien, Europa, Nord- und Südamerika
Systematik Überklasse: Kiefermäuler (Gnathostomata) Reihe: Landwirbeltiere (Tetrapoda) Klasse: Säugetiere (Mammalia) Unterklasse: Höhere Säugetiere (Eutheria) Überordnung: Afrotheria Ordnung: Rüsseltiere Wissenschaftlicher Name Proboscidea Illiger 1811 Die Ordnung der Rüsseltiere (Proboscidea) wurde nach dem auffälligsten Merkmal, dem Rüssel (lat. proboscis), benannt. Ihre einzigen heute noch lebenden Vertreter sind die Elefanten.
Inhaltsverzeichnis
Merkmale
Allgemein
Die Rüsseltiere unterscheiden sich nicht nur durch den markanten Rüssel, sondern auch durch ihren Körperbau und ihre charakteristische Bezahnung (Stoßzähne bzw. große Mahlzähne) von allen anderen Landsäugern. Besonders markant ist die Rüsselbildung, welche anfänglich kaum vorhanden war und den frühen, wohl sumpflebenden Rüsseltieren wahrscheinlich als Schnorchel diente. Später entwickelte sich der Rüssel, der bei heutigen Tieren aus bis zu 150.000 längs, zirkular oder schräg verlaufenden Muskelfasern einer Vielzahl verschiedener Muskeln besteht, zu einem feinfühligen Greiforgan, welches das Erreichen der Blätter auf höheren Bäumen ebenso ermöglichte wie das Abreißen von Grasbüscheln in den Steppen.[1]
Schädel und Skelett
Besondere skelettanatomische Merkmale sind die säulenförmigen Gliedmaßen, die senkrecht unter dem Körper stehen, wobei die oberen und unteren Partien der Extremitäten einen Winkel von 180° bilden. Dies unterscheidet die Rüsseltiere von zahlreichen anderen Säugetieren, deren Beine in einem leichten Winkel angeordnet sind. Die deutlich vertikale Stellung, die möglicherweise schon im Eozän vollständig ausgebildet war, unterstützte dabei die enorme Gewichtszunahme der frühen Vertreter dieser Ordnung.[2] Weiterhin besitzen die Langknochen keine Knochenmarkhöhle sondern der Raum ist mit Spongiosa gefüllt, was den Beinen eine größere Festigkeit gibt. Die Blutbildung findet dabei in den Zwischenräumen statt.[1]
Eine andere Besonderheit weist der sehr große Kopf der Rüsseltiere auf, dessen Schädeldach aus luftgefüllten Hohlräumen besteht. Diese bienenwabenartig geformten Hohlräume, die durch dünne Knochenplättchen voneinander getrennt sind, verringern nicht nur das Gewicht des gesamten Schädels, sondern ermöglichten gleichzeitig auch einen enormen Zuwachs an Volumen der Schädeloberfläche. Dieser Volumenzuwachs war notwendig, um einerseits über die mächtige Nackenmuskulatur den Halt des Kopfes inklusive der evolutiv immer größer werdenden Stoßzähne zu gewährleisten, andererseits aber auch um der kräftigen Kaumuskulatur für den massiven Unterkiefer als Ansatzfläche zu dienen. Die Entwicklung eines derartigen luftgefüllten Schädels begann stammesgeschichtlich schon sehr früh bei den Rüsseltieren und ist bei einigen Vertretern schon im Oligozän, möglicherweise auch schon im späten Eozän nachgewiesen.[1][3]
Gebiss
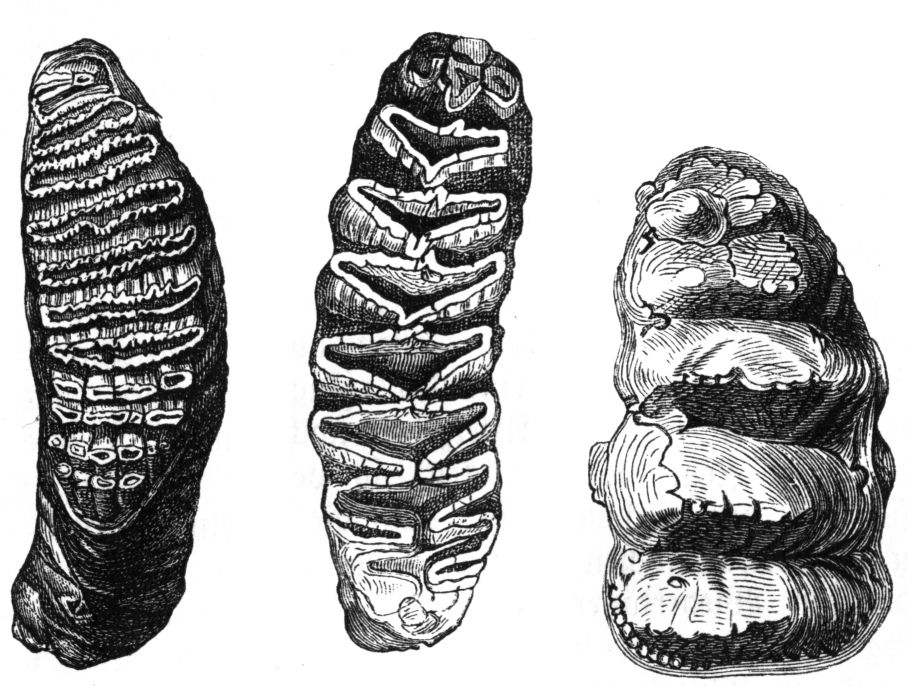
Das ursprüngliche permanente Rüsseltiergebiss besaß noch die vollständige Bezahnung der relativ altertümlichen tertiären Säugetiere mit drei Schneidezähnen, einem Eckzahn, vier Prämolaren und drei Molaren je Kieferast.[4] Im Laufe der stammesgeschichtlichen Evolution reduzierte sich die Zahnanzahl kontinuierlich bis zu den heutigen Elefanten, die nur einen Schneidezahn im Oberkiefer (Stoßzahn) und drei Molaren je Kieferbogen aufweisen, wovon aber nur jeweils ein Molar im Ober- und im Unterkiefer in jeder Kieferhälfte zur Verfügung steht. Diese werden beim Kauen der Pflanzennahrung stark abgenutzt, können jedoch bei den heutigen Elefanten fünf Mal nachgeschoben werden, umfassen also sechs Generationen. Diese sechs Generationen beinhalten drei Milchprämolaren (diese werden von einigen Experten aufgrund der starken Ähnlichkeit zu den Molaren auch als Milchmolaren angesprochen,[5] sind aber aus ontogenetischer Sicht Prämolaren) und drei Dauermolaren.[1] Der Zahnwechsel erfolgt dabei horizontal, indem sich ein neuer Zahn erst von hinten herausschiebt, wenn der vordere weitgehend abgekaut ist. Dieser horizontale Zahnwechsel, der sich von dem vertikalen der meisten andern Säugetiere deutlich unterscheidet, entstand durch die Verkürzung des Kiefers in der Rüsseltierevolution, so dass nicht mehr alle Zähne gleichzeitig Platz fanden. Erstmals aufgetreten ist dieses Merkmal im mittleren Oligozän.[6] Auch die Stoßzähne des Oberkiefers fallen bei den jungen Elefanten nach dem ersten Jahr aus, werden dann aber durch ständig wachsende ersetzt.
Zur systematischen Einteilung der Rüsseltiere wird hauptsächlich der Aufbau der Molaren herangezogen. Die Kaufläche dieser Mahlzähne ist sehr vielgestaltig und der jeweiligen Lebensweise der Tiere angepasst. Sie kann höckerige Strukturen (bunodont) aufweisen, wie bei einigen sehr frühen Rüsseltieren, oder aus einzelnen Querleisten aufgebaut sein (lophodont bis zygodont), die teilweise durch verschieden starke Abnutzung härterer und weicherer Stellen eine dachartige Struktur ausbilden wie bei den Stegodonten bzw. lamellenartig geformt sein wie bei den Mammuts und den heutigen Elefanten.[1]
Stoßzähne
Markantestes Skelettelement sind die Stoßzähne, die gerade, gedrillt, nach oben oder unten gebogen, schaufelförmig, dem Ober- oder Unterkiefer oder beiden entspringend, weit auseinander oder eng zusammenstehend, und in den verschiedensten Längen (bis zu 4 m) und Stärken vorhanden waren. Die Stoßzähne sind eine Bildung der Schneidezähne, die anfänglich noch recht klein und weitgehend senkrecht im Kiefer standen, später in Verbindung mit der allgemeinen Reduktion der Zahnanzahl im Gebiss der Rüsseltiere aber einen übergroßen Wuchs erreichten. Dabei entstanden die Stoßzähne des Oberkiefers aus den zweiten Schneidezähnen (I2) des jeweiligen Kieferastes, während der Ursprung der Unterkieferstoßzähne unter Experten lange diskutiert, mittlerweile aber der erste Schneidezahn (I1) des Kieferastes identifiziert wurde.[7] Heute leben nur noch die Elefanten mit ihren beiden Stoßzähnen im Oberkiefer.
Ökologie
Die Anpassungen der Stoß- und besonders der Backenzähne ermöglichten den Rüsseltieren die Erschließung verschiedener pflanzlicher Nahrungsquellen, von den Sumpfpflanzen, die dem frühen Moeritherium als Nahrung dienten, über Blätter und Zweige, die mit den Rüsseln abgerissen wurden, oder Rinden, die wie bei den Deinotherien mit den Stoßzähnen abgeschält werden konnten, bis zu den Gräsern der Savannen und Steppen, welche u. a. die Mammuts bevorzugten und die besonders intensiv gekaut werden müssen.
Ein zotteliges Haarkleid schützte einige nördliche Arten der Echten Mastodonten und Mammuts vor der Kälte der Eiszeit. Die beiden riesigen gebogenen Stoßzähne des Mammuts scheinen auch als Schneepflug zur Nahrungssuche unter der Schneedecke wertvolle Dienste geleistet zu haben. Zwergbildungen bei Stegodonten und Elefanten (siehe: Zwergelefant) ermöglichten über lange Zeiträume hinweg das Überleben auch auf kleineren Inseln des Mittelmeerraums und in Indonesien.
Daneben haben Rüsseltiere auch Einfluss auf ihr unmittelbares Biotop. Durch das Entrinden von Bäumen, Fressen von Blättern, Abknicken von Zweigen und Ästen, das Herausreißen von Büschen und kleinen Bäumen oder das Spalten größerer wirken sie zusammen mit anderen Megaherbivoren stark auf die Landschaft ein. Das gilt nicht nur für die heutigen Elefanten in den Savannen Ost- und Südafrikas sondern sicher auch für die Mammuts der Mammutsteppe.[8][9] Dabei ist dies offensichtlich ein Verhalten, das schon sehr früh im Rüsseltier-Stammbaum auftrat und bis ins frühe Miozän oder gar Oligozän zurückreicht, als u. a. mit den Deinotherien die ersten größeren Rüsseltiere mit sehr großen Stoßzähnen auftraten, deren überlieferten Abnutzungsspuren und Beschädigungen auf solche Handlungsweisen schließen lassen.[10]
Verbreitung
Die Rüsseltiere entstanden zweifellos in Afrika . Dies geschah zu einer Zeit, als dieser Kontinent noch nicht über Landbrücken mit anderen Erdteilen verbunden war. Solche Landbrücken entstanden erst im unteren Miozän vor mehr als 20 Millionen Jahren, als sich der nördlich gelegene Tethys-Ozean schloss und so eine Verbindung zum heutigen Eurasien entstand. Zu den ersten Auswanderern gehörten die die Echten Mastodonten und die Gomphotherien, die erstmals eurasischen Boden betraten. Einige Vertreter, wie z. B. Zygolophodon oder Gomphotherium, erreichten über Nordasien sogar den nordamerikanischen Kontinent und bildeten eigenständige Entwicklungslinien. Den ersten Auswanderern folgten die Deinotherien, allerdings verbreiteten sie sich nicht so weit, wie die Mastodonten und Gomphotherien, sondern blieben auf Eurasien beschränkt. Die erste Auswanderungswelle erfolgte dabei vor rund 20 bis 22 Millionen Jahren. Das Auftreten der Rüsseltiere außerhalb Afrikas wird als Proboscidean datum event bezeichnet, wobei dieses ursprünglich als singulär angesehene Ereignis nach neueren Untersuchungen aus mindesten sechs einzelnen Phasen bestand. Im Zuge der Bildung des Isthmus von Panama und der Entstehung einer geschlossenen amerikanischen Landmasse im Pliozän vor 3 Millionen Jahren kam es zum Großen Amerikanischen Faunenaustausch, wober letztendlich einige Vertreter der Gomphotherien auch Südamerika besiedelten.[11][12]
Die Rüsseltiere besiedelten somit einen Großteil der Alten und Neuen Welt, lediglich den Australischen Kontinent und die meisten weit vom Festland entfernten Inseln, wie Madagaskar und Neuguinea haben sie niemals erreicht. Dabei umfasste ihr Lebensraum sowohl tropische als auch arktische Lebensräume, wobei sie meistens Tiefländer, einige Arten wie Cuvieronius aber auch gebirgige Hochländer erschlossen.[12] Noch bis ins späte Pleistozän waren sie mit vier Familien über Amerika, Eurasien und Afrika verbreitet. Heute findet man sie nur noch in Afrika und Südasien in Form der Elefanten.
Stammesgeschichte
Evolutionstrends
Rüsseltiere sind eine relativ alte Ordnung der Säugetiere, erste Vertreter traten bereits im Paläozän vor mehr als 60 Millionen Jahren auf. Dabei kann die stammesgeschichtliche Entwicklung grob in drei Stufen eingeteilt werden, verbunden mit einer jeweiligen Auffächerung in zahlreiche Gattungen und Arten sowie Anpassung an unterschiedliche ökologische Nischen (adaptive Radiation). Generelle Trends in der Evolution der Rüsseltiere sind eine markante Größenzunahme - die ältesten Formen waren weniger als einen Meter groß, während spätere Formen bis zu mehr als 4 m Schulterhöhe erreichten -, Vergrößerung des Schädels, vor allem des Schädeldaches als Ansatzstelle für eine mächtige Nacken- und Kaumuskulatur, verbunden mit der Verkürzung des Kieferbereiches, Verkürzung des Halsbereiches, Ausbildung eines Rüssels, Hypertrophie der jeweils zweiten bzw. ersten Schneidezähne mit Ausbildung großer Stoßzähne ebenso wie die Tendenz zur Bildung großer Molaren bei gleichzeitigem Verlust der vorderen Prämolaren und weitgehend auch des vorderen Gebisses sowie die Änderung des Zahnaustausches vom für Säugetiere typischen vertikalen hin zum horizontalen Wechsel. Weiterhin bedeutend ist, dass frühere Rüsseltiere eher Blattfresser (browser) waren, während die späteren Formen stärker auf Grasnahrung (grazer) spezialisiert .waren.[13][14]
Erste Radiation
 Prodeinotherium
Prodeinotherium
 Mammut
MammutDie erste Radiation erfolgte vor 61 bis etwa 24 Millionen Jahren. Die Vertreter dieser urtümlichsten Rüsseltiere hatten noch deutlich bunodont aufgebaute Zähne mit maximal vier Querleisten am dritten Molaren, die jeweils einen hohen Zahnschmelzhöcker an den Enden aufwiesen. Einige Formen besaßen auch noch einen Eckzahn je Kieferast. Charakteristisch ist der hier noch vorkommende vertikale Zahnwechsel, so dass alle Zähne gleichzeitig in Gebrauch waren.[14][12]
Als ältestes Rüsseltier gilt derzeit Eritherium, das 2009 erstmals beschrieben wurde. Es war ein nur 3 bis 8 kg schweres Tier mit drei Leisten auf dem letzten Molar und lebte im nördlichen Afrika.[4] Phosphatherium, das 1996 in Marokko ausgegraben wurde, lebte vor etwa 55 Millionen Jahren und war kaum größer als ein Fuchs. Es hatte rein äußerlich wenig mit späteren Rüsseltieren gemeinsam, sein Zahnbau, der dem von Eritherium ähnelte aber stärker ausgebildete Leisten zwischen den Zahnhöckern besaß und so eine Tendenz zur Lophodontie aufweist, verrät jedoch die enge Verwandtschaft. Eine noch stärkere Tendenz zu diesen Zahnformen zeigen u. a. Numidotherium und Daouitherium, welche etwa gleichalt sind. Moeritherium aus dem Eozän Nordafrikas war ein weiteres frühes Mitglied der Rüsseltiere. Es war etwa so groß wie ein Tapir (etwa 110 cm), und besaß einen schweineähnlichen Kopf mit einer verlängerten Nasen-Oberlippe sowie leicht verlängerten Schneidezähnen im Ober- und Unterkiefer. Neben Elefantenmerkmalen trug der Schädel auch gemeinsame Merkmale mit dem der Seekühe. Des Weiteren zeichnete sich die Gattung durch einen sehr langen Körper aus. Mit Barytherium fand die erste enorme Körpergrößenzunahme innerhalb der Rüsseltier-Linie statt. Die Tiere erreichten eine Schulterhöhe von 2,5 bis 3 m und besaßen insgesamt acht Stoßzähne, je zwei pro Kieferast.[15][11]
Ebenfalls in die erste Radiationsphase gehören die Deinotherien (Deinotheriidae), die erstmals im mittleren Oligozän erschienen und eine frühe Abspaltung darstellen. Charakteristisch für diese Rüssseltiergruppe sind die Stoßzähne, die sich nur im Unterkiefer befanden und abwärts gebogen waren. Sie dienten als Werkzeuge zum Abschaben von Baumrinde. Die frühen Formen, wie Chilgatherium und Prodeinotherium waren noch relativ klein, während Deinotherium im Pliozän und Pleistozän teilweise bis zu 4 m Schulterhöhe erreichte. Sie stellten eine der ersten erfolgreichen Rüsseltiergruppen dar und starben in Europa im Mittleren Pliozän (Piacenzium), in Afrika im Unteren Pleistozän, vor rund einer Million Jahren aus.[16][17] Einige Autoren stellen die Zugehörigkeit der Deinotherien zu den Rüsseltieren aufgrund der Zahn- und Gebissmorphologie in Frage und möchten sie eher in näherer Verwandtschaft zu den Seekühen stellen,[18] diese Auffassung wird aber nur wenig geteilt.[19]
Palaeomastodon und Phiomia waren weitere sehr frühe Rüsseltiergattungen aus dem Eozän und Oligozän Nordafrikas. Sie gehören ebenfalls noch zu Vertretern der ersten Radiation, sind aber mit den späteren Rüsseltierarten schon deutlich näher verwandt als mit den früheren. Es bereitet derzeit noch Probleme, die frühen Formen mit diesen beiden Gattungen zu verbinden, da offensichtlich noch Zwischenglieder fehlen.[11][20] Wahrscheinlich umfasst Phiomia die Schwesterlinie zu den späteren Gomphotherien (Gomphotheriidae), während Palaeomastodon jene der Echten Mastodonten (Mammutidae) darstellt.[21]
Die Echten Mastodonten (Mammutidae) stellen die letzte Gruppe innerhalb der ersten Radiation dar. Ihre Entwicklungslinie begann laut molekulargenetischen Untersuchungen bereits vor wenigstens 26 Millionen Jahren.[22] Die Molaren sind zygodont mit maximal vier Schmelzleisten auf dem letzten Zahn. Der spezielle Aufbau der Backenzähne kennzeichnet sie als weitgehende Blattfresser. Weiterhin waren diese Rüsseltiere durch zwei Stoßzähne im Oberkiefer charakterisiert, während ältere Formen ebenfalls zwei kleinere Stoßzähne im Unterkiefer hatten, die im Laufe der weiteren Evolution erst reduziert und später verloren gingen. Die älteste Gattung stellt Eozygodon dar, während die bekannteste Gattung Mammut ist, zu der u. a. auch das Amerikanische Mastodon (Mammut americanum) gehörte, das zeitgleich mit den Vertretern der späteren Gattung Mammuthus lebte und gegen Ende der letzten Eiszeit im oberen Pleistozän ausstarb. Der Gattungsname Mammut führt häufig zur Verwirrung, da dessen Vertreter mit den eigentlichen Mammuts, deren Gattungsname Mammuthus lautet, nicht näher verwandt sind.[23][11]
Zweite Radiation
Die zweite Radiationsphase erfolgte im Miozän. Bei den Vertretern dieser Gruppe ist erstmals der horizontale Zahnwechsel nachweisbar, der durch die Verkürzung der Kieferknochen entstand. Bemerkenswert ist, dass die Echten Mastodonten zwar in die erste Radiationsphase gehören, die späteren Vertreter wie Mammut das Merkmal des horizontalen Zahnwechsels aber ebenfalls besaßen.[14] Weitere Merkmale in dieser Radiationsphase sind die deutlich hochkronigeren Backenzähne und die Entwicklung von bunodonten über lophodonte zu zygodonten Molaren mit einer teils deutlich dachartigen Struktur der einzelnen Zahnleisten, wobei sich die Zahl dieser Leisten des hintersten Molars auf sechs erhöhte.[14][12] Möglicher weise an der Basis der zweiten Radiation stand Eritreum aus dem späten Oligozän Nordostafrikas, das in seiner Zahnmorphologie noch zwischen Phiomia bzw. Palaeomastodon und den späteren Rüsseltieren vermittelte, aber schon den horizontalen Zahnwechsel aufwies.[21] Die wichtigsten Gruppen der zweiten Radiationsphase sind die Gomphotherien (Gomphotheriidae) und die Stegodonten (Stegodontidae), zwei Rüsseltierfamilien, die ursprünglich zusammen mit den Echten Mastodonten zur Überfamilie der Mastodonten (Mastodontoidea) zusammengefasst wurden.[24] Der Begriff Mastodon wird aber heute nur noch für die Echten Mastodonten gebraucht.[25]
Die bedeutendste Rüsseltierlinie stellten die Gomphotherien (Gomphotheriidae) dar. Diese waren eine sehr erfolgreiche Gruppe der Rüsseltiere, da sie sich aufgrund der starken Ausbreitung offener Landschaften im Miozän und der Umstellung auf Grasnahrung in zahlreiche Gruppen aufspalteten. So stellen sie heute fast die Hälfte aller bekannten Taxa, die sich in mehrere Unterfamilien aufteilen.[14] Allgemein handelte es sich bei den Gomphotherien um Rüsseltiere mit vier Stoßzähnen, je zwei im Ober- und im Unterkiefer. Ein weiteres Merkmal sind ein bunodonter bis lophodonter Zahnbau der Molaren, wobei die Milchzähne und die ersten beiden Dauermolaren drei Schmelzleisten besaßen - weswegen sie ursprünglich auch als trilophodonte Gomphotherien bezeichnet wurden -, während der letzte Backenzahn vier, fünf und mehr Rippen hatte.[26][12] Zur Unterteilung der Gomphotherien werden teilweise auch die Stoßzähne herangezogen. So wiesen die Gomphotheriinae mit Gomphotherium im Oberkiefer zwei deutlich nach unten gerichtete Stoßzähne auf, während jene des Unterkiefers langgestreckt und abgeflacht waren. Die Choerolophodontinae hatten dagegen kurze, deutlich reduzierte Unterkieferstoßzähne während die Rhynchotheriinae der ersten Gruppe ähnelten, aber seitlich abgeflachte Unterkieferstoßzähne aufwiesen. Diese wurden im späteren Verlauf, wie Stegomastodon und Cuvieronius zeigen, zusätzlich noch deutlich reduziert. Bemerkenswert sind die Amebelodontinae wie Platybelodon aus Asien und Amebelodon aus Nordamerika, die mit stark verlängerten und verbreiterten, schaufelartig umgebildeten Unterkieferstoßzähnen ausgestattet waren, während die Oberkieferstoßzähne nur eine geringe Größe aufwiesen. Einige der zu den Gomphotherien gezählten Arten überlebten bis ins späte Pleistozän.[11][12]
Deutlich weiter entwickelt als die Gomphotherien waren Tetralophodon und Anancus, die im mittleren bzw. im späten Miozän erstmals auftraten. Ursprünglich wurden sie als tetralophodonte Gomphotherien bezeichnet, da ihre Milchzähne und die vorderen Dauermolaren je vier Schmelzleisten besaßen. Gegenwärtig werden sie aber aufgrund modernerer Schädelmerkmale in die nähere Verwandtschaft zu den Stegodonten (Stegodontidae) und Elefanten (Elephantidae) gestellt und der Überfamilie Elephantoidea zugewiesen.[27] Beide Gattungen wiesen stark reduzierte Unterkieferstoßzähne auf, die teils nur noch im Milchgebiss ausgeprägt waren.
Die zweite große Gruppe innerhalb der zweiten Radiationsphase umfasste die Stegodonten, die sich im mittleren Miozän vor etwa 15 Mio. Jahren in Nordasien aus Gomphotherien mit bunodonten Molaren entwickelten.[28] Die älteste Form ist Stegolophodon. Aus dieser entstanden dann die späteren Gattungen, wie Stegodon, die typische gerippte und häufig sehr hochkronige Molaren besaßen. In der Regel hatten die Stegodonten nur obere Stoßzähne, während die unteren in den verkürzten Unterkiefern weitgehend reduziert oder nicht mehr ausgebildet waren. Diese Rüsseltiergruppe war über einen Großteil Eurasiens verbreitet, im späten Miozän und frühen Pliozän kam sie auch in Afrika vor während sie Amerika aber nicht erreichte.[29]
Dritte Radiation
Die dritte Radiationsphase begann im späten Miozän vor 7 Mio. Jahren und umfasste die Gruppe der Elefanten (Elephantidae), die einzige Rüsseltierfamilie, die bis heute überlebt hat. In dieser Phase wurde der Schädel weiter verkürzt und die Stirn erhöht. Die Molaren wurden weiter verlängert und waren nun deutlich lamellenartig aufgebaut, wobei die Anzahl der Lamellen von acht bis 30 variierte. Dabei waren die Lamellen deutlich flach ausgebildet und nicht mehr so prominent erhöht wie bei den vorhergehenden Rüsseltiergruppen. Außerdem kam es zu einem Rückgang der Zahnschmelzdicke je Lamelle. Alle diese Merkmale zeigen, dass diese Rüsseltiere sehr gut an Grasnahrung angepasst waren.[14]
Die älteste Form war Primelephas, welcher sich vor etwas mehr als 7 Mio. Jahren in Afrika aus Stegodon entwickelte, Aus ihm gingen die heutigen Gattungen Loxodonta und Elephas hervor.[11] Die Trennung der beiden Gruppen fand schon sehr früh in der Entwicklung von Primelephas statt und sollte nach Genanalysen bereits vor 7,6 Mio. Jahren abgeschlossen sein. Nur wenig später, vor 6,7 Mio. Jahren, spalteten sich Elephas und Mammuthus ab.[22] Während Loxodonta weitgehend auf dem afrikanischen Kontinent beschränkt blieb, erreichten Elephas und Mammuthus vor rund 3 Mio. Jahren auch Eurasien,[30] während einige Vertreter von Mammuthus als einzige Elefanten vor knapp 2 Mio. Jahren auch erstmals Amerika betraten.[31] Aufgrund der Verbreitung und Anpassung an das kühlere bis kalte Klima des nördlichen Eurasiens während des Pleistozäns zeichneten sich beide Gattungen durch eine recht hohe Radiation mit zahlreichen Arten aus. Während alle Elefanten eine sehr große Körpergröße aufwiesen, brachte Mammuthus mit dem Steppenmammut (Mammuthus trogontherii) die vermutlich größte bisher bekannt Rüsseltierart hervor, deren Schulterhöhe bei teilweise über 4,5 m lag.[14]
Im Neogen, besonders im Pleistozän, fand eine weltweite Verbreitung der Rüsseltiere auf alle Kontinente, außer Australien und Antarktika, mit zahlreichen Arten statt. Diese Verbreitung kann nur durch die Annahme ausgedehnter Wanderungen über Landbrücken, die sich vor ca. 20 Millionen Jahren zwischen Afrika und Eurasien gebildet haben, stattgefunden haben. Am Ende des Pleistozän, also bis vor etwa 12.000 Jahren lebten noch 7 Gattungen von Rüsseltien: Stegodon, Stegomastodon (inkl. Haplomastodon), Cuvieronius, Mammut, Mammuthus, Elephas und Loxodonta. Nur die letzten beiden überlebten bis heute. Dabei ging mit dem Aussterben der meisten Rüsseltiergattungen, aber auch anderer großer Säugetiere bis zum Beginn des Holozäns, möglicherweise die Ausbreitung des modernen Menschen (Homo sapiens) einher. Allerdings können neben dem Menschen auch die starken Klimaschwankungen der Warm- und Kaltzeiten als Ursache angesehen werden. Über die Gründe der sogenannten Quartären Aussterbewelle wird stark diskutiert.
Heute gibt es nur noch drei Arten der Rüsseltiere, die zwei Gattungen angehören. Dabei umfasst Loxodonta den Afrikanischen Elefanten (Loxodonta africana) und den Waldelefanten (Loxodonta cyclotis), während Elephas den Asiatischen Elefanten (Elephas maximus) einschließt.
Externe Systematik
Die nächsten lebenden Verwandten der Rüsseltiere sind die Seekühe. Diese bilden zusammen mit den ausgestorbenen Desmostylia und Embrithopoda das Taxon der Tethytheria. Ebenfalls relativ nahe verwandt sind die Schliefer, die die Außengruppe der Tethytheria bilden und mit diesen gemeinsam das Taxon der Paenungulata formen.
Man fasst diese Tiere heute mit einigen anderen Säugetierordnungen, die ebenfalls afrikanischen Ursprungs sind zur Überordnung der Afrotheria zusammen. Dazu gehören neben den oben erwähnten folgende noch lebende Ordnungen:
- Röhrenzähner (Tubulidentata)
- Rüsselspringer (Macroscelidea)
- Tenrekartige (Afrosoricida)
Interne Systematik
Die Ordnung der Rüsseltiere umfasst derzeit mehr als 160 Arten,[14] davon mehr als 130 aus Afrika, Asien und Europa, die sich auf rund 40 Gattungen und mehr als10 Familien verteilen. Einige Arten wurden erst in der Zeit nach dem Jahr 2000 entdeckt, darunter vor allem Vertreter der ältesten Rüsseltiere, wie das Eritherium oder das Daouitherium.[20] Generell lassen sich zwei große Formengruppen (Unterordnungen) unterscheiden: Plesielephantiformes und Elephantiformes, die anhand der Zahnmorphologie unterschieden werden. So besitzen Plesielephantiformes nur zwei Leisten (bilophodont) auf den ersten beiden Dauermolaren, während Elephantiformes drei, vier (tri- und tetralophodont) oder mehr besitzen. Dabei ist die Stellung der Deinotherien innerhalb der Plesielephantiformes problematisch, da sein zweiter Molar zwar biliphodont ist, dies aber möglicherweise kein ursprüngliches sondern ein abgeleitetes Merkmal darstellt.[32] Innerhalb der Elephantiformes gibt es noch die Teilordnung Elephantimorpha, deren Hauptmerkmal der horizontale Zahnwechsel ist. Vor allem stammesgeschichtlich ältere Gattungen sind teilweise in ihrer Stellung unsicher, ebenso einige fortgeschrittenere, wie Eritreum, das wohl am Übergang zu den Elephantimorpha steht.[21] Aber auch bei stammesgeschichtlich jüngeren Formen, vor allem der Gomphotherioidea gibt es noch taxonomische Unklarheiten. So wurden ursprünglich Anancus und Tetralophodon den Gomphotherien zugeordnet, in jüngere Zeit revidierte man diese Zuweisung wieder und stellte beide zu den moderneren Elephantoidea. Die Gliederung basiert auf den Bearbeitungen von Jeheskel Shoshani und Pascal Tassy 2005 unter Berücksichtigung neuer Ergebnisse.[27][12][33][11]
- Ordnung: Proboscidea Illiger 1811
-
-
-
- Familie incertae sedis: Eritherium Gheerbrant 2009
- Familie incertae sedis: Daouitherium Gheerbrant, Sudre, Cappetta, Iarochène, Amaghzaz und Bouya 2002
- Familie: Moeritheriidae Andrews 1906
-
- Moeritherium Andrews 1906
-
- Familie: Phosphatheriidae Gheerbrant, Sudre und Tassy 2005
-
- Phosphatherium Gheerbrant, Sudre und Cappetta 1996
-
-
-
- Plesielephantiformes: Shoshani 2001
-
-
- Familie incertae sedis: Arcanotherium Delmer 2009
- Familie: Numidotheriidae Shoshani und Tassy 1992
-
- Numidotherium Mahboubi et al. 1986
-
- Familie: Barytheriidae Andrews 1906
-
- Barytherium Andrews 1901
-
- Familie: Deinotheriidae Bonaparte 1841
- Unterfamilie: Chilgatheriinae Sanders, Kappelman und Rasmussen 2004
- Chilgatherium Sanders, Kappelman und Rasmussen 2004
- Unterfamilie: Deinotheriinae Sanders, Kappelman und Rasmussen 2004
- Deinotherium = (+ Prodeinotherium) Kaup 1829
- Unterfamilie: Chilgatheriinae Sanders, Kappelman und Rasmussen 2004
-
-
- Elephantiformes Tassy 1988
-
-
- Familie incertae sedis: Hemimastodon Pilgrim 1912
- Familie incertae sedis: Eritreum Shoshani, Walter, Abraha, Berhe, Tassy und Sanders 2006
- Familie: Palaeomastodontidae Andrews 1906
-
- Palaeomastodon Andrews 1901
-
- Familie: Phiomiidae Kalandadze und Rautian 1992
-
- Phiomia Andrews und Beadnell 1902
-
-
- Elephantimorpha Tassy und Shoshani 1997
- Überfamilie: Mammutoidea Hay 1922
- Familie: Mammutidae Hay 1922
-
- Mammut Blumenbach 1799
- Eozygodon Tassy und Pickford 1983
- Zygolophodon Vacek 1877
-
- Familie: Mammutidae Hay 1922
- Überfamilie: Gomphoterioidea Hay 1922
- Familie: Gomphotheriidae Hay 1922
- Unterfamilie: Choerolophodontinae Gaziry 1976
- Choerolophodon Gaudry und Lartet 1856
- Afrochoerodon McInnes 1942
- Unterfamilie: Gomphotheriinae Hay 1922
- Gomphotherium Burmeister 1837
- Unterfamilie: Amebelodontinae Barbour 1927
- Archaeobelodon Tassy 1984
- Protanancus Arambourg 1945
- Serbelodon Frick 1933
- Amebelodon Barbour 1927
- Platybelodon Borissiak 1928
- Unterfamilie: Rhynchotheriinae Hay 1922
- Rhynchotherium Falconer 1868
- Cuvieronius Osborn 1923
- Stegomastodon Pohlig 1912
- Gnathabelodon Barbour und Sternberg 1935
- Eubelodon Barbour 1914
- Sinomastodon Tobien, Chen und Li 1986
- Unterfamilie: Choerolophodontinae Gaziry 1976
- Familie: Gomphotheriidae Hay 1922
- Überfamilie: Elephantoidea Gray 1821
- Familie incertae sedis: Tetralophodon Falconer 1857; ursprünglich Unterfamilie Tetralophodontinae van der Maarel 1932 der Gomphotheriidae
- Familie incertae sedis: Anancus Aymard 1855; ursprünglich Unterfamilie Anancinae Hay 1922 der Gomphotheriidae
- Familie: Stegodontidae Osborn 1918
-
- Stegolophodon Schlesinger 1917
- Stegodon Falconer 1857
- Selenotherium Mackaye, Brunet und Tassy 2005
-
- Familie: Elephantidae Gray 1821
- Unterfamilie Stegotetrabelodontinae Aquirre 1969
- Stegotetrabelodon Petrocchi 1941
- Unterfamilie Elephantinae Gray 1821
- Primelephas Maglio 1970
- Loxodonta Anonymous 1827
- Elephas (+ Palaeoloxodon) Linnaeus 1758
- Mammuthus Brookes 1828
- Unterfamilie Stegotetrabelodontinae Aquirre 1969
- Überfamilie: Mammutoidea Hay 1922
-
-
Literatur
- E. Thenius: Grundzüge der Faunen- und Verbreitungsgeschichte der Säugetiere, 2.Auflage, Gustav Fischer Verlag, Stuttgart, 1980
- Ronald M. Nowak: Walker’s mammals of the world. 6. Auflage. Johns Hopkins University Press, Baltimore 1999, ISBN 0-8018-5789-9.
- Gregor. H-J., Kuhn, R., Storch, D.H.,: Deinotherium - ? ein Proboscidier ?, documenta naturae, no 130, München 2000, ISSN 0723-8428
- Jordi Augusti: Mammoths, Sabertooths and Hominids 65 Million Years of Mammalian Evolution in Europe, Columbia University Press, 2002. ISBN 0-231-11641-1
Einzelnachweise
- ↑ a b c d e Jeheskel Shoshani: Skeletal and basic anatomical features of elephants. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 9–20
- ↑ G. E. Weissengruber, F. K. Fuss, G. Egger, G. Stanek, K. M. Hittmair und G. Forstenpointner: The elephant knee joint: morphological and biomechanical considerations. Journal of Anatomy 208 (1), 2006, S. 59–72
- ↑ Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Theiss-Verlag, Stuttgart 2002, ISBN 3-8062-1734-3.
- ↑ a b Emmanuel Gheerbrant: Paleocene emergence of elephant relatives and the rapid radiation of African ungulates. PNAS 106 (26), 2009, S. 10717–10721
- ↑ Ekke W. Günther: Die Gebisse der Waldelefanten von Bilzingsleben. In: Dietrich Mania u. a. (Hrsg.): Bilzingsleben IV. Homo erectus – Seine Kultur und seine Umwelt. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 44, Berlin 1991, S. 149–174
- ↑ Jeheskel Shoshani, Robert C. Walter, Michael Abraha, Seife Berhe, Pascal Tassy, William J. Sander, Gary H. Marchant, Yosief Libsekal, Tesfalidet Ghirmai und Dietmar Zinner: A proboscidean from the late Oligocene of Eritrea, a ‘‘missing link’’ between early Elephantiformes and Elephantimorpha, and biogeographic implications. PNAS 103 (46), 2006, S. 17296–17301
- ↑ Cyrille Delmer: Reassessment of the generic attribution of Numidotherium savagei and the homologies of lower incisors in proboscideans. Acta Palaeontologica Polonica 54 (4), 2009, S. 561–580
- ↑ Ulrich Joger: Lebensweise und Ökologie moderner Elefanten: Sind Rückschlüsse auf den Waldelefanten und seine Umwelt möglich? In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 314–321
- ↑ Margret Bunzel-Drüke, Joachim Drüke und Henning Vierhaus: Der Einfluß von Großherbivoren auf die Naturlandschaft Mitteleuropas. Arbeitsgemeinschaft Biologischer Umweltschutz Kreis Soest e.V. 2001
- ↑ Friedrich Bachmayer und Helmuth Zapfe: Ein bedeutender Fund von Dinotherium aus dem Pannon von Niederösterreich. Analen des Naturhistorischen Museums zu Wien 80, 1976, S. 145–162
- ↑ a b c d e f g Jan van der Made: The evolution of the elephants and their relatives in the context of a changing climate and geography. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 340–360
- ↑ a b c d e f g María Teresa Alberdi, José Luis Prado, Edgardo Ortiz-Jaureguizar, Paula Posadas und Mariano Donato: Paleobiogeography of trilophodont gomphotheres (Mammalia: Proboscidea). A reconstruction applying DIVA (Dispersion-Vicariance Analysis). Revista Mexicana de Ciencias Geológicas 28 (2), 2011, S. 235–244
- ↑ Jeheskel Shoshani: Summary, conclusions, and a glimpse into the future. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 335–348
- ↑ a b c d e f g h Jeheskel Shoshani: Understanding proboscidean evolution: a formidable task. Tree 13, 1998, S. 480–487
- ↑ Jeheskel Shoshani, Robert M. West, Nicholas Court, Robert J. G. Savage und John M. Harris: The earliest proboscideans: general plan, taxonomy, and palaeoecology. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 57–75
- ↑ William Sanders, John Kappelmann und D. Tab Rassmussen: New large-bodied mammals from the late Oligocene site of Chilga, Ethiopia. Acta Palaeontologica Polonica 49 (3), 2004, S. 365–392
- ↑ Ursula B. Göhlich: Tertiäre Urelefanten aus Deutschland. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 362–372
- ↑ H. J. Gregor, R. Kuhn und D. H. Storch: Deinotherium? ein Proboscidier? Documenta Naturae 130, 2000, S. 1–141
- ↑ Kati Huttunen und Ursula Bettina Göhlich: A partial skeleton of Prodeinotherium bavaricum (Proboscidea, Mammalia) from the Middle Miocene of Unterzolling (Upper Freshwater Molasse, Germany). Geobios 35, 2002, S. 489–514
- ↑ a b Karol Schauer: Anmerkungen und Quellenangaben zur Evolutionstafel der Proboscidea in Afrika und Asien. In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, 2010, S. 630–650
- ↑ a b c Jeheskel Shoshani, Robert C. Walter, Michael Abraha, Seife Berhe, Pascal Tassy, William J. Sanders, Gary H. Marchant, Yosief Libsekal, Tesfalidet Ghirmai, und Dietmar Zinner: A proboscidean from the late Oligocene of Eritrea, a ‘‘missing link’’ between early Elephantiformes and Elephantimorpha, and biogeographic implications. PNAS 103 (46), 2006, S. 17296–17301
- ↑ a b Nadin Rohland, Anna-Sapfo Malaspinas, Joshua L. Pollack, Montgomery Slatkin, Paul Matheus und Michael Hofreiter: Proboscidean Mitogenomics: Chronology and Mode of Elephant Evolution Using Mastodon as Outgroup. PLOSBiology 5 (August), 2007, S. 1663–1671
- ↑ Heinz Tobien: Evolution of the zygodons with emphasis of dentition. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 76–84
- ↑ Henry Fairfield Osborn: Adaptive radiation and classification of the Proboscidea. Proceedings of the Natural American Society 7, 1921, S. 231–234
- ↑ Pascal Tassy: Who is who among the Proboscidea? In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 40–48
- ↑ María Teresa Alberdi, José Luis Prado, Edgardo Ortiz-Jaureguizar, Paula Posadas und Mariano Donato: Historical biogeogeography of trilophodont Gomphotheres (Mammalia, Proboscidea) reconstructed applying dispersion vicariance analysis. E. Díaz-Martínez und I. Rábano (Hrsg.:, 4th European Meeting on the Palaeontology and Stratigraphy of Latin America. Cuadernos del Museo Geominero, nº 8. Instituto Geológico y Minero de España, (Madrid) 2007, S. 9–14
- ↑ a b Jeheskel Shoshani und Pascal Tassy: Advances in proboscidean taxonomy & classification, anatomy & physiology, and ecology & behavior Quarternary International 126-128, 2005, S. 5–20
- ↑ Jon E. Kalb, David J. Froehlich und Gordon L. Bell: Palaeobiogeography of late Neogene African and Eurasian Elephantoidea. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 117-123
- ↑ Haruo Saegusa: Stegodontidae: evolutionary relationships. In: Jeheskel Shoshani und Pascal Tassy (Hrsg.): The Proboscidea. Evolution and palaeoecology of the Elephants and their relatives. Oxford, New York, Tokyo, 1996, S. 178–190
- ↑ Adrian M. Lister und Hans van Essen: The earliest Mammoth in Europe. Abstracts of the 18th International Senckenberg Conference 25. bis 30. April 2004 in Weimar
- ↑ Adrian Lister und Paul Bahn: Mammuts – Die Riesen der Eiszeit. Sigmaringen, 1997
- ↑ Jeheskel Shoshani, W. J. Sanders und Pascal Tassy: Elephants and other Proboscideans: a summary of recent findings and new taxonomic suggestions. In: G. Cavarretta et al. (Eds.): The World of Elephants - International Congress. Consiglio Nazionale delle Ricerche. Rom, 2001, S. 676–679
- ↑ Emmanuel Gheerbrant, Jean Sudre, Pascal Tassy, Mbarek Amaghzaz, Baâdi Bouya und Mohamed Iarochène: Nouvelles données sur Phosphatherium escuilliei (Mammalia, Proboscidea) de l’Éocène inférieur du Maroc, apports à la phylogénie des Proboscidea et des ongulés lophodontes.Geodiversitas 27 (2), 2005, S. 239–333
Weblinks
 Commons: Rüsseltiere – Sammlung von Bildern, Videos und Audiodateien
Commons: Rüsseltiere – Sammlung von Bildern, Videos und Audiodateien- The Paleobiology Database: Proboscidea








Wikimedia Foundation.