- Titanosauria
-
Titanosauria 
Hypothetische Skelettrekonstruktion von Argentinosaurus in einer Sonderausstellung des Naturmuseums Senckenberg
Zeitraum Kimmeridgium (Oberjura) bis Maastrichtium (Oberkreide) 155,6 bis 65,5 Mio. Jahre Fundorte - Europa, Asien, Afrika, Australien, Nordamerika, Südamerika
Systematik Dinosaurier (Dinosauria) Echsenbeckensaurier (Saurischia) Sauropodomorpha Sauropoden (Sauropoda) Titanosauria Wissenschaftlicher Name Titanosauria Bonaparte & Coria, 1993 
Die Titanosauria waren eine arten- und formenreiche Gruppe von sauropoden Dinosauriern. Sie lebten vor allem während der Kreidezeit, als sie die dominierende Sauropodengruppe waren, und starben als letzte Gruppe der Sauropoden erst vor etwa 65,5 Millionen Jahren an der Kreide-Tertiär-Grenze aus, zugleich mit allen anderen Nichtvogel-Dinosauriern. Fossilien wurden auf allen Kontinenten außer Antarktika entdeckt. Bisher sind über 40 Gattungen anerkannt, die mehr als ein Drittel der bekannten Arten der Sauropoden ausmachen. Der Bauplan dieser Pflanzenfresser ist wie bei allen Sauropoden durch einen tonnenförmigen Rumpf, einen langen Hals und einen im Verhältnis winzigen Kopf gekennzeichnet. Verschiedene Gattungen der Titanosauria wiesen Hautknochenplatten (Osteoderme) auf, die jedoch wahrscheinlich nicht zur Verteidigung dienten.[2] Funde von Eiern und Nestern geben Hinweise auf die Entwicklungs- und Reproduktionsbiologie. Diese Dinosaurier sind nach den Titanen der griechischen Mythologie benannt. Tatsächlich waren unter ihnen einige der größten und schwersten Landtiere aller Zeiten, beispielsweise Argentinosaurus oder Paralititan, es sind jedoch auch Zwergformen wie Magyarosaurus bekannt.[3]
Die meisten Gattungen sind aufgrund des sehr lückenhaften Fossilberichts nur schlecht dokumentiert. So sind Schädelknochen und Partien des Restskeletts, die sich bei der Entdeckung noch im anatomischen Verbund befinden, sehr selten. Die Verwandtschaftsbeziehungen innerhalb der Gruppe werden erst seit 2001 ernsthaft diskutiert und bleiben bis heute stark umstritten.[4] Zusammen mit den Brachiosauriden und verwandten Gattungen bilden die Titanosauria die Gruppe Titanosauriformes, welche zu den Macronaria zählt.[3]
Inhaltsverzeichnis
Merkmale
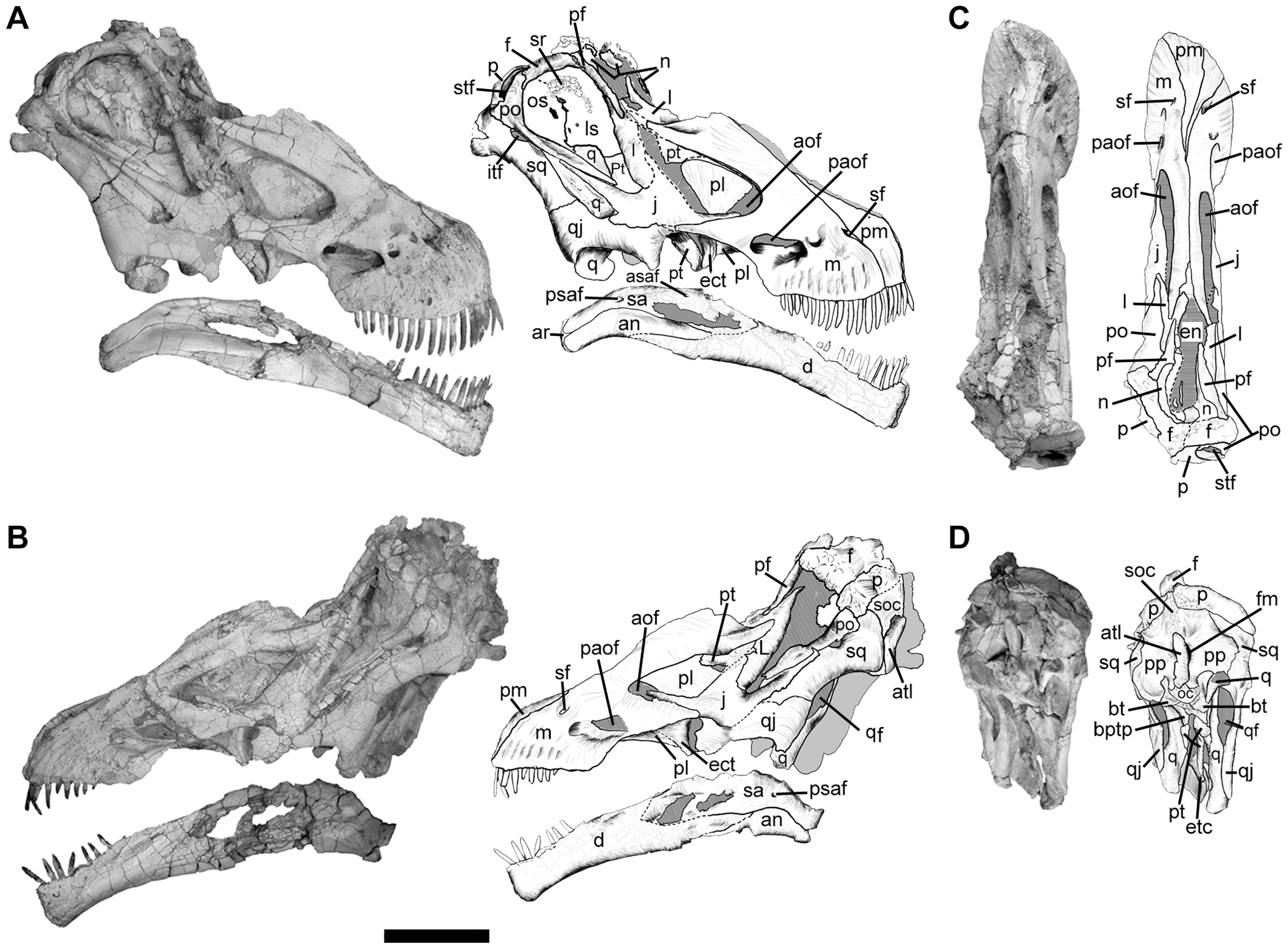
Schädel
Nahezu vollständige Schädel sind lediglich von Rapetosaurus, Nemegtosaurus, Quaesitosaurus und Tapuiasaurus[5] bekannt. Diese Schädel ähnelten in ihrer Form denjenigen der Diplodociden: So war die Schnauze verlängert, während sich die Nasenöffnungen weit oben am Schädel auf Höhe der Augenhöhlen befanden. Dennoch zeigten diese Schädel gemeinsame Merkmale, die bei Diplodociden und anderen Sauropodengruppen fehlten. So grenzt beispielsweise das Schuppenbein (Squamosum) nicht an das Supratemporalfenster, ein Schädelfenster auf der hinteren Oberseite des Schädels, an. Des Weiteren ist die Schnauze, also das Zwischenkieferbein (Prämaxillare) sowie die vorderen Teile des Ober- und Unterkiefers, mit zahlreichen Öffnungen (Foramina) durchzogen. Bislang ist jedoch zu wenig Schädelmaterial bekannt, um Merkmale des Schädels zur Definition von Gruppen innerhalb der Titanosauria zu verwenden.[6][7][8]
Nemegtosaurus besaß wahrscheinlich 13 Zähne auf jeder Seite der Ober- und Unterkiefer, was einer Gesamtanzahl von 52 Zähnen entspricht.[8] Die Zahnkronen waren lang und dünn und ähnelten den noch dünneren, stiftartigen Zahnkronen der Diplodociden; bei beiden Gruppen hat sich diese Morphologie jedoch unabhängig voneinander (konvergent) entwickelt. So weisen ursprünglichere Sauropoden wie Camarasaurus oder Omeisaurus spatelförmige Zahnkronen auf, während ursprünglichere Titanosauriformes wie Brachiosaurus eine Zwischenform zwischen den ursprünglicheren, spatelförmigen Zahnkronen und den dünneren Zahnkronen der Titanosauria zeigen.[4][6]
Wirbel
Die Wirbel der Titanosauria waren generell breiter und niedriger als bei anderen Sauropodengruppen.[9] Die vorderen Schwanzwirbel aller Titanosauria außer Opisthocoelicaudia waren procoel, das heißt, auf der Vorderseite konkav. Die seitlichen Aushöhlungen (Pleurocoele) der Rückenwirbel waren im Gegensatz zu denen anderer Sauropoden augenförmig.[4][10] Bisher fehlen Funde von zusammenhängenden Wirbelsäulen, weshalb die generelle Anzahl der Hals-, Rücken- und Schwanzwirbel unbekannt ist. Viele stärker abgeleitete Titanosauria zeigten jedoch einen zusätzlichen, sechsten Kreuzbeinwirbel. So wiesen andere Neosauropoden typischerweise fünf Kreuzbeinwirbel auf; bei sehr ursprünglichen Sauropoden wie Barapasaurus waren es nur vier. Des Weiteren besaßen einige abgeleitetere (modernere) Titanosaurier wie Opisthocoelicaudia einen deutlich kürzeren Schwanz. Bestand der Schwanz bei ursprünglicheren Sauropoden aus etwa 50 und bei Diplodociden aus etwa 80 Schwanzwirbeln, waren es bei diesen abgeleiteten Titanosauria nur noch 35.[6][4][11]
Die Wirbel zeichneten sich insbesondere bei abgeleiteteren Titanosauria wie den Saltasauriden durch eine Reduzierung der mechanischen Verbindungselemente aus, was zu einer größeren Flexibilität der Wirbelsäule führte. Dieser Trend zeigte sich sogar in den Schwanzwirbeln. Besonders bezeichnend ist das Fehlen der Hyposphen-Hypantrum-Verbindungen der Rückenwirbel bei allen Titanosauria außer einigen ursprünglichen Formen wie Andesaurus und Phuwiangosaurus. Diese Verbindungselemente führten bei anderen Sauropoden zu einer zusätzlichen Stabilisierung der Wirbelsäule und unterstützten wahrscheinlich eine zunehmende Körpergröße.[9]
Die vor dem Kreuzbein gelegenen Wirbel (Präsakralwirbel) und gelegentlich die Kreuzbeinwirbel sind mit einer komplexen, bienenwabenartigen Struktur aus zahlreichen Kammern durchzogen, die teilweise vom Luftsacksystem des Tieres ausgefüllt wurden und das Gewicht der Wirbelsäule reduzierten. Eine derartig komplexe interne Struktur der Wirbel findet sich auch bei den Diplodociden und bei Mamenchisaurus, während weniger abgeleitete Sauropoden ein einfacheres Kammersystem zeigten. Die Evolution dieses komplexen Kammersystems in verschieden Gruppen der Sauropoden steht vermutlich mit der zunehmenden Körpergröße und Halslänge im Zusammenhang.[6][12]
Schultergürtel, Becken und Gliedmaßen
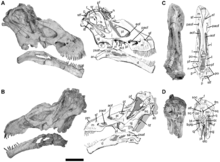
 Handskelett von Diamantinasaurus. Dieser Fund ist der bisher einzige Nachweis einer Handkralle bei Titanosauriern (rechts im Bild). Aus Hocknull und Kollegen, 2009.[1]
Handskelett von Diamantinasaurus. Dieser Fund ist der bisher einzige Nachweis einer Handkralle bei Titanosauriern (rechts im Bild). Aus Hocknull und Kollegen, 2009.[1]
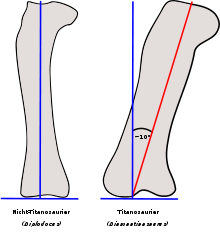 Oberschenkelknochen von einem Nicht-Titanosaurier (links, Diplodocus) und einem Titanosaurier (rechts, Diamantinasaurus) im Vergleich. Die blauen Linien markieren die Horizontale und die Vertikale, die rote Linie markiert die Schaftachse. Während bei anderen Sauropoden wie Diplodocus die Schaftachse der vertikalen Ausrichtung entspricht, war der Oberschenkelknochen der Titanosaurier leicht abgespreizt und steht somit in einem Winkel von etwa 10° zur vertikalen Ausrichtung.
Oberschenkelknochen von einem Nicht-Titanosaurier (links, Diplodocus) und einem Titanosaurier (rechts, Diamantinasaurus) im Vergleich. Die blauen Linien markieren die Horizontale und die Vertikale, die rote Linie markiert die Schaftachse. Während bei anderen Sauropoden wie Diplodocus die Schaftachse der vertikalen Ausrichtung entspricht, war der Oberschenkelknochen der Titanosaurier leicht abgespreizt und steht somit in einem Winkel von etwa 10° zur vertikalen Ausrichtung.Im Appendikulärskelett – dem Schultergürtel, dem Becken und den Gliedmaßen – findet sich bei den Macronaria und insbesondere bei den Titanosauria ein bedeutender Teil der für die Klassifikation wichtigen Merkmale.[11] Viele dieser Merkmale hängen wahrscheinlich mit der Evolution einer weiteren Beinstellung innerhalb der Titanosauria zusammen: So standen die Beine nicht säulenartig gerade unter dem Körper, wie bei primitiveren Sauropoden-Gruppen, sondern waren von vorne betrachtet leicht nach außen gebogen. Somit berührten die Füße den Boden in einigem Abstand von der Mittellinie des Körpers, und standen nicht, wie bei primitiveren Sauropoden-Gruppen, nahe an oder auf der Mittellinie. Analog können fossile Fährtenfolgen von Sauropoden in zwei Gruppen eingeordnet werden, den „schmalspurigen“ Typ, Parabrontopodus, der wahrscheinlich die ursprüngliche Konstellation primitiverer Sauropodengruppen zeigt, sowie den „breitspurigen“ Typ, Brontopodus, der die Konstellation bei Titanosauriern und evtl. anderen Macronariern widerspiegelt.[13]
Der Schultergürtel zeichnete sich durch ein im Vergleich mit anderen Sauropoden verlängertes Rabenbein (Coracoid) und ein größeres, mondsichelförmiges Brustbein (Sternum) aus, was zu einem insgesamt breiteren Brustkorb führte. Die Gliedmaßen waren generell robust und im Verhältnis kürzer als bei anderen Sauropoden[14]. Verschiedene Merkmale der Vorderbeine der Titanosauria fanden sich bei anderen Gruppen der Saurischia, fehlen jedoch bei den übrigen Sauropodengruppen. Dabei handelt es sich um Rückentwicklungen von Merkmalen, die sich früh in der Evolution der Sauropoden herausbildeten und mit der Evolution von säulenartigen Beinen in Verbindung standen[11]. So zeigte der Oberarmknochen (Humerus) einen ausgeprägten Deltopectoralkamm sowie geteilte und anterior (nach vorne) erweiterte distale (untere) Gelenkknorren. Des Weiteren erstreckte sich ein Knochensporn am oberen Ende der Elle (Olekranon) über die Gelenkfläche der Elle. Diese Merkmale lassen auf nach außen gebeugte Vorderbeine schließen. Der aus Elle und Speiche (Radius) bestehende Unterarm war zudem extrem robust – beispielsweise betrug die Dicke der Speiche an ihrem oberen (proximalen) Ende mindestens ein Drittel ihrer gesamten Länge, ein gemeinsam abgeleitetes Merkmal (Synapomorphie) der Titanosauria[4]. Die Mittelhandknochen waren kürzer und robuster als die stark verlängerten Mittelhandknochen ursprünglicherer Macronaria wie Brachiosaurus. Die Handwurzelknochen (Carpalia) scheinen komplett gefehlt zu haben, während von den Fingern lediglich extrem reduzierte Reste einiger Fingerglieder (Phalangen) übrig geblieben waren. Krallen (Unguals) am Vorderfuß wurden bislang bei keinem Titanosauria außer Diamantinasaurus[1] nachgewiesen.[15][6]
Das Darmbein (Ilium) der meisten Sauropoden zeigte auf der Oberseite der vorderen (präacetabularen) Hälfte einen markanten, gerundeten Kamm. Bei den Titanosauriformes wurde dieser Kamm höher und keulenartig, sodass das Darmbein seinen höchsten Punkt weit vor der Hüftgelenkpfanne (Acetabulum) zeigte. Bei einigen Titanosauria, beispielsweise bei Neuquensaurus oder Alamosaurus, war diese Keule nach außen gedreht und bildete eine annähernd horizontale Plattform.[10][16] Das Schambein (Pubis) war bei vielen Arten deutlich länger als das Sitzbein (Ischium), im Gegensatz zu anderen Sauropoden, bei denen dieses Größenverhältnis meist umgekehrt war.[10]. Das kürzere Sitzbein der Titanosauria war meist breiter als das anderer Sauropoden.[4]
Der Oberschenkelknochen (Femur) war nicht senkrecht orientiert wie bei anderen Sauropoden, sondern leicht nach außen abgespreizt, was zu der typischen weiteren Beinstellung führte. Dies zeigt sich zum einen an dem nach innen gebogenen proximalen (oberen) Drittel dieses Knochens. Zum anderen standen die distalen (unteren) Gelenkknorren nicht senkrecht zur Schaftachse, wie bei anderen Sauropoden, sondern waren relativ zur senkrechten Ausrichtung um 10° geneigt. Im Querschnitt war der Schaft des Oberschenkelknochens der meisten Sauropoden mediolateral breiter als anteroposterior; von vorne betrachtet war der Knochen also breiter als von der Seite betrachtet, der Querschnitt erscheint als Ellipse. Bei den Titanosauria war diese Ellipse deutlich exzentrischer als bei den übrigen Sauropodengruppen. Diese Anpassung wirkte vermutlich den durch die weitere Beinstellung verursachten größeren Beugungsmoment entgegen.[13][6]
Die Füße trugen im Gegensatz zu den Händen gekrümmte Krallen. Bisher sind lediglich drei vollständige und artikulierte Fußskelette gefunden worden, die in ihrem Aufbau stark variieren. Die Anzahl der Zehenglieder (Phalangen) jeder der fünf Zehen lässt sich durch sogenannte Phalangenformeln beschreiben: So zeigt Epachthosaurus eine Phalangenformel von 2-2-3-2-0, während sie bei Opisthocoelicaudia 2-2-2-1-0 und bei einem kürzlich in Patagonien entdeckten Skelett 2-2-2-2-0 beträgt.[17]
Paläobiologie
Riesenwuchs und Inselverzwergung
 Dieses Fragment eines Oberschenkelknochens des gigantischen Titanosauriers Argentinosaurus im Museo de La Plata (Argentinien) misst 1,18 Meter Länge. Ein vollständiger Oberschenkelknochen wäre 2,5 Meter lang gewesen.
Dieses Fragment eines Oberschenkelknochens des gigantischen Titanosauriers Argentinosaurus im Museo de La Plata (Argentinien) misst 1,18 Meter Länge. Ein vollständiger Oberschenkelknochen wäre 2,5 Meter lang gewesen.Sauropoden als die größten Landtiere aller Zeiten waren im Durchschnitt schätzungsweise zwischen 15 und 40 Tonnen schwer, während das Gewicht nur sehr weniger Arten auf unter 5 Tonnen geschätzt wird. Besonders riesige Formen entwickelten sich mehrfach unabhängig voneinander innerhalb verschiedener Entwicklungslinien, sowohl bei basalen Formen wie beispielsweise Turiasaurus, bei Diplodociden wie beispielsweise Amphicoelias, bei Brachiosauriden wie beispielsweise Sauroposeidon und – wahrscheinlich ebenfalls unabhängig voneinander – bei verschiedenen Formen innerhalb der Titanosauria. Die größten bekannten Titanosauria schließen Argentinosaurus mit ein, der auf eine Länge von 30 Metern[18] und ein Gewicht von 60 bis 88 Tonnen[19] geschätzt wird. Der ägyptische Paralititan wird auf 26 Meter Länge[18] und 59 Tonnen Gewicht[20] geschätzt, während die ebenfalls riesenhaften Formen Antarctosaurus giganteus mit schätzungsweise 70 Tonnen[19] und Puertasaurus nur wenig bekannt sind. Während die meisten der größten Sauropoden nur durch sehr fragmentarische Überreste bekannt sind, wurden von dem schätzungsweise 32 bis 34 Meter langen Futalognkosaurus etwa 70 % des Skeletts entdeckt, was ihn zu dem am vollständigsten überlieferten gigantischen Sauropoden macht.[21][22]
Im Kontrast zu diesen Riesenformen schließen die Titanosauria auch einige der kleinsten bekannten Sauropoden mit ein. So erreichte der rumänische Magyarosaurus lediglich eine Länge von etwa fünf Metern.[23] Diese vergleichsweise geringe Größe wird auf Inselverzwergung zurückgeführt – so war Europa in der Kreidezeit ein Inselarchipel. Bei Rapetosaurus aus Madagaskar sowie bei Ampelosaurus aus Westeuropa könnte es sich ebenfalls um Inselformen gehandelt haben.[22]
Ernährung
Die dünnen Zahnkronen der abgeleiteteren Titanosauria sowie der Diplodociden lassen darauf schließen, dass diese Sauropoden ihre Nahrung nicht im Mund zerkleinern konnten, anders als frühe Macronaria und ursprünglichere Sauropoden, die wahrscheinlich mit ihren spatelförmigen Zahnkronen Nahrung zu einem gewissen Grad verarbeiten konnten.[24] Dinosaurierfaunen mit jeweils mehreren zeitgleich lebenden Titanosauria sind aus Südamerika, Indien, Madagaskar und Europa[25] bekannt. Dies legt nahe, dass verschiedene Titanosauria zur Vermeidung von Konkurrenz unterschiedliche ökologische Nischen für die Nahrungssuche besetzten.[26]
Prasad und Kollegen (2005) berichten über Koprolithen (versteinerter Dung) aus der Oberkreide Indiens, der zusammen mit den Überresten von Titanosauriern entdeckt wurde. Diese Koprolithen enthalten die Überreste von Koniferen, zweikeimblättrigen Pflanzen, Palmen sowie verschiedene Gruppen der Gräser, die Nahrung der Titanosaurier bestand demnach aus einer großen Bandbreite verschiedener Pflanzen. Die Zuordnung dieser Koprolithen zu den Titanosauria bleibt jedoch unsicher.[27][22]
Funktion des Schwanzes
Die Schwänze von Sauropoden aus der Gruppe der Diplodocidae endeten in einem charakteristischen Peitschenschwanz, der eventuell zur Verteidigung oder zur Lärmerzeugung verwendet wurde. Einige Autoren sehen die erhöhte Flexibilität des Schwanzes sowie die wie bei Diplodociden bikonvex geformten hinteren Schwanzwirbel der Titanosaurier als Hinweis darauf, dass auch Titanosaurier ein solches peitschenartiges Schwanzende besessen haben könnten.[9] Andere Autoren widersprechen dieser Hypothese und geben an, dass die Serie dieser bikonvexen Wirbel deutlich kürzer als bei den Diplodociden war, weshalb man nicht von einem Peitschenschwanz sprechen könne. Die Hypothese eines peitschenartigen Schwanzes tauchte bereits in den 1980er-Jahren auf, war damals jedoch der fehlerhaften Einordnung der Titanosaurier als enge Verwandte der Diplodociden geschuldet.[28]
Bei den Saltasauriden war der Schwanz mit nur 35 Wirbeln deutlich verkürzt. Dieser verkürzte Schwanz könnte nach Ansicht einiger Forscher als eine Art drittes Hinterbein gedient haben, während sich die Tiere zum Fressen oder zur Paarung aufrichteten.[11]
Eier und Nester
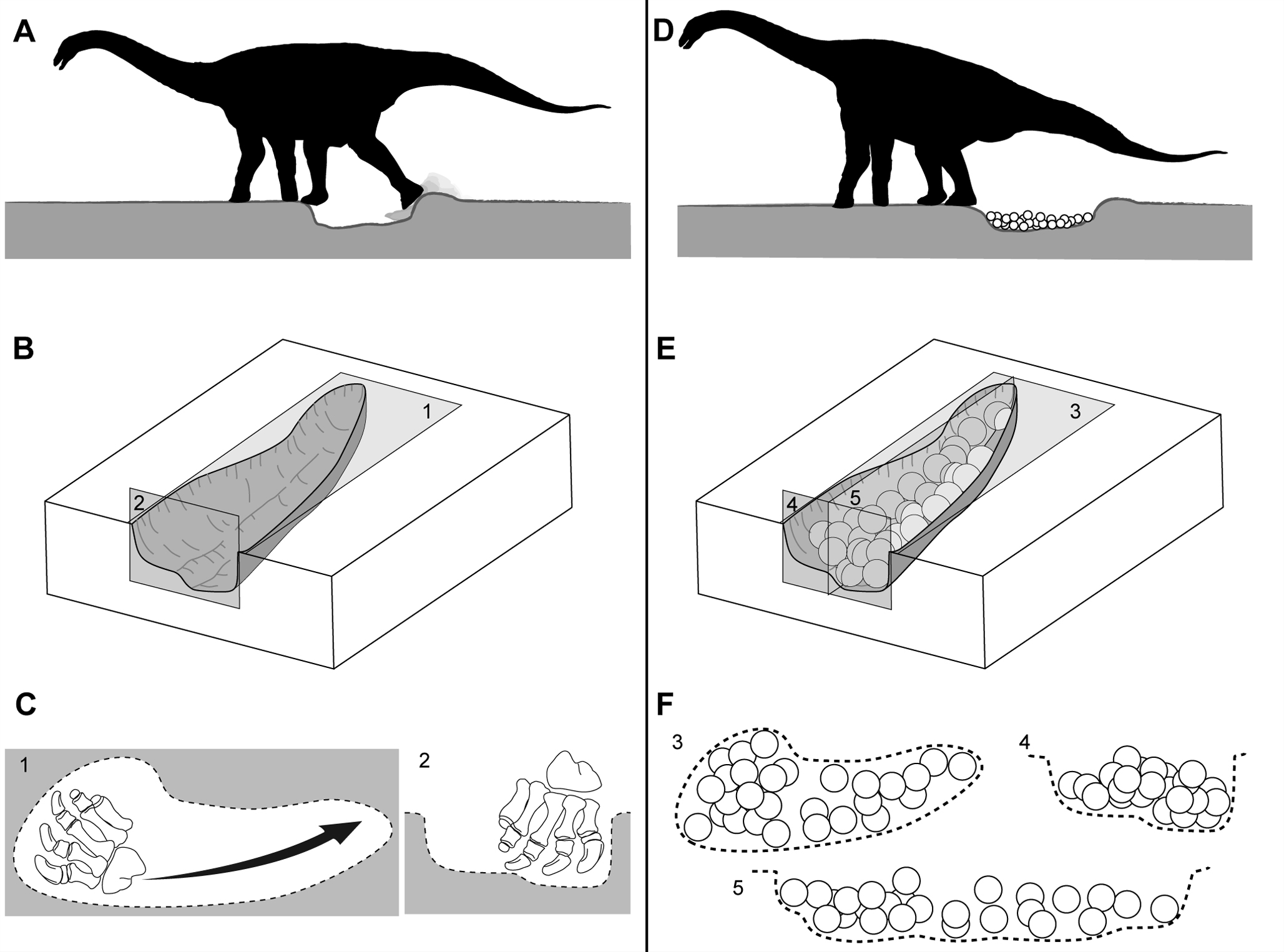
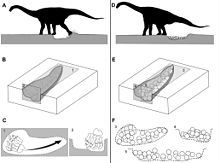 Titanosaurier hoben Nestgruben mit ihren flexiblen Hinterfüßen aus. Die Gelege bestanden typischerweise aus 20 bis 40 Eiern. Abbildung aus Vila und Kollegen, 2010[29]
Titanosaurier hoben Nestgruben mit ihren flexiblen Hinterfüßen aus. Die Gelege bestanden typischerweise aus 20 bis 40 Eiern. Abbildung aus Vila und Kollegen, 2010[29] Megaloolithus-Nestgrube (clutch) aus Pinyes (Spanien). Abbildung aus Vila und Kollegen, 2010[29]
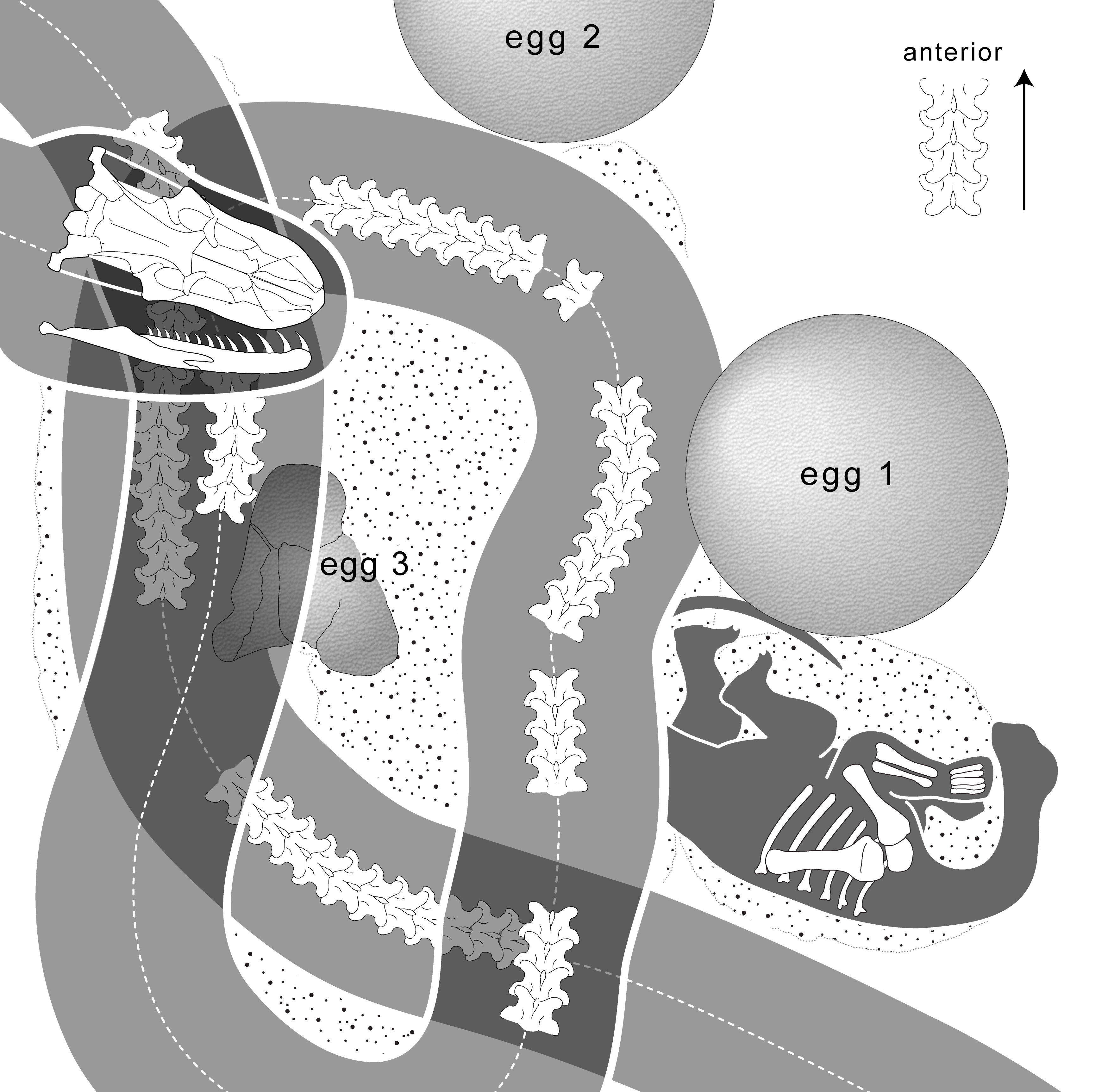
Megaloolithus-Nestgrube (clutch) aus Pinyes (Spanien). Abbildung aus Vila und Kollegen, 2010[29] Ein vermutlicher Nesträuber, die Schlange Sanajeh indicus. Rechts ein assoziiertes Sauropodenei, darunter ein frisch geschlüpfter oder noch embryonaler Titanosaurier. Aus Wilson und Kollegen, 2010[2]
Ein vermutlicher Nesträuber, die Schlange Sanajeh indicus. Rechts ein assoziiertes Sauropodenei, darunter ein frisch geschlüpfter oder noch embryonaler Titanosaurier. Aus Wilson und Kollegen, 2010[2]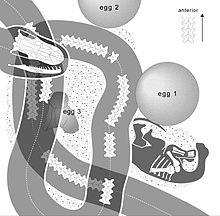 Das Diagramm verdeutlicht die Position der Schlange im oben wiedergegebenen Fossil. Ei 2 fehlt in der obigen Abbildung des Originalfossils. Aus Wilson und Kollegen, 2010[2]
Das Diagramm verdeutlicht die Position der Schlange im oben wiedergegebenen Fossil. Ei 2 fehlt in der obigen Abbildung des Originalfossils. Aus Wilson und Kollegen, 2010[2]Bis heute wurden zahlreiche Fundorte mit fossilen Eiern vom Typ (Oogenus) Megaloolithus entdeckt, die traditionell sauropoden Dinosauriern zugeschrieben werden. Diese Eier zeichnen sich durch eine rundliche Form, eine relativ dicke Eierschale, die aus einer einzelnen Schicht aus Calcit besteht, sowie eine tuberkelartige Oberfläche dieser Eierschale aus. Gut untersuchte Fundorte gibt es vor allem in Gesteinen der Oberkreide im südlichen Frankreich, im nördlichen Spanien, in Indien sowie in Südamerika. Mit der Entdeckung der Fundstelle Auca Mahuevo 1997 im nördlichen Patagonien, einer riesigen Nestkolonie mit tausenden Megaloolithus-Eiern, konnte die Zuordnung zumindest dieser Nestkolonie zu den Titanosauria bestätigt werden. So fanden sich in einigen Eiern gut erhaltene Skelette von Embryonen, die einige Synapomorphien der Titanosauria zeigen.[30] Viele Autoren schreiben andere Megaloolithus-Nestkolonien aufgrund der Ähnlichkeit mit den Funden aus Auca Mahuevo ebenfalls Titanosauriern zu.[29] Andere Autoren betonen jedoch, dass die Nachweise nicht ausreichen, um alle Megaloolithus-Funde den Titanosauria zuzuordnen. So fanden sich in Rumänien Megaloolithus-Eier in der Nähe von Überresten neugeborener Hadrosaurier der Art Telmatosaurus transylvanicus. Somit besteht die Möglichkeit, dass einige Megaloolithus-Funde tatsächlich zu anderen Gruppen gehörten.[30][31]
Die Fundorte der Eier in Auca Mahuevo erstrecken sich über sechs unterschiedliche stratigraphische Schichten, was vermuten lässt, dass Titanosauria an diesem Ort während mindestens sechs verschiedenen Zeiträumen genistet haben. Die große Anzahl der Gelege lässt auf ein Herdenleben dieser Sauropoden zumindest zur Brutsaison schließen. Obwohl Brutpflege bei anderen Dinosauriergruppen wie den Ornithopoden und den Theropoden nachgewiesen wurde und auch die nächsten heute lebenden Verwandten der Sauropoden, die Vögel und die Krokodile, Brutpflege betreiben, vermuten viele Forscher, dass die Sauropodennester nach der Eiablage nicht weiter betreut wurden. Darauf weist die riesige Größe der erwachsenen Tiere sowie die hohe Konzentration an Nestern auf relativ kleiner Fläche in Auca Mahuevo hin. Zudem fehlen in Auca Mahuevo Hinweise auf von den Sauropoden zertrampelte Oberflächen (Dinoturbation).[30]
Die Eier haben typischerweise 13 bis 15 Zentimeter Durchmesser, während die Eischale ca. 1,3 mm dick ist. Sie finden sich in Ansammlungen aus meist 20 bis 40 Eiern, die in bis zu drei Lagen übereinandergestapelt sind. Das eigentliche Nest ist dabei jedoch nur selten erhalten, weshalb lediglich von Gelegen (engl. clutches) und nicht von Nestern gesprochen wird. Einige Gelege zeigen dennoch Hinweise auf die Architektur des Nestes: Sie befinden sich innerhalb länglicher, elliptischer bis nierenförmiger Eindrücke in einer Grundschicht aus Sandstein, die etwa 100 bis 140 Zentimeter lang und 10 bis 18 Zentimeter tief sind. Ein Wall aus strukturlosem Sandstein um die Eindrücke herum ist wahrscheinlich bei der Aushebung dieser Nester durch die Titanosaurier entstanden.[30] Ähnliche längliche Eindrücke finden sich in einigen anderen Megaloolithus-Nestkolonien in Südamerika, Europa und Indien. Eine neuere Studie vermutet, dass Titanosaurier diese Gruben mit ihren flexiblen Hinterfüßen durch Schürfen ausgehoben haben.[29]
Die meisten Studien über Megaloolithus-Eier lassen vermuten, dass die Eier vergraben wurden.[29] Diese Interpretation wird durch die Anzahl der Poren in der Eierschale und die daraus resultierende Durchlässigkeit für Wasserdampf (GH2O) unterstützt. So sind die Eier heutiger Vögel meistens der Atmosphäre ausgesetzt, weshalb ihre Eier relativ wenige Poren zeigen, um einen möglichen Wasserverlust zu vermeiden. Die Eier heutiger Reptilien werden dagegen häufig in der Erde oder in Vegetation vergraben und zeichnen sich durch entsprechend zahlreiche Poren aus. Viele Megaloolithus-Eierschalen zeigen eine Wasserdampf-Durchlässigkeit, die deutlich höher als bei heutigen Vögeln, aber etwas geringer als bei heutigen Reptilien ist. Beispielsweise zeigen bei Pinyes im nördlichen Spanien gefundene Eier eine zehn Mal größere Wasserdampf-Durchlässigkeit als heutige Vogeleier. Eine Ausnahme scheinen die Eier aus Auca Mahuevo zu bilden, für die lediglich eine zwei Mal größere Wasserdampf-Durchlässigkeit relativ zu Vogeleiern berechnet wurde.[32] Die Auca-Mahuevo-Eier könnten daher, anders als andere Megaoolithus-Eier, in offenen Nestern bebrütet worden sein. Dies wird dadurch unterstützt, dass die Nestgruben dieser Fundstelle nicht mit dem Sandstein ausgefüllt sind, in welchem die Nester hineingegraben worden wären, sondern mit dem erst nachträglich durch eine Überschwemmung eingetragenen Schlammstein.[30] Grellet-Tinner und Kollegen (2004) vermuten, dass die Titanosaurier von Auca Mahuevo ihre Nester mit Pflanzenmaterial bedeckten. Darauf lässt die tuberkelartige Oberfläche der Eierschalen schließen, die wahrscheinlich die Funktion hatte zu verhindern, dass Partikel die Poren der Eierschale blockierten. Da die Erhebungen in einem Abstand zwischen 0,6 und 1,6 Millimeter voneinander stehen, konnten sie lediglich Partikel fern halten, die größer waren als 0,6 Millimeter, weshalb nur Pflanzenmaterial als Bedeckung in Frage kommt. Diese Interpretation wird durch die Entdeckung von Kohlenstoff-Resten in den Nestgruben unterstützt, die vermutlich von Pflanzenmaterial herrühren.[33][31]
In der Oberkreide Indiens fanden sich Hinweise auf einen Nesträuber: So wurde das Skelett der 3,5 Meter langen Schlange Sanajeh in Assoziation mit drei Eiern und dem vorderen Teil des Rumpfes eines entweder frisch geschlüpften oder embryonalen Titanosauriers gefunden. Eines der Eier ist vom Körper der Schlange umwunden und zerdrückt, während die anderen beiden Eier intakt sind. Hieraus schließen Forscher, dass das Titanosaurier-Skelett zu demselben Tier gehören könnte, das zuvor aus dem nun zerdrückten Ei geschlüpft ist. Die Überreste des Sauropoden gehörten zu einem etwa 0,5 Meter langen Tier.[2]
Individualentwicklung
In einigen Eiern von Auca Mahuevo wurden annähernd vollständige und sich im anatomischen Verbund befindliche Embryonenschädel entdeckt. Diese Schädel stellen die derzeit vollständigsten bekannten Titanosauria-Schädel dar und geben Einblicke in die Individualentwicklung dieser Sauropoden. Die Embryonenschädel waren breit und von der Seite betrachtet annähernd dreieckig. Sie zeigten große, runde Augenhöhlen sowie eine kurze Schnauze mit einem großen, dreieckigen Antorbitalfenster. Auf der Schnauze saß eine Eizahn-artige Spitze, die vom Zwischenkieferbein (Prämaxillare) gebildet wurde und wahrscheinlich zum Aufbrechen der Eierschale gedient hat.[34] Wie Vergleiche mit den am besten erhaltenen Schädeln erwachsener Titanosauria zeigen, veränderten sich die Schädelknochen während des Wachstums drastisch: So verlängerte sich die Schnauze deutlich, während der Oberkieferknochen (Maxillare) eine Verbindung mit dem Quadratojugal entwickelte, die das Jochbein (Jugale) von der Unterseite des Schädels trennte. Die Nasenöffnungen wurden größer und wanderten bis über die Augenhöhlen. Die Augenhöhlen nahmen eine umgekehrt tränenartige Form an. Stirnbein (Frontale) und Scheitelbein (Parietale) verloren stark an Größe, was zu einem schmaleren Schädel führte.[30][35]
Osteoderme und Hautabdrücke
 Hautknochenplatte von Ampelosaurus atacis.
Hautknochenplatte von Ampelosaurus atacis.Im Jahr 1896 schrieb Charles Depéret eine große, zylindrische Platte dem Titanosaurier „Titanosaurus“ madagascariensis zu und war damit der erste, der eine Panzerung bei einem Sauropoden vermutete. Lange Zeit unbeachtet, konnte Depérets Hypothese erst 1980 mit der Entdeckung einer Panzerung bei Saltasaurus bestätigt werden. An einem Skelett dieser Gattung zeigen sich kleine, rundliche Knöchelchen, die im Kontakt miteinander stehen und eine mosaikartige Oberfläche bilden. Daneben waren größere, ovale Platten mit konischer Außenfläche vorhanden.[36] Seit diesem Fund wurden weitere Skelette mit assoziierten Osteodermen sowie isoliert vorgefundene, einzelne Osteoderme vor allem in Südamerika, aber auch in Afrika, Madagaskar und Europa entdeckt. Osteoderme sind für 10 der über 40 derzeit anerkannten Titanosaurier-Gattungen nachgewiesen. Wie zusammenhängende (artikulierte) Skelette verschiedener Gattungen ohne assoziierte Osteoderme zeigen, fehlte die Panzerung jedoch zumindest bei einigen Gattungen. Die bisher bekannten Osteoderme variieren in ihrer Größe von wenigen Millimetern bis einigen Dezimetern; der größte Fund zeigt einen Durchmesser von 59 Zentimetern. Die Titanosauria sind die einzigen Sauropoden, von denen gesicherte Funde von Osteodermen vorliegen, obwohl es Hinweise auf mögliche Osteoderme bei einigen Gattungen aus anderen Gruppen gibt, wie beispielsweise bei „Pelorosaurus“ becklesii.[37]
Funde von Titanosaurier-Osteodermen sind im Vergleich zu Knochenfunden sehr spärlich: So sind derzeit nur fast 90 Titanosaurier-Osteoderme bekannt, in Verbindung mit Skelettfunden sind es jeweils höchstens einige wenige, unabhängig vom Artikulationsgrad der Skelette. Im Gegensatz dazu sind Osteodermfunde anderer Gruppen, beispielsweise der Krokodile oder der thyreophoren Dinosaurier, sehr häufig. Dies deutet darauf hin, dass die Osteoderm tragenden Titanosaurier lediglich schwach gepanzert waren.[37] Leonardo Salgado (2003) bemerkt, dass die von ihm untersuchten Osteoderme entweder asymmetrisch oder zweiseitig symmetrisch sind, wobei die zweiseitig symmetrischen deutlich überwiegen. Im Gegensatz dazu sind die Osteoderme heutiger Alligatoren, die in Querstreifen über den Rücken verlaufen, nicht zweiseitig symmetrisch. Daraus schließt er, dass die symmetrischen Osteoderme in einer Reihe auf der Mittellinie des Körpers über den Wirbelstacheln verliefen, während die asymmetrischen Osteoderme auf den Flanken der Tiere verteilt waren.[38] Michael D'emic und Kollegen (2009) stellten fest, dass gepanzerte Titanosaurier generell kleiner waren als ungepanzerte – so zeigten gepanzerte Arten eine durchschnittliche Femurlänge von 87 Zentimetern, während die Femurlänge ungepanzerter Arten durchschnittlich 149 Zentimeter betrug. Die Evolutionsgeschichte der Osteoderme innerhalb der Titanosauria ist nicht bekannt.[37]
Verschiedene Autoren haben versucht, die bekannten Osteoderme anhand ihrer Form in Gruppen (Morphotypen) einzuteilen. Der jüngste Ansatz stammt von Michael D'emic und Kollegen (2009) und sieht vier Morphotypen vor: Der ellipsoide Typ (Morphotyp 1), die am häufigsten gefundene Morphe, die sich unter anderem bei Saltasaurus findet, ist durch oft große, ovale Platten mit konvexer Oberfläche charakterisiert und zeigt gelegentlich eine stachelartige Erhebung. Der gekielte Typ (Morphotyp 2) zeigt einen länglichen Kamm auf der Oberfläche. Der zylindrische Typ (Morphotyp 3) ist durch runde bis ovale, flache Platten gekennzeichnet, ist bisher aber lediglich von vier Exemplaren bekannt. Dem Mosaik-Typ (Morphotyp 4) werden die kleinen, mosaikartig angeordneten Plättchen zugeordnet, die beispielsweise bei Saltasaurus auftraten, sowie unregelmäßig geformte Osteoderme, die keinem anderen Typ zugeordnet werden können.[37]
Innerhalb einiger Eier von Auca Mahuevo fanden sich Hautabdrücke, die zeigen, dass die Hautoberfläche wie bei anderen Dinosauriern mit tuberkelartigen Schuppen bedeckt war, die sich nicht überlappten. Bei den Hautabdrücken der Embryonen von Auca Mahuevo fanden sich zudem längliche und rosettenförmige Strukturen, die durch eng beieinander liegende Schuppen gebildet wurden.[39] Einige Forscher vermuten, dass diese Strukturen in späteren Entwicklungsstadien verknöcherten und zu Osteodermen wurden. Diese Osteoderme könnten junge Titanosaurier vor kleinen Prädatoren geschützt haben. Für ausgewachsene Titanosaurier erscheint die Anzahl der Osteoderme jedoch als zu gering, um einen wirksamen Schutz vor Räubern geboten zu haben – es wird daher in Erwägung gezogen, dass die Osteoderme bei erwachsenen Titanosauriern als Calcium-Reservoire umfunktioniert wurden.[40]
Ursprung und Paläobiogeographie
Der älteste bekannte Titanosauria ist Janenschia aus dem Oberjura Tansanias, obwohl die Zuordnung dieser Gattung zu den Titanosauria nicht von allen Forschern anerkannt wird. Unzweifelhafte Titanosauria tauchen erstmals in Gesteinen der Unterkreide in Europa, Nord- und Südamerika, Afrika und Australien auf. Fährtenfolgen vom weitgängigen Typ, der von vielen Forschern Titanosauriern zugeschrieben wird, könnte jedoch darauf hinweisen, dass Titanosaurier bereits seit dem mittleren Jura existierten.[6]
Lydekker (1895) vermutete, dass Titanosaurier erst nach der Trennung der nördlichen (Laurasia) und südlichen Landmassen (Gondwana), die im Oberjura stattfand, auf Gondwana entstanden sind und in ihrer Verbreitung auf Gondwana beschränkt blieben. Zwar wurden in der Folgezeit einige Titanosaurier auch in den nördlichen Kontinenten gefunden – diese Funde wurden jedoch oft als Ausnahmen betrachtet, die von unabhängigen Vorstößen einiger Populationen von Süden nach Norden herrührten. Zahlreiche neuere Funde, die seitdem auch auf den Kontinenten des ehemaligen Laurasias gemacht werden, zeigen jedoch, dass Titanosaurier nicht auf Gondwana beschränkt waren. Heute gehen viele Forscher davon aus, dass sich Titanosaurier weltweit verbreiteten, noch bevor der Superkontinent Pangäa endgültig zerbrach, als noch Verbindungen zwischen den großen Landmassen bestanden.[6][41]
Forschungsgeschichte
 William Henry Sleeman fand 1828 zwei Wirbel, die später als Titanosaurus indicus beschrieben wurden.
William Henry Sleeman fand 1828 zwei Wirbel, die später als Titanosaurus indicus beschrieben wurden.Bereits 1828 entdeckte der britische Offizier William Henry Sleeman zwei Wirbel im zentralen Indien nahe Jabalpur. Diese Knochen gelangten 1832 ins Museum von Kalkutta, wo sie 1862 vom schottischen Paläontologen Hugh Falconer beschrieben wurden. Falconer erkannte, dass es sich um die Schwanzwirbel eines Reptils handelte und gab diagnostische Merkmale an, stellte jedoch keinen neuen Namen auf. Somit beschrieb erst Richard Lydekker 1877 auf Basis dieser Wirbel sowie eines fragmentarischen Oberschenkelknochens vom gleichen Fundort eine neue Art und Gattung, Titanosaurus indicus. Weitere Titanosaurus-Arten wurden einige Jahre später aus Madagaskar und Südamerika beschrieben. In der Folgezeit wurden Titanosaurus sowie die Titanosauridae zu sogenannten Papierkorb-Taxa, denen nahezu sämtliche Sauropodenfunde aus der Kreide zugeschrieben wurden, selbst wenn keine Gemeinsamkeiten mit anderen bekannten Titanosaurier-Überresten festgestellt werden konnten. Erst seit den 1970er-Jahren wurden weitere wichtige Funde insbesondere in Südamerika gemacht, wie beispielsweise Saltasaurus. Einige ursprüngliche Gattungen wie Andesaurus ähnelten zwar anderen Titanosauriern, ihnen fehlten jedoch wichtige gemeinsam abgeleitete Merkmale der abgeleiteteren Titanosaurier, wie beispielsweise procoele (auf der Vorderseite konkave) Schwanzwirbel. Um diese ursprünglichen Formen mit einzuschließen, wurde 1993 die Gruppe Titanosauria benannt. Mit Rapetosaurus wurde Anfang des 21. Jahrhunderts erstmals ein nahezu vollständiges Skelett eines Titanosauriers einschließlich Schädel entdeckt. Dieser Fund zeigte unter anderem, dass Nemegtosaurus und Quaesitosaurus, die durch isoliert gefundene, diplodociden-ähnliche Schädel bekannt sind, keine Diplodociden waren, sondern Titanosaurier. Bedeutend für das Verständnis der Evolution von Schädelmerkmalen und der Individualentwicklung war zudem die Entdeckung gut erhaltener embryonaler Schädel innerhalb von Eiern in Auca Mahuevo.[35] Neueste Funde schließen Diamantinasaurus[1] aus Australien sowie Atsinganosaurus[25] aus dem südlichen Frankreich mit ein, die in den Jahren 2009 bzw. 2010 beschrieben wurden.[6][41]
Systematik
Definition
Die Gruppe Titanosauria wurde 1993 von Bonaparte und Coria aufgestellt.[42] Diese Autoren listen zwar diejenigen Gattungen auf, die sie innerhalb der Titanosauria zusammenfassen wollen, geben jedoch keine exakte Definition dieser Gruppe an. Somit bleibt unklar, ob neu entdeckte, ursprüngliche Formen in diese Gruppe eingeschlossen werden sollten oder nicht. Spätere Autoren schlagen verschiedene Definitionen vor, die jedoch umstritten bleiben. Die erste Definition liefern Salgado und Kollegen (1997)[10]: Nach diesen Autoren sind die Titanosauria ein knotenbasiertes Taxon (node-based definition), das den letzten gemeinsamen Vorfahr von Andesaurus delgadoi und der Titanosauridae sowie alle Nachfahren dieses Vorfahren mit einschließt. Sereno und Wilson (1998) kritisieren diesen Vorschlag, da Andesaurus delgadoi wenig bekannt ist und andere Gattungen wie beispielsweise Chubutisaurus, die bisher als Vertreter der Titanosauria betrachtet wurden, möglicherweise basaler waren als Andesaurus. Diese Autoren stellen daher eine alternative, stammlinienbasierte Definition (stem-based definition) auf:[43] Nach dieser Definition schließen die Titanosauria alle Taxa mit ein, die näher mit Saltasaurus loricatus als mit Euhelopus zdanskyi und Brachiosaurus brancai verwandt sind. Des Weiteren benennen diese Autoren eine neue Gruppe, die Somphospondyli, die alle Taxa mit einschließen soll, die näher mit Saltasaurus loricatus als mit Brachiosaurus brancai verwandt sind. Nach diesen Definitionen schließt die Somphospondyli also, im Gegensatz zur Titanosauria, Euhelopus nicht aus. Die jüngste Definition stammt von Upchurch und Kollegen (2004).[4] Diese Autoren kritisieren die von Sereno und Wilson aufgestellte Definition der Titanosauria, da sie drei Referenztaxa enthält, was nach Ansicht dieser Autoren unpraktisch ist und zu Verwirrungen führt. Zudem basiere die Unterscheidung zwischen Somphospondyli und Titanosauria auf der systematischen Position von Euhelopus, die jedoch stark umstritten ist. Folglich synonymisieren diese Autoren Symphospondyli und Titanosauria und geben für die Titanosauria dieselbe Definition an, die von Sereno und Wilson für die Symphospondyli verwendet wurde: Ein stammlinienbasiertes Taxon, das alle Taxa mit einschließt, die näher mit Saltasaurus als mit Brachiosaurus verwandt sind.[44]
Äußere Systematik
Die Titanosauria bilden zusammen mit der Brachiosauridae und einigen ursprünglichen Formen die Gruppe Titanosauriformes. Einige Autoren sehen die Gattung Euhelopus als enge Verwandte der Titanosauria und fassen beide innerhalb der Gruppe Somphospondyli zusammen – diese Gruppe ist jedoch umstritten (siehe unten). Die Titanosauriformes wiederum bilden zusammen mit der Camarasauridae die Gruppe Macronaria. Auch auf dieser Ebene wird von einigen Autoren eine weitere Gruppe zwischengeschaltet, die Camarasauromorpha, die einige ursprüngliche Macronaria ausschließt.[45][4] Die Macronaria bilden zusammen mit den Diplodocoidea (Diplodocidae, Dicraeosauridae und Rebbachisauridae) die Gruppe Neosauropoda. Es folgt eine Beispielsystematik (vereinfacht nach Upchurch und Kollegen, 2004)[4].
Sauropoda Eusauropoda Neosauropoda Macronaria Titanosauriformes Titanosauria
Bis Ende der 90er Jahre wurden Titanosaurier als enge Verwandte der Diplodociden betrachtet. Eine nähere Verwandtschaft dieser beiden Gruppen wurde erstmals 1929 von Friedrich von Huene bei seiner Beschreibung des Schädels von Antarctosaurus vorgeschlagen: So zeigt dieser Schädel die für Diplodociden typischen schmalen Zahnkronen und die hoch am Schädel gelegenen Nasenöffnungen. Werner Janensch (1929) teilte die Sauropoden anhand ihrer Zahnmorphologie in zwei Gruppen ein – die schmalkronigen Arten (Titanosauridae und Diplodocidae) und die breitkronigen Arten (z. B. Brachiosauridae). Einige Autoren ordneten die Diplodocidae innerhalb der Titanosauridae ein (Romer, 1956, 1966, 1968), andere fassten beide Gruppen in einer Atlantosauridae genannten Gruppe zusammen (Steel, 1970). Erst die Arbeiten von Salgado und Kollegen (1997) und Wilson und Sereno (1998) zeigten, dass Titanosaurier nahe mit Brachiosaurus-ähnlichen Formen verwandt waren.[6][3]Innere Systematik
Die Innere Systematik der Titanosauria ist bis heute stark umstritten. So existiert eine Reihe von Vorschlägen, die sich in Aufbau und Benennung der Kladen häufig widersprechen. Erschwerend kommt hinzu, dass viele Analysen lediglich einige wenige Gattungen berücksichtigen, während viele andere, valide Gattungen bisher in keiner Analyse auftauchen.[4] Der Paläontologe Jeffrey Wilson schreibt: “The interrelationships of Titanosauria remain as one of the last frontiers in dinosaur systematics” („Die Verwandtschaftsbeziehungen der Titanosauria sind eine der letzten Fronten der Dinosaurier-Systematik“). Dennoch stimmen die Analysen in einigen Punkten miteinander überein: So wird Saltasaurus stets als die am meisten abgeleitete Gattung betrachtet, während Neuquensaurus meistens als Schwestertaxon von Saltasaurus behandelt wird. Beide Gattungen werden häufig als Saltasaurinae zusammengefasst – häufig zusammen mit Rocasaurus und manchmal zusätzlich mit weiteren Gattungen. Fast alle Analysen sehen Alamosaurus und Opisthocoelicaudia als nahe Verwandte der Saltasaurinae, manchmal werden beide Gattungen als Opisthocoelicaudiinae der Saltasaurinae gegenübergestellt. Die Opisthocoelicaudiinae und die Saltasaurinae werden häufig als Saltasauridae zusammengefasst. Andesaurus wird meistens als ursprünglichster Vertreter der Titanosauria betrachtet. Auch Malawisaurus wird von vielen Analysen als sehr ursprünglicher Titanosauria erachtet, obwohl einige Autoren (Upchurch, 1995 und Rogers, 2005) eine mehr abgeleitete Position dieser Gattung vorschlagen und vermuten, dass Malawisaurus, Saltasaurus und Neuquensaurus näher miteinander verwandt sind als mit Opisthocoelicaudia.[6]
Für Diskussionen sorgt die Frage, ob es monophyletische Gruppen ursprünglicher Titanosaurier gab. Bonaparte und Coria (1993) teilen die Titanosauria in zwei Familien, die ursprüngliche Andesauridae, bestehend aus Andesaurus und Argentinosaurus, und die abgeleitetere Titanosauridae. Upchurch (1998) behält die Andesauridae als ursprüngliche Titanosauriergruppe bei und ordnet ihr die Gattungen Andesaurus, Malawisaurus und Phuwiangosaurus zu. Dagegen halten Salgado und Kollegen (1997) und viele spätere Studien die Andesauridae für paraphyletisch und somit für ungültig. Calvo und Kollegen (2007) stellen mit den Lognkosauria eine neue Gruppe ursprünglicher Titanosaurier auf, die Mendozasaurus sowie den neuentdeckten Futalognkosaurus enthalten soll.[21] Des Weiteren werden Vorschläge diskutiert, die neben der Saltasauridae weitere Gruppen abgeleiteter Titanosaurier vorsehen. So fasst Wilson (2005) Quaesitosaurus und Nemegtosaurus zusammen mit Rapetosaurus innerhalb der Nemegtosauridae zusammen. Quaesitosaurus und Nemegtosaurus wurden häufig als Vertreter der Diplodocoidea betrachtet,[4] gelten in vielen neueren Analysen jedoch als Vertreter der Titanosauria.[8]
Uneinigkeit herrscht in Bezug auf die Benennung von Gruppen innerhalb der Titanosauria. So verwenden viele Analysen den Namen Titanosauridae, um abgeleitetere Titanosaurier unter Ausschluss basaler Formen wie Andesaurus zusammenzufassen. Wilson und Upchurch (2003) veröffentlichten eine Revision der Gattung Titanosaurus und deklarieren die Typusart Titanosaurus indicus als ungültig, da sie lediglich auf zwei Schwanzwirbeln basiert, die keine diagnostisch verwertbaren Merkmale zeigen. Konsequent betrachten diese Autoren daher rangbasierte Taxa, die auf Titanosaurus als nominotypisches Taxon basieren – die Titanosauridae, die Titanosaurinae sowie die Titanosauroidea – ebenfalls als ungültig. Titanosauria bleibt nach diesem Vorschlag als rangloses Taxon weiterhin gültig.[46][41] Upchurch und Kollegen (2004) fassen die stärker abgeleiteten Titanosauria in einer neuen, der Titanosauridae entsprechenden Gruppe zusammen, die Lithostrotia.[4] Während dieser Name von einigen neueren Analysen übernommen wird,[3] verwenden Calvo und Kollegen (2007) für dieselbe Gruppe weiterhin den Namen Titanosauridae. Einige Autoren benutzen des Weiteren den Namen Eutitanosauria für abgeleitete Titanosauria.[21]
Dieses Kladogramm folgt Calvo und Kollegen (2007):[21]
Titanosauria Titanosauridae (=Lithostrotia) Lognkosauria Eutitanosauria Aeolosaurini Loma Lindero sp.
Lirainosaurus
Opisthocoelicaudiinae Saltasaurinae Rocasaurus
Gattungsliste der Titanosauria (ergänzt nach Upchurch und Kollegen, 2004)[4]Literatur
- Upchurch, Barrett, Dodson: Sauropoda. In: Weishampel, Dodson, Osmólska (Hrsg.): The Dinosauria. 2te Auflage. University of California Press, 2004, ISBN 0-520-24209-2, S. 259–322.
- Jeffrey A. Wilson: An Overview of Titanosaur Evolution and Phylogeny. In: III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos 2006, S. 169–190.
- Kristina Curry Rogers: Titanosauria. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- Leonardo Salgado, Rodolfo Anibal Coria, Jorge Orlando Calvo: Evolution of Titanosaurid Sauropods. I: Phylogenetic Analysis Based on the Postcranial Evidence. In: Ameghiniana. 34, Nr. 1, 1997, ISSN 0002-7014, S. 3–32.
- Luis M. Chiappe, Frankie Jackson, Rodolfo A. Coria, Lowell Dingus: Nesting Titanosaurs from Auca Mahuevo and Adjacent Sites. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- Jeffrey A. Wilson, Paul Upchurch: A Revision of Titanosaurus Lydekker (Dinosauria - Sauropoda), the first dinosaur genus with a 'gondwanan' distribution. In: Journal of Systematic Palaentology. 1, Nr. 3, 2003, S. 125–160.
Einzelnachweise
- ↑ a b c d e Scott A. Hocknull, Matt A. White, Travis R. Tischler, Alex G. Cook, Naomi D. Calleja, Trish Sloan, David A. Elliott: New mid-Cretaceous (latest Albian) dinosaurs from Winton, Queensland, Australia. In: PLoS ONE. 4, Nr. 7, 2009, doi:10.1371/journal.pone.0006190.
- ↑ a b c d e J. A. Wilson, D. M. Mohabey, S. E. Peters, J. J. Head: Predation upon Hatchling Dinosaurs by a New Snake from the Late Cretaceous of India. In: PLoS Biol. 8, Nr. 3 e1000322, 2010, doi:10.1371/journal.pbio.1000322.
- ↑ a b c d Kristina Curry Rogers: Titanosauria. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- ↑ a b c d e f g h i j k l m Upchurch, Barrett, Dodson: Sauropoda. In: Weishampel, Dodson, Osmólska (Hrsg.): The Dinosauria. 2te Auflage. University of California Press, 2004, ISBN 0-520-24209-2, S. 259–322.
- ↑ a b c Hussam Zaher, Diego Pol, Alberto B. Carvalho, Paulo M. Nascimento, Claudio Riccomini, Peter Larson, Rubén Juarez-Valieri, Ricardo Pires-Domingues, Nelson Jorge da Silva Jr., Diógenes de Almeida Campos: A Complete Skull of an Early Cretaceous Sauropod and the Evolution of Advanced Titanosaurians. In: PLoS ONE. 6, Nr. 2, 2011, S. e16663, doi:10.1371/journal.pone.0016663 (http://www.plosone.org/article/browse.action?field=&pageSize=10&startPage=0&catName=Evolutionary+Biology).
- ↑ a b c d e f g h i j k Jeffrey A. Wilson: An Overview of Titanosaur Evolution and Phylogeny. In: III Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno. Salas de los Infantes, Burgos 2006, S. 169–190.
- ↑ Kristina Curry Rogers, Catherine A. Forster: The Skull of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. In: Journal of Vertebrate Paleontology. 24, Nr. 1, 2004, S. 121–144.
- ↑ a b c d e f Jeffrey A. Wilson: Redescription of the Mongolian Sauropod Nemegtosaurus mongoliensis Nowinski (Dinosauria: Saurischia) and comments on Late Cretaceous Sauropod diversity. In: Journal of Systematic Palaeontology. 3, Nr. 3, 2005, S. 283–318, doi:10.1017/S1477201905001628.
- ↑ a b c Sebastián Apesteguía: Evolution of the Hyposphene-Hypantrum Complex within Sauropoda. In: Thunder-Lizards: The Sauropodomorph Dinosaurs, herausgegeben von Virginia Tidwell und Kenneth Carpenter, 2005, Indiana University Press, ISBN 0-253-34542-1
- ↑ a b c d Leonardo Salgado, Rodolfo Anibal Coria, Jorge Orlando Calvo: Evolution of Titanosaurid Sauropods. I: Phylogenetic Analysis Based on the Postcranial Evidence. In: Ameghiniana. 34, Nr. 1, 1997, ISSN 0002-7014, S. 3–32.
- ↑ a b c d Jeffrey Wilson: Overview of Sauropod Phylogeny and Evolution. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- ↑ Mathew Wedel: Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. In: Paleobiology. 29, Nr. 2, 2003, S. 243–255 (PDF).
- ↑ a b Jeffrey A. Wilson, Matthew T. Carrano: Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. In: Paleobiology. 52, Nr. 2, 1999, S. 252–267.
- ↑ Matthew T. Carrano: The Evolution of Sauropod Locomotion – morphological diversity of a secondarily quadrupedal radiation. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- ↑ Sebastián Apesteguía: Evolution of the Titanosaur Metacarpus. In: Thunder-Lizards: The Sauropodomorph Dinosaurs, herausgegeben von Virginia Tidwell und Kenneth Carpenter, 2005, Indiana University Press, ISBN 0-253-34542-1
- ↑ Alejandro Otero: The appendicular skeleton of Neuquensaurus, a Late Cretaceous saltasaurine sauropod from Patagonia, Argentinia. In: Acta Palaeontologica Polonica. 55, Nr. 3, 2010, S. 399–426.
- ↑ Bernardo J. González Riga, Jorge Orlando Calvo, Juan Porfiri: An articulated titanosaur from Patagonia (Argentina): New evidence of neosauropod pedal evolution. In: Palaeoworld. 17, 2008, S. 33–40.
- ↑ a b Kenneth Carpenter: Biggest of the Big: A Critical Re-Evaluation of the Mega-Sauropod Amphicoelias fragillimus Cope, 1878. In: John R. Foster, Spencer G. Lucas (Hrsg.): Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 2006, S. 131–138.
- ↑ a b Gerardo V. Mazzetta, Per Christiansen, Richard A. Fariña: Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs. In: Historical Biology. 65, 2004, S. 1–13, doi:10.1080/08912960410001715132 (PDF).
- ↑ G. P. Burness, T. Flannery: Dinosaurs, dragons, and dwarfs: The evolution of maximal body size. In: Proceedings of the National Academy of Sciences. 98, Nr. 25, 2001, S. 14518–14523.
- ↑ a b c d e Jorge O. Calvo, Juan D. Porfiri, Bernardo J. Gonzáles-Riga, Alexander W. A. Kellner: A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur. In: Anais da Academia Brasileira de Ciências. 79, Nr. 3, 2007, ISSN 0001-3765, S. 529–541.
- ↑ a b c P. Martin Sander, Andreas Christian, Marcus Clauss, Regina Fechner, Carole T. Gee, Eva-Maria Griebeler, Hanns-Christian Gunga, Jürgen Hummel, Heinrich Mallison, Steven F. Perry, Holger Preuschoft, Oliver W. M. Rauhut, Kristian Remes, Thomas Tütken, Oliver Wings, Ulrich Witzel: Biology of the sauropod dinosaurs: the evolution of gigantism. In: Biological Reviews. Cambridge Philosophical Society, 2010, doi:10.1111/j.1469-185X.2010.00137.x (PDF).
- ↑ Jeffrey Wilson, Kristi Curry Rogers: Body Size Evolution. In: Evolution & Phylogeny of Titanosauria. Abgerufen am 23. September 2010.
- ↑ Oliver Wings, Martin Sander: No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches. In: The Royal Society. 274, 2006, S. 635–640 (PDF).
- ↑ a b c G. Garcia, S. Amico, F. Fournier, E. Thouand, X. Valentin: A new Titanosaur genus (Dinosauria, Sauropoda) from the Late Cretaceous of southern France and its paleobiogeographic implications. In: Bulletin de la Societe Geologique de France. 181, Nr. 3, 2010, S. 269–277.
- ↑ Paul M. Barrett, Paul Upchurch: Sauropodomorph Diversity through Time: Paleoecological and macroevolutionary implications. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- ↑ Vandana Prasad, Caroline Stromberg, Habib Alimohammadian, Ashok Sahni: Dinosaur Coprolites and the Early Evolution of Grasses and Grazers. In: Science. 310, 2005, S. 1177–1180.
- ↑ Jeffrey A. Wilson, Ricardo N. Martinez, Oscar Alcober: Distal tail segment of a titanosaur (Dinosauria: Sauropoda) form the Upper Cretaceous of Mendoza, Argentinia. In: Journal of Vertebrate Paleontology. 19, Nr. 3, 1999, S. 591–594.
- ↑ a b c d e Bernat Vila, Frankie D. Jackson, Josep Fortuny, Albert G. Selles, Angel Galobart: 3-D Modelling of Megaloolithid Clutches: Insights about Nest Construction and Dinosaur Behaviour. In: PLoS ONE. 5, Nr. 5, e10362, 2010, doi:10.1371/journal.pone.0010362.
- ↑ a b c d e f Luis M. Chiappe, Frankie Jackson, Rodolfo A. Coria, Lowell Dingus: Nesting Titanosaurs from Auca Mahuevo and Adjacent Sites. In: Rogers, Wilson (Hrsg.): The Sauropods: Evolution and Paleobiology. University of California Press, 2005, ISBN 0-520-24623-3.
- ↑ a b Gerald Grellet-Tinner, Luis M. Chiappe, R. Coria: Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentinia). In: Can. J. Earth Sci.. 41, 2004, S. 949–960, doi:10.1139/E04-049.
- ↑ Frankie D. Jackson, David J. Varricchio, Robert A. Jackson, Bernat Vila, Luis M. Chiappe: Comparison of water vapor conductance in a titanosaur egg from the Upper Cretaceous of Argentina and a Megaloolithus siruguei egg from Spain. In: Paleobiology. 34, Nr. 2, 2008, S. 229–246.
- ↑ Luis M. Chiappe, James G. Schmitt, Frankie D. Jackson, Alberto Garrido, Lowell Dingus, Gerald Grellet-Tinner: Nest Structure for Sauropods: Sedimentary Criteria for Recognition of Dinosaur Nesting Traces. In: Palaios. 19, 2004, S. 89–95.
- ↑ Rodolfo A. Gracía: An “egg-tooth”-like structure in titanosaurian sauropod embryos. In: Journal of Vertebrate Paleontology. 27, Nr. 1, 2007, S. 247–252.
- ↑ a b Luis M. Chiappe, Leonardo Salgado, Rodolfo A. Coria: Embryonic Skulls of Titanosaur Sauropod Dinosaurs. In: Science. 293, Nr. 2444, 2001, doi:10.1126/science.1063723.
- ↑ Ignacio A. Cerda, Jaime E. Powell: Dermal armor histology of Saltasaurus loricatus, an Upper Cretaceous sauropod dinosaur from Northwest Argentinia. In: Acta Palaeontologica Polonica. 55, Nr. 3, 2010, S. 389–398.
- ↑ a b c d M. D. d’Emic, J. A. Wilson, S. Chatterjee: The titanosaur (Dinosauria: Sauropoda) osteoderm record: review and first definitive specimen from India. In: Journal of Vertebrate Paleontology. 29, 2009, S. 165–177.
- ↑ Leonardo Salgado: Considerations on the bony plates assigned to titanosaurs (Dinosauria, Sauropoda). In: Ameghiniana. 40, Buenos Aires, ISSN 0002-7014, S. 441–456 (Abstract).
- ↑ Rodolfo A. Coria, Luis M. Chiappe: Embryonic skin from late cretaceous sauropods (Dinosauria) of Auca Mahuevo, Patagonia, Argentinia. In: J. Paleont.. 81, Nr. 6, 2007, S. 1528–1532.
- ↑ Thiago da Silva Marinho: Functional aspects of titanosaur osteoderms. In: Nature Precedings. 2007.
- ↑ a b c d e Jeffrey A. Wilson, Paul Upchurch: A Revision of Titanosaurus Lydekker (Dinosauria - Sauropoda), the first dinosaur genus with a 'gondwanan' distribution. In: Journal of Systematic Palaentology. 1, Nr. 3, 2003, S. 125–160.
- ↑ J. F. Bonaparte, R. A. Coria: Un nuevo y gigantesco saurópodo titanosaurio de la Formación Río Limay (Albiano–Cenomanio) de la Provincia del Neuquén, Argentinia. In: Ameghiniana. 30, 1993, S. 271–282.
- ↑ Jeffrey A. Wilson, Paul C. Sereno: Early Evolution and Higher-level Phylogeny of Sauropod Dinosaurs. In: Journal of Vertebrate Paleontology. 18, Ergänzung zu Nummer 2, 1998.
- ↑ Paul Sereno: Titanosauria. In: Taxon Search. Abgerufen am 18. September 2010.
- ↑ Paul Sereno: Camarasauromorpha. In: Taxon Search. Abgerufen am 28. September 2010.
- ↑ Paul Sereno: Titanosauridae. In: Taxon Search. Abgerufen am 20. September 2010.
- ↑ R. A. Santucci, R. J. Bertini: A new titanosaur from western São Paolo State, Upper Cretaceous Bauru Group, south-east Brazil. In: Palaeontology. 49, Nr. 1, 2006, S. 171–185.
- ↑ Sebastián Apesteguía: The sauropod diversity of the La Amarga Formation (Barremian), Neuquén (Argentina). In: Gondwana Research. 12, Nr. 4, 2007, S. 533–546, doi:10.1016/j.gr.2007.04.007.
- ↑ Leonardo Salgado, Rodolfo A. Coria, 2009: Barrosasaurus casamiquelai gen. et sp. nov., a new titanosaur (Dinosauria, Sauropoda) from the Anacleto Formation (Late Cretaceous: early Campanian) of Sierra Barrosa (Neuquén, Argentina). In: Zootaxa 2222, Seiten 1–16. Abstract
- ↑ A. W. A. Kellner, D. d. A. Campos, M. N. F. Trotta: Description of a titanosaurid caudal series from the Bauru Group, Late Cretaceous of Brazil. In: Arquivos do Museu Nacional, Rio de Janeiro. 63, Nr. 3, 2005, S. 529–564.
- ↑ Sebastián Apesteguía: Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia. In: Naturwissenschaften. 91, 2004, S. 493–497, doi:10.1007/s00114-004-0560-6.
- ↑ A. Martinelli, A. M. Forasiepi: Late Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Río Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae). In: Revista del Museo Argentino de Ciencias Naturales, nuevo serie. 6, Nr. 2, 2004, S. 257–305.
- ↑ H. You, Q. Ji, M. C. Lamanna, J. Li, Y. Li: A titanosaurian sauropod dinosaur with opsithocoelous caudal vertebrae from the early Late Cretaceous of Liaoning province, China. In: Acta Geologica Sinica. 78, Nr. 4, 2004, S. 907–911.
- ↑ H. You, F. Tang, Z. Luo: A new basal titanosaur (Dinosauria: Sauropoda) from the Early Cretaceous of China. In: Acta Geologica Sinica. 77, Nr. 4, 2003, S. 424–429.
- ↑ L. Salgado, R. A. Coria: Sauropods of Patagonia: systematic update and notes on global sauropod evolution. In: Thunder-Lizards: The Sauropodomorph Dinosaurs, herausgegeben von Virginia Tidwell und Kenneth Carpenter, 2005, Indiana University Press, ISBN 0-253-34542-1
- ↑ E. M. Gomani: Sauropod dinosaurs from the Early Cretaceous of Malawi. In: Palaeontologia Electronica. 8, Nr. 1, 2005, S. 1–37.
- ↑ A. W. A. Kellner, D. A. Campos, S. A. K. Azevedo, M. N. F. Trotta , D. D. R. Henriques, M. M. T. Craik, H. P. Silva: On a new titanosaur sauropod from the Bauru Group, Late Cretaceous of Brazil. In: Boletim do Museu Nacional (Geologia). 74, 2006, S. 1–31 (PDF).
- ↑ a b B. J. González Riga, E. Previtera, C. A. Pirrone: Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. In: Cretaceous Research. 30, Nr. 1, 2009, S. 135–148.
- ↑ J. O. Calvo, B. J. González-Riga, J. D. Porfiri: Muyelensaurus pecheni gen. et sp. nov., a new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina. In: Arquivos do Museu Nacional. 65, Nr. 4, Rio de Janeiro 2007, S. 485–504 (PDF).
- ↑ J. O. Calvo, J. D. Porfiri, 2010: Panamericansaurus schroederi gen. nov. sp. nov. Un nuevo Sauropoda (Titanosauridae-Aeolosaurini) de la Provincia del Neuquén, Cretácico Superior de Patagonia, Argentina. In: Brazilian Geographical Journal: Geosciences and Humanities Research Medium 1, Seiten 100–115.
- ↑ Fernando E. Novas, Leonardo Salgado, Jorge Calvo, Federico Agnolin: Giant titanosaur (Dinosauria, Sauropoda)from the Late Cretaceous of Patagonia. In: Revisto del Museo Argentino de Ciencias Naturales, n. s.. 7, Nr. 1, 2005, S. 37–41 (PDF).
- ↑ Jorge Coria, B. J. G. Riga: Rinconsaurus caudamirus gen. et sp nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina. In: Revista Geologica de Chile. 30, Nr. 2, 2003, S. 333–353 (http://www.scielo.cl/scielo.php?script=sci_arttext&pid=S0716-02082003000200011&lng=es&nrm=iso&tlng=en).
- ↑ X. Xu, X. Zhang, Q. Tan, X. Zhao, L. Tan: A new titanosaurian sauropod from Late Cretaceous of Nei Mongol, China. In: Acta Geologica Sinica. 80, Nr. 1, 2006, S. 20–26.
- ↑ R. D. Juárez Valieri, J. O. Calvo, 2011: Revision of MUCPv 204, a Senonian basal titanosaur from northern Patagonia. In: J. O. Calvo, J. D. Porfiri, B. J. González Riga, D. Dos Santos (herg.): Dinosaurios y Paleontología desde América Latina. Anales del III Congreso Latinoamericano de Paleontología. Editorial de la Universidad Nacional de Cuyo, Neuquén 143–152. PDF
- ↑ D. d. A. Campos, A. W. A. Kellner, R. J. Bertini, R. M. Santucci, 2005: On a titanosaurid (Dinosauria, Sauropoda) vertebral column from the Bauru Group, Late Cretaceous of Brazil. In: Arquivos do Museu Nacional, Rio de Janeiro, Band 63, Nummer 3, Seiten 565–593
- ↑ L. Salgado, I. d. S. Carvalho, 2008: Uberabatitan ribeiroi, a new titanosaur from the Marília Formation (Bauru Group, Upper Cretaceous), Minas Gerais, Brazil. In: Palaeontology 51, 4. Seiten 881–901.
- ↑ José F. Bonaparte, Bernardo J. González Riga, Sebastián Apesteguía: Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. In: Cretaceous Research. 27, Nr. 3, 2006, S. 364–376.
Weblinks
 Commons: Titanosauria – Sammlung von Bildern, Videos und Audiodateien
Commons: Titanosauria – Sammlung von Bildern, Videos und Audiodateien- Evolution & Phylogeny of Titanosauria Museum of Paleontology – The University of Michigan


Dieser Artikel wurde am 12. November 2010 in dieser Version in die Liste der exzellenten Artikel aufgenommen. Kategorien:- Titanosauria
- Titanosaurier
- Fauna des Juras
- Fauna der Kreide
- Wikipedia:Exzellent







Wikimedia Foundation.