- Aiphanes
-
Stachelpalmen .jpg)
Aiphanes horrida
Systematik Unterklasse: Commelinaähnliche (Commelinidae) Ordnung: Palmenartige (Arecales) Familie: Palmengewächse (Arecaceae) Tribus: Bactridinae Untertribus: Cocoeae Gattung: Stachelpalmen Wissenschaftlicher Name Aiphanes Willd. Die Stachelpalmen (Aiphanes), auch Stilettpalmen, sind eine Gattung von 25 Arten der Palmengewächse (Arecaceae), die auf den Kleinen Antillen, sowie von Venezuela bis Bolivien entlang der Anden verbreitet sind. Bis auf eine Art, die noch Panama erreicht, fehlen die Stachelpalmen in Mittelamerika. Sie fallen vor allem durch ihre bis über 25 Zentimeter langen Stacheln auf, die die Pflanzen fast komplett bedecken.
Inhaltsverzeichnis
Beschreibung
Stachelpalmen sind ausdauernde, verholzende Pflanzen. Durch unterschiedliche Verzweigungsarten können die Palmen verschiedene Erscheinungsformen (Habitus) besitzen. So gibt es beispielsweise von A. erinaceae solitär stehende Einzelbäume, aber auch strauchartige Wuchsformen mit über 20 Stämmen, die auf offenen Ebenen stehen.
Charakteristisch sind die etwa einen Millimeter bis über 25 Zentimeter langen Stacheln, die fast die gesamte Pflanze bedecken. Bewehrt sind Stamm, Blätter sowie Blütenstiele und Tragblätter der Blütenstände. Die Stacheln sind grau bis schwarz, bei A. erinacea, A. simplex und A. tricuspidata aber gelb. Die Stacheln wachsen aus einer Gruppe runder, dickwandiger Zellen aus, die zunächst eine Verdickung (Pulvinus) bilden. Kleine Stacheln sind Einzelzellen, mit stark verdickter (sklerenchymatisierter) Zellwand. Große Stacheln bestehen aus äußeren Sklerenchymzellen und innerem Parenchym.
Die Chromosomenzahl ist umstritten, in der Literatur finden sich widersprüchliche Angaben, wie zum Beispiel für A. minima n = 15 oder n = 18, und für A. horrida n = 16 oder n = 15. Wahrscheinlich handelt es sich bei den Unterschieden innerhalb einer Art aber um Artefakte. Wahrscheinlich, aber nicht überprüft, ist n = 15 für die ganze Gattung. Für die nahe verwandten Acrocomia, Gastrococcus und Astrocaryum gilt in jedem Fall n = 15.
Wurzeln
Fast alle Arten bilden oberirdische Adventivwurzeln aus, die manchmal dem unteren Teil des Stammes Kegelform verleihen. Sie sind zugleich auch Stützwurzeln und gleichen damit das fehlende sekundäre Dickenwachstum der Sprossachse aus. Die Adventivwurzeln sind gräulich bis rötlich braun und oft verzweigt. Sie erreichen einen Durchmesser zwischen fünf und 15 Millimetern. Sie sind mit weißlichen, warzenartigen Lentizellen übersät, die endogen entstehen und werden Pneumatophore oder Atemwurzeln genannt. Sie dienen dem Gasaustausch auf sehr feuchtem Boden.
Die Exodermis ist deutlich ausgeprägt. Die Rinde besteht hauptsächlich aus Parenchym mit unregelmäßigen luftgefüllten Interzellularräumen. Die Endodermis ist verholzt. Die Stele ist von Mark umgeben. Bislang konnten bei A. macroloba, A. ulei und A. weberbaueri Arbuskuläre Mykorrhizapilze in der Wurzelrinde nachgewiesen werden.
Sprossachse
.jpg) Stark bewehrte Ansatzstelle der Blätter an der Sprossachse
Stark bewehrte Ansatzstelle der Blätter an der Sprossachse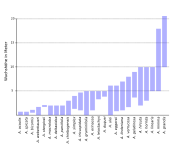 Verteilung der Wuchshöhe bei den Stachelpalmenarten
Verteilung der Wuchshöhe bei den StachelpalmenartenDie Sprossachse ist verzweigt oder unverzweigt. Sie kann sehr kurz sein und bei A. acaulis und A. spicata komplett unterirdisch liegen. Bei A. grandis wird sie bis über 20 Meter hoch und erreicht einen Durchmesser bis zu 20 Zentimetern.
Der Stamm ist stets deutlich ringförmig durch Blattnarben gezeichnet und mit Stacheln bewehrt, die ring- oder spiralförmig um die Nodi stehen. Die Länge der Internodien variiert und spiegelt unterschiedliche Wachstumsgeschwindigkeiten wider. Bei den kleinen Arten wie A. chiribogensis sind sie üblicherweise zwischen ein und zwei Zentimetern, bei den großen Arten wie A. eggersii bis zu 15 Zentimeter lang.
Verzweigungen entstehen aus den basalen (unterirdischen oder knapp über der Erdoberfläche liegenden) Blattachseln, oder selten distal direkt aus der Sprossachse.
Der Querschnitt entspricht dem charakteristischen Bau der Einkeimblättrigen: außen liegt die Epidermis, die eine Schicht primärer Rinde umgibt. Diese Schicht ist üblicherweise sehr dünn oder kaum vorhanden, bei A. macroloba aber sehr ausgeprägt und etwa 5 Millimeter dick. Weiter im Inneren liegt ein Zylinder aus Parenchym, in dem verteilt die Leitbündel verlaufen (Ataktostele). Der äußere Teil des Zylinders besteht aus schwarzen Sklerenchymfasern.
Blätter
.jpg) Wedel von Aiphanes horrida, gut erkennbar ist die fadenartige distale Verlängerung der Rhachis.
Wedel von Aiphanes horrida, gut erkennbar ist die fadenartige distale Verlängerung der Rhachis..jpg) Details: gut erkennbar sind die Stacheln aus der Mittelrippe der Blattunterseite.
Details: gut erkennbar sind die Stacheln aus der Mittelrippe der Blattunterseite.Fast alle Arten haben eine schraubige (disperse) Blattstellung (Phyllotaxis), nur bei A. linearis, A. verrucosa und A. lindeniana stehen die Blätter zweizeilig (distich). An Jungpflanzen stehen bei allen Arten die Blätter zweizeilig, was sich aber schnell verliert. Die Laubblätter sind große Fiedern (Palmwedel), deren Anzahl pro Krone von drei bis mehr als 20 variiert.
Die Jungblätter stecken in zylindrischen Blatthüllen, die bei allen Arten dicht mit Stacheln übersät sind. Die Blatthülle bricht dann an beiden Seiten auf und gibt das gestielte Blatt und ein deutlich sichtbares Blatthäutchen (Ligula) frei. Dieses bildet sich dann zurück, Reste bleiben aber auch an alten Blättern sichtbar. Die Blatthülle fällt bei allen Arten mit Ausnahme von A. hirsuta subsp. fosteriorum sauber ab. Die Jungblätter sind dicht mit vielverzweigten Blatthaaren (Trichomen) bedeckt.
Die Länge der Blattstiele variiert zwischen wenigen Zentimetern bis zu über einem Meter. Kurze Blattstiele sind oben eingekerbt, wohingegen lange rundlich ohne Kerbe sind. Die mittlere Hauptachse des Fiederblatts (Rhachis) verlängert sich distal in einen langen Faden (Filament), der dann mit der Zeit abbricht. Blattstiele und Rhachis sind oft dicht mit Stacheln übersät, bei wenigen Arten aber komplett unbewehrt.
Die Anordnung der einzelnen Fiederblättchen an der Blattspindel variiert zwischen den Arten stark. Sie reicht von komplett unregelmäßig über in Haufen gruppiert hin zu paarig gegenüberstehenden Blättchen.
Während des Wachstums wird das Blättchen zweimal von der Spitze abwärts (basipetal) eingeschnitten, und bildet drei ungleichmäßige Lappen. Die Mittelrippe der Blattunterseite ist bei allen Arten mit einem oder mehreren Stacheln bewehrt. Bei einigen Arten, wie zum Beispiel A. ulei, ist aber auch die Blattoberseite mit Reihen von Stacheln besetzt. Bei A. minima können die Stacheln auch aus den Seitenrippen auswachsen.
Blattanatomie
Die Blattspreite ist dorsiventral. Die Epidermis ist nur eine Zelllage stark. Die Zellen sind rhomboedrisch oder spindelförmig. Die außen liegende Zellwand ist schwach bis stark cutinisiert. Die antiklinen, das heißt senkrecht zur Blattoberfläche stehenden, Zellwände sind periodisch verdickt. Die Verdickungen erinnern an auf eine Schnur aufgereihte Perlen. Die Spaltöffnungen (Stomata) liegen bevorzugt abaxial, das heißt an der Blattunterseite. Selten sind diese eingesunken oder herausgehoben.
Die Hypodermis, das heißt die Schicht direkt unter der Epidermis, ist ebenfalls nur eine Zelle stark. Die Zellen sind doppelt so breit wie die der Epidermis. Die Atemhöhlen unter den Spaltöffnungen sind von neun Zellen umgeben. Das Chlorenchym ist einheitlich und ein bis drei Zellen dick. Rhaphiden, Bündel aus nadelförmigen Calciumoxalat-Kristallen, sind häufig. Fasern, die nicht mit den Gefäßbündeln assoziiert sind, sind kleiner als fünf Mikrometer im Durchmesser dick und ohne Sklerenchym. Sie liegen in Strängen aus zwei bis vier Schichten.
Die Blattnerven verlaufen im Mesophyll, sie bestehen aus einer äußeren Parenchymschicht, die vor allem größere Nerven oft nicht vollständig umschließt, und einer inneren ein bis sieben Zellen starken Schicht aus Sklerenchym. Das Phloem der Hauptnerven besteht aus zwei bis vier Strängen.
Blütenstände
Stachelpalmen blühen mehrmals im Leben, sie sind also ausdauernd (pollakanth). Die Blüten sind getrenntgeschlechtig einhäusig (monözisch).
Die Blütenstände stehen zunächst aufrecht und können während der Anthese, wenn sich die Blüten öffnen, kippen, bis sie herabhängen. Während der Fruchtreife werden sie häufig gebogen. Bei fast allen Arten entwickelt sich ein Blütenstand pro Blattachse, bei A. gelatinosa sind es jedoch häufig drei. Die Tragblätter (Brakteen) sind sehr variabel, sie können verdickt und holzig oder dünn und papierartig sein. Sie sind häufig mit Stacheln bewehrt, dünne Tragblätter fallen häufig ab, während holzige erhalten bleiben.
Der Blütenstand ist bei fast allen Arten kolbenartig, dabei aber einfach verzweigt. Die Blütenstandsachse besteht aus einem bewehrten Blütenstandsstiel (Pedunculus) mit einem Durchmesser von drei bis 50 Millimeter und einer Rhachis, von der Achsen erster Ordnung (Rhachillae) abzweigen, an denen dann die Blüten sitzen. Bei A. acaulis, A. spicata und A. macroloba, ist der Blütenstand jedoch immer, bei A. simplex meistens, unverzweigt. Am Blütenstiel sitzen flache, abgerundete und an den Seiten geflügelte Vorblätter (Brakteolen).
Die Blüten sitzen immer in Dreiergruppen aus zwei männlichen Blüten (mit Androeceum) und einer weiblichen Blüte (mit Gynoeceum). Selten ergeben sich an der Spitze der Blütenstände (distal) Zweiergruppen, bei denen die weibliche Blüte fehlt. Bei A. deltoidea und A. minima ergeben sich selten Vierergruppen, aus jeweils zwei Geschlechterpaaren.
Männliche Blüten
Die männlichen Blüten sind gestielt oder fast aufsitzend. Die Blütenhülle besteht aus drei freien, gekielten und häutchenartigen Kelchblättern (Sepalen) und drei freien oder an der Basis verwachsenen, fleischigen Kronblättern (Petalen). Beide sind spitz. Die Blütenfarbe variiert von cremefarben zu gelb-orange, oder von weiß ins purpurn oder violett. Häufig sind die Blüten vor der Anthese noch grün.
Die Kelchblätter bestehen nur aus ein bis zwei Schichten schmaler Zellen. Die Kronblätter sind deutlich dicker, neben der Epidermis bestehen sie aus fünf oder sechs Schichten von Parenchymzellen, mit vielen Rhaphiden und tanninhaltigen Zellen, und einer Schicht aus Fasern, die nicht mit den Gefäßbündeln assoziiert sind. Sie werden durch ein einzelnes Leitbündel mit zwei Tracheen aus Metaxylem versorgt. Die innere Epidermis besteht aus einer Schicht großer Zellen mit einem zähen Zytoplasma und großen Zellkernen.
Die sechs Staubblätter stehen in zwei Quirlen zu je drei. Sie werden durch ein einzelnes Leitbündel mit drei bis vier Tracheen aus Metaxylem versorgt. Die Staubfäden sind an der Basis verwachsen. Sie stehen aufrecht und sind nie länger als die Kronblätter. Im Durchmesser haben sie zehn bis 15 Zellen und bestehen aus Parenchym. Die Antheren öffnen sich zum Zentrum der Blüte hin (intrors) oder zur Seite hin (latrors). Sie sind abgerundet und gebogen, oder aufrecht – bei der Anthese neigen sie sich oft bis fast in die Horizontale. Die Länge variiert zwischen 0,3 und vier Millimetern und korreliert mit der Größe der Kronblätter. Die Pollensäcke sind reich an Raphiden. Es existieren kleine, drüsige Stempelrudimente, die zu Nektarien umgebildet sind.
Pollen
Die Pollenkörner sind monosulcat, das heißt, sie haben nur eine Keimfurche. Diese liegt häufig in der Südhälfte des Pollenkorns (meridionosulcat). Selten finden sich dreiarmige Keimfurchen (trichotomosulcat). Sie sind kugelförmig bis ellipsoid, selten dreieckig. Die Längsachse ist zwischen 20 und 30 Mikrometer lang. Der Durchmesser variiert zwischen 20 und 30 Mikrometer.
Die äußere Schicht der Pollenkörner (Exine) ist ganz oder zum Teil mit einem Tectum bedeckt, einer Schicht, die die Columellae genannten stäbchenförmigen Strukturen bedeckt. Auf dem Tectum sitzen häufig kurze oder lange Dornen, Warzen oder mehr oder weniger stark verwachsene zarte Auswüchse. Die Exinestruktur und Ornamentierung ist insgesamt sehr viel diverser als bei anderen Bactridinae-Gattungen.[1]
Weibliche Blüten
Die Blütenhülle der weiblichen Blüten besteht aus drei freien, breit gekielten, dünnen, papierähnlichen Kelchblättern und drei gebogenen oder nur wenig gekielten, fleischigen Kronblättern. Letztere sind auf halber Länge basal verwachsen. Die Kelchblätter sind vier bis fünf Zellen stark und enthalten eine Schicht aus Fasern, die nicht mit den Gefäßbündeln assoziiert sind. Die Farbgebung der weiblichen Blüten scheint der der männlichen zu folgen. Bei A. deltoidea sind sie jedoch, im Gegensatz zu den männlichen orangefarbenen Blüten, grünlich.
Jede Blüte enthält sechs sterile Staubblätter, Staminodien. Sie sind zu einer spitz gelappten becherförmigen Hülle verwachsen, die wiederum mit der unteren Hälfte der Kelchblätter verwachsen ist. Die drei kugeligen Fruchtblätter sind verwachsen (synkarp) und der Griffel, dessen Länge etwa der der Staubfäden entspricht, trägt eine dreiteilige Narbe. Gerade unterhalb der Narben und an der Öffnung des zentralen Griffelkanals, zwischen den Narben, werden kleine Mengen Nektar sekretiert.
Früchte und Samen
Die Früchte der Stachelpalmen sind bei den meisten Arten runde rote, einsamige Steinfrüchte, mit einem dicken, harten, den Samen umgebenden verholzten Kern, (Endokarp). Das Endokarp ist zwischen 0,5 und zwei Millimeter dick und von brauner oder schwarzer Farbe. Es trägt drei distinkte Keimporen, die jeweils von runden, aufliegenden Fasern in einem sternförmigen Muster umringt sind. Die mittlere Fruchtwand (Mesokarp) ist fleischig und saftig. Der Durchmesser der Früchte schwankt zwischen fünf und 25 Millimetern und ist recht klein im Vergleich zu anderen Cocoeae.
Variationen ergeben sich zum Beispiel bei A. macroloba mit ellipsoiden Früchten, einige Arten, wie zum Beispiel A. grandis, bilden Früchte mit schnabelförmigen Auswüchsen. Auch die Farbe weicht bei einigen Arten von rot ab, so hat A. grandis mattgrüne, und andere Arten, wie zum Beispiel A. verrucosa, weiße Früchte. Purpurne Früchte finden sich zum Beispiel bei A. hirsuta.
Die Samenschale ist dünn. Das Nährgewebe (Endosperm) ist weiß und homogen, oft mit einer unregelmäßigen Höhlung im Inneren. Der Geschmack des Endosperms ist süß und erinnert an Kokos, der Ölgehalt schwankt innerhalb der Gattung stark und beträgt zum Beispiel bei A. horrida 37 %, bei A. minima aber 65 %. Wesentlicher Bestandteil des Öls ist Laurinsäure, bei A. horrida fast 63 %. Der Embryo ist von hellbrauner Farbe, hat eine umgekehrt konische Form und ist 0,5 bis ein Millimeter lang. Seine Spitze zeigt zu einer der drei Keimporen.
Aus den Früchten von A. horrida wurde neben iso-Rhapontigenin, Piceatannol und Luteolin, Aiphenol, eine neue Stilben-Verbindung isoliert, die als Inhibitor von Cyclooxygenase-Reaktionen fungiert.[2]
Verbreitung
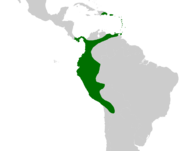 Natürliches Verbreitungsgebiet der Gattung
Natürliches Verbreitungsgebiet der Gattung Verteilung der Höhenlagen der Verbreitungsgebiete der Aiphanes-Arten
Verteilung der Höhenlagen der Verbreitungsgebiete der Aiphanes-ArtenStachelpalmen sind auf den Kleinen Antillen, sowie von Venezuela bis Bolivien entlang der Anden verbreitet. A. hirsuta subsp. hirsuta erreicht Panama, ansonsten fehlt die Gattung in Mittelamerika. Die Ostgrenze des Verbreitungsgebiets wird durch die Westgrenze des Amazonasbecken markiert und erreicht in einem sehr dünnen Streifen an der Grenze zu Peru Brasilien. Angebliche Funde in Guyana und aus dem Süden Venezuelas konnten nicht bestätigt werden.
Das Mannigfaltigkeitszentrum der Gattung, das heißt das Gebiet mit der größten Artenvielfalt, liegt in Westkolumbien und Ecuador, ein Unterzentrum findet sich im nordöstlichen Peru. Die am weitesten verbreitet Art ist A. horrida, die sich von Trinidad bis Bolivien, mit einer Lücke von Zentralkolumbien bis Zentralperu findet. Viele andere Arten besitzen nur ein kleines oder sehr kleines Verbreitungsgebiet.
In Tansania in Afrika existieren neophytische Vorkommen von A. horrida, die sich zwar wild verbreiten, aber nicht als invasiv eingestuft werden.[3]
Stachelpalmen können Höhenlagen bis 2800 Meter bewohnen, wobei im Flachland andere Arten als im Hochland verbreitet sind. Auch haben sich die Aiphanes Arten an sehr verschiedene Standorte angepasst und es existieren sowohl auf sehr feuchte, als auch auf sehr trockene Bedingungen spezialisierte Arten. Auch verhalten sich die Pflanzen im offenen Gelände anders als in dichtbewachsenen Waldgebieten. Während zum Beispiel A. ulei, A. weberbaueri, A. parvifolia oder A. tricuspidata im offenen Gelände nicht überleben können, passen sich beispielsweise A. erinacea oder A. hirsuta an die veränderten Bedingungen an und bilden viele kurze Stämme, und viel mehr Blütenstände.
Gefährdung
Wie bei vielen Gattungen mit stark spezialisierten oder endemischen Arten sind viele Arten gefährdet. Die International Union for Conservation of Nature and Natural Resources (IUCN) listet sechs der 25 Arten in einer Gefährdungskategorie auf ihrer Roten Liste.
Drei Arten gelten als „stark gefährdet“ (endangered) (A. grandis, A. leiostachys und A. verrucosa) und weitere drei als „gefährdet“ (vulnerable) (A. chiribogensis, A. duquei, A. lindeniana). Bei drei Arten (A. chiribogensis, A. grandis, A. verrucosa) verschlechtert sich die Lage beständig und steht eine weitere Verschlechterung zu befürchten. Vor allem A. grandis ist stark gefährdet, da keine der verbliebenen Populationen in Ecuador sich innerhalb eines Schutzgebiets befindet. Zumindest A. erinacea ist ebenfalls stark gefährdet, auch wenn die Art nicht auf der Roten Liste gelistet wird.[4]
Bei allen Arten ist Habitatzerstörung der Hauptgrund für die Gefährdung. Standorte werden vor allem durch Rodung und Urbarmachung des Landes zerstört. Für A. erinacea konnte gezeigt werden, dass die Art nur in unberührten Wäldern keimt, in auch nur leicht durchforsteten Wäldern aber eingeht, dies gilt wahrscheinlich auch für andere Stachelpalmenarten.[4].
Ökologie
Die meisten Arten blühen das ganze Jahr über und in einer einzigen Population können Individuen in allen Stadien des Knospens, Blühens und der Fruktation zur selben Zeit gefunden werden, oft finden sich sogar an einem einzigen Individuum mehrere Stadien zur selben Zeit. Dies erklärt sich vor allem aus den relativ gleichmäßigen klimatischen Bedingungen in großen Teilen des Verbreitungsgebiets. Es existieren jedoch auch einzelne Populationen, die sich parallel saisonal entwickeln, wie eine Population von A. horrida bei Canavi in Bolivien oder Populationen in sehr großen Höhenlagen.
Die Anthese dauert mindestens 80 Tage, gemessen bei A. chiribogensis und eine neue Blüte entsteht binnen etwa 25 Tagen nach dem Ende der Anthese der alten Blüte, gemessen bei A. eggersii.
Bestäubung
Die Blütenstände sind nichtüberlappend proterandrisch, das heißt, dass die männlichen Blüten den Pollen zu einem Zeitpunkt entlassen, zu dem die Narben der weiblichen Blüten noch nicht belegungsfähig sind. Ein Individuum hat nur selten mehr als einen Blütenstand zur selben Zeit – so wird Selbstbestäubung verhindert.
Die Stachelpalmenarten werden von verschiedenen Insekten bestäubt (Entomophilie), bei einigen Arten spielt auch Windbestäubung (Anemophilie) eine Rolle. Finn Borchsenius untersuchte die Bestäubung von A. chiribogensis, A. eggersi und A. erinaceae in Westecuador. In den männlichen Blüten von A. eggersii fand er viele Larven von Kleinschmetterlingen (Microlepidoptera), er schloss jedoch auf Bestäubung durch Bienen (Apiformes) und Wind. Die Blüten von A. erinacea werden von hunderten von Fliegen, vor allem Taufliegen (Drosophilidae), Schwebfliegen (Syrphidae), Gnitzen (Ceratopogonidae) und Blattkäfern (Chrysomelidae) besucht, die wahrscheinlich auch die Bestäubung übernehmen. Bienen beobachtete er hier nicht. Die Blüten von A. chiribogensis werden von deutlich weniger Insekten besucht. Hier finden sich Taufliegen, Pilzmücken (Mycetophilidae), Trauermücken (Sciaridae), Gallmücken (Cecidomyiidae), Gnitzen und Kleinschmetterlinge. Bienen oder Schwebfliegen fehlen.[5]
Die Blüten von A. grandis und A. minima duften süßlich, um Insekten anzulocken. Eine Analyse des Dufts der männlichen Blüte von A. minima ergab 15 Inhaltsstoffe. Die Hauptbestandteile sind Pentadekan (75,5 %), Tetradekan (3,9 %), 1,3,7-Nonatrien Linalool (1,2 %) und Dihydro-β-Jonon (1,2 %).[6]
A. horrida wird von Stachellosen Bienen (Meliponini) und Schnabelkerfen (Hemiptera) besucht, die Bestäubung findet durch Bienen statt. Glanzkäfer (Nitidulidae) und Rüsselkäfer (Curculionidae) finden sich ausschließlich in männlichen Blüten.
Insgesamt werden wahrscheinlich die Arten mit großen, weißen oder gelben Blüten und linearen Antheren vor allem durch Bienen, und die Arten mit kleinen, zumeist weiß bis purpurnen Blüten mit kleinen, ovalen Antheren vor allem durch Fliegen bestäubt.
Samenausbreitung
 Der Fettschwalm (Steatornis caripensis) frisst die Früchte von A. horrida und breitet die Samen aus.
Der Fettschwalm (Steatornis caripensis) frisst die Früchte von A. horrida und breitet die Samen aus.Die reifen Früchte von A. horrida werden von Eichhörnchen (Sciurus) gefressen, die den Stamm trotz der vielen Stacheln ersteigen können. Die Früchte sind energiereich, enthalten viele Vitamine und werden wahrscheinlich auch von vielen anderen Tieren gerne konsumiert. Die Stacheln an der ganzen Pflanze dienen insgesamt dem Schutz vor Pflanzenfressern und Tieren, die die Stämme erklettern wollen, um an die Früchte zu gelangen. Die hellroten Früchte von A. horrida werden auch vom Fettschwalm (Steatornis caripensis) gefressen, der sie im ganzen verschluckt und die Samen ausbreitet (Endochorie).
Verwendung
Die Früchte von A. horrida werden unter dem Namen Corozo oder Mararay auf vielen Märkten in Kolumbien angeboten und finden sich sogar in Supermärkten in Medellín. Sie werden roh gegessen. Kandiert sind die Früchte als Süßigkeiten in den Anden sehr beliebt.[7] A. horrida ist die einzige Art der Gattung die wegen ihrer Früchte kultiviert wird, die Früchte der anderen Arten werden ausschließlich in der Natur gesammelt.[8]
Die Früchte von A. linearis sind ebenfalls schmackhaft und werden in Kolumbien gegessen.[9] Die Fruchtkerne von A. minima sind essbar, und werden als Nüsse gehandelt.[10]
Trotz der Stacheln werden A. horrida und A. minima gelegentlich als Solitärpflanze in Gärten verwendet. In Botanischen Gärten sind die beiden Arten weit verbreitet. Das Holz von Stachelpalmen hat keinerlei wirtschaftliche Bedeutung.
Botanische Geschichte und Etymologie
Exemplare der Gattung Aiphanes wurden erstmals von Charles Plumier gesammelt, einem französischen Missionar und Botaniker, der zwischen 1689 und 1695 drei Reisen in die Karibik unternahm. Er fertigte Zeichnungen an und beschrieb zwei Arten, die er Palma dactylifera, aculeata, fructu corallino, major und Palma dactylifera, aculeata, fructu corallino, minor nannte. Beides waren Exemplare der heutigen Art A. minima. Dieselbe Art wurde dann nochmals im Jahr 1763 von Nikolaus Joseph von Jacquin als Palma grigri martinicensibus beschrieben.
1779 fertigte José Mutis eine sehr genaue Beschreibung der Art an, die heute als A. lindeniana bekannt ist. Im Jahr 1791 beschrieb der Deutsche Joseph Gärtner die Samen von A. minima in seinem Buch: „De fructibus et seminibus plantarum“ unter dem Namen Bactris minima – minima ist damit das älteste anerkannte Art-Epitheton einer Aiphanes-Art.
Der Gattungsname Aiphanes wurde erstmals 1801 in einer Vorlesung an der Königlichen Akademie der Wissenschaften in Berlin von Karl Ludwig Willdenow verwendet.[11] Der Name setzt sich aus dem altgriechischen αεί, ai (= immer) und φανερός, phaneros (= offenkundig, sichtbar, auffällig) zusammen. Ironischerweise sind Stachelpalmen üblicherweise nur schwierig in dichter Vegetation auszumachen und wurden aus diesem Grund auch nur selten in Herbarien gesammelt. Wahrscheinlich bezieht sich der Name eher auf die auffällige Erscheinung der Pflanzen.[12]
Willdenow beschrieb nur eine Art der Gattung A. aculeata, was heute ein Synonym von A. horrida ist. Das holotypische Exemplar, war von Franz Bredemeyer in Caucagua, Venezuela gesammelt worden. Bredemeyer ging dann nach Schönbrunn und nahm entweder den Holotypus oder Samen dieses Exemplars mit. 1809 beschrieb Joseph Franz von Jacquin denselben Holotypus oder aber ein nachgezogenes Exemplar unter dem Namen Caryota horrida – heute ist das Epipheton horrida als das gültige anerkannt.[13]
Zwischen 1794 und 1816 wurden mehrere Palmen unter dem Gattungsnamen Martinezia beschrieben – die Gattung war aber inkonsistent und wurde 1847 von Carl Friedrich Philipp von Martius mit Aiphanes synonymisiert. Da Martinezia der ältere Name war, wurde der Gattungsname Aiphanes bis 1932 nicht mehr verwendet. Die Gattung Martinezia enthielt aber viele Palmen, die nicht mit den Stachelpalmen gleichzusetzen sind. Carl Burret führte deswegen 1932 wieder den Namen Aiphanes ein.[14] Der Großteil der Arten aus Martinezia wurde zur Gattung der Kohlpalmen (Euterpe) gestellt, weshalb Martinezia heute als Synonym zu Euterpe gilt. Die meisten von Burret beschriebenen Arten wurden von Wilhelm Kalbreyer gesammelt, der zwischen 1877 und 1881 Nordkolumbien bereist hatte und eine umfangreiche Sammlung von Palmen mitbrachte, aus der 69 neue Palmenarten beschrieben werden konnten.[15]
Zwischen 1932 und 1996 wurden 15 weitere Stachelpalmenarten beschrieben, was die Anzahl der Arten auf 47 steigerte. 1996 veröffentlichten Finn Borchsenius und Rodrigo Bernal eine umfangreiche Monographie über die Gattung, in der die Anzahl der Arten auf 22 gesenkt wurde. Seither wurden allerdings drei neue Arten beschrieben.[16][17][18]
Systematik
Äußere Systematik
Uhl und Dransfield ordneten die Stachelpalmen zusammen mit den Gattungen Gastrococos, Acrocomia, Astrocaryum, Desmoncus und Bactris 1987 in die Tribus Bactridinae. Gemeinsames Merkmal der Gattungen sind Stacheln an zumindest Teilen der Pflanzen und das neotropische Verbreitungsgebiet.[19] Neben den bestachelten Palmen dieser Tribus, existieren aber auch noch andere Gattungen bestachelter Palmen nicht nur in der Neotropis, diese sind mit der Gattung Aiphanes aber nicht näher verwandt.[20]
Eine molekulargenetische Untersuchung aus dem Jahr 2002 ergab folgendes Kladogramm, dass die Verwandtschaftsverhältnisse innerhalb der Subtribus Cocoeae darstellt.[21]
┌───────── Gastrococos ┌─┤ │ └───────── Acrocomia ─┤ │ ┌───────── Astrocaryum └─┤ ┌────── Aiphanes └──┤ ┌─── Desmoncus └──┤ └─── BactrisInnere Systematik
Heute gehören 25 anerkannte Arten zur Gattung.[13] Unklar ist die Stellung von Aiphanes leiospatha Burret, die Art wird als „Unplaced Name“ geführt.[22]
Die Gattung Aiphanes ist eine morphologisch klar umrissene Einheit. Ihre Stellung als monophyletische Gruppe wird daher nicht angezweifelt.
Carl Burret teilte die Gattung 1992 in zwei Untergattungen Macroanthera und Brachyanthera. Er unterschied diese vor allem anhand von morphologischen Merkmalen der Blüten, vor allem der Länge der Antheren und der Stellung der Blütenstände.[14] Diese Merkmale sind aber so vage, dass eine eindeutige Zuordnung nicht möglich ist. Dennoch scheinen die drei Arten in der Untergattung Macroanthera nah verwandt zu sein, wobei aber nicht klar ist, ob diese Gruppe monophyletisch ist.[23] Heute wird der Einteilung in der Regel nicht mehr gefolgt – in Ermangelung einer neueren Untergliederung wird sie hier dennoch verwendet.
Die Arten der Gattung sind:
Sektionen und Arten der Gattung AiphanesLiteratur
Die Informationen dieses Artikels entstammen zum größten Teil den folgenden Quellen:
- Finn Borchsenius, Rodrigo Bernal: Aiphanes (Palmae). In: Finn Borchsenius, Rodrigo Bernal (Hrsg.): Flora Neotropica. 70, The New York Botanical Garden, New York, ISBN 0-89327-408-9, ISSN 0071-5794.
- Carmen Ulloa Ulloa, Peter Møller Jørgensen: Aiphanes. In: Arboles y arbustos de los Andes del Ecuador. 2004 (volltext).
Weblinks
- Aiphanes in der Roten Liste gefährdeter Arten der IUCN 2007
- Die Gattung in Ecuador auf palmbase.org
- Bilder vieler Arten bei fieldmuseum.org
Einzelnachweise
- ↑ M. A. Sowunmi: Pollen morphology of the palmae and its bearing on taxonomy. In: Review of Palaeobotany and Palynology. 13, Nr. 1, Februar 1972, S. 1–80 (doi:10.1016/0034-6667(72)90044-9).
- ↑ Dongho Lee, Muriel Cuendet, Jose Schunke Vigo, James G. Graham, Fernando Cabieses, Harry H. S. Fong, John M. Pezzuto, A. Douglas Kinghorn: A Novel Cyclooxygenase-Inhibitory Stilbenolignan from the Seeds of Aiphanes aculeata. In: Organic Letters. 3, Nr. 14, Juni 2001, S. 2169–2171 (doi:10.1021/ol015985j).
- ↑ Darwin Initiative (Hrsg.): Aiphanes horrida. In: Usambara Invasive Plants. (webseite ; Stand: 14. Dezember 2007).
- ↑ a b Jens-Christian Svenning: The effect of land-use on the local distribution of palm species in an Andean rain forest fragment in northwestern Ecuador. In: Biodiversity and Conservation. 7, 1998, S. 1529–1537 (doi:10.1023/A:1008831600795).
- ↑ Finn Borchsenius: Flowering biology and insect visitation of three Ecuadorean Aiphanes species. In: Principes. 37, 1993, S. 139–150.
- ↑ J. T. Knudsen, L. Tollsten. F. Ervik: Flower Scent and Pollination in Selected Neotropical Palms. In: Plant Biology. Nr. 3, 2001, S. 642–653 (doi:10.1055/s-2001-19366).
- ↑ Mark J. Plotkin, Lisa Famolare: Sustainable Harvest and Marketing of Rain Forest Products. Island Press, Washington, D.C. Juni 1992, ISBN 978-1-55963-169-3, S. 159.
- ↑ Geo Coppens d'Eeckenbrugge, Dimary Libreros Ferla: Aiphanes. In: Fruits from America, An ethnobotanical inventory. (website ; Stand: 11. Januar 2008).
- ↑ Food and Agriculture Organisation of the United Nations (FAO) (Hrsg.): Tropical Palms. 1998, ISBN 978-92-5-104213-7 (volltext).
- ↑ G. E. Wickens: Edible Nuts. Food and Agriculture Organisation of the United Nations (FAO), 1995, ISBN 978-92-5-103748-5.
- ↑ Karl Ludwig Willdenow: Über einige neue Südamericanische Palmen. In: Sammlung der Deutschen Abhandlungen Welche in der Königlichen Akademie der Wissenschaften zu Berlin Vorgelesen Wurde. 1803, S. 250f (volltext).
- ↑ Helmut Genaust: Etymologisches Wörterbuch der botanischen Pflanzennamen. 3. Auflage. Nikol, Hamburg 2005, ISBN 978-3-937872-16-2, S. 46.
- ↑ a b Rafaël Govaerts, J. Henderson; S. F. Zona; D. R. Hodel; A. Henderson; The Board of Trustees of the Royal Botanic Gardens, Kew (Hrsg.): World Checklist of Arecaceae. 2006 (website ; Stand: 14. Dezember 2007).
- ↑ a b M. Burret: Die Palmengattungen Martinezia und Aiphanes. In: Notizblatt des Königl. botanischen Gartens und Museums zu Berlin. 11, Nr. 107, 15. Dezember 1932, S. 557–577 (abstract).
- ↑ Rodrigo G. Bernal, Gloria Galeano-Garces, Andrew Henderson: Neotypification of Colombian Palms Collected by W. Kalbreyer. In: Taxon. 38, Nr. 1, Februar 1989, S. 98–107 (doi:10.2307/1220905).
- ↑ Carlos E. Cerón, Rodrigo Bernal: Una Nueva Especie de Aiphanes (Palmae) del Occidente de Ecuador. In: Caldasia. 26, Nr. 2, Dezember 2004, S. 433–438 (pdf).
- ↑ Gloria Galeano, Rodrigo Bernal: New Species and New Records of Columbian Palms. In: Caldasia. 24, Nr. 2, Dezember 2002, S. 277–292 (pdf).
- ↑ Santos Miguel Niño, L. J. Dorr, F. W. Stauffer: Una nueva especie de Aiphanes (Arecaceae) de la Cordillera de Mérida, Venezuela.. In: Sida, Contributions to Botany. 21, Nr. 3, 2005, S. 1599–1606 (abstract).
- ↑ Natalie W. Uhl, John Dransfield: Genera palmarum: a classification of palms based on the work of Harold E. Moore. Allen Press, Lawrence 1987, ISBN 978-0-938868-30-9.
- ↑ John Dowe: Spiny Palms. In: Palms & Cycads. Nr. 12, 1986 (website ; Stand: 11. Januar 2008).
- ↑ William J. Hahn: A phylogenetic analysis of the Arecoid Line of palms based on plastid DNA sequence data. In: Molecular Phylogenetics and Evolution. 23, Nr. 2, Juni 2002, S. 189–204 (doi:10.1016/S1055-7903(02)00022-2).
- ↑ R. Govaerts: Aiphanes leiospatha. In: The Board of Trustees of the Royal Botanic Gardens, Kew (Hrsg.): World Checklist of Arecaceae. 2006 (website ; Stand: 14. Dezember 2007).
- ↑ Borchsenius, Bernal. Seite 33
.jpg)
.jpg)

.jpg)
.jpg)



Wikimedia Foundation.