- Großhirnrinde
-
Die Großhirnrinde (lat. Cortex cerebri, kurz: Cortex) ist die äußere, an Nervenzellen (Neuronen) reiche Schicht des Großhirns (Telencephalon).
Obwohl das lateinische Wort cortex übersetzt eigentlich schlicht „Rinde“ bedeutet und fachsprachlich die (gesamte) Hirnrinde (Cortex) bezeichnet, wird „Cortex“ in der Fachsprache auch einschränkend für die Großhirnrinde verwendet. Infolgedessen wird auch das Adjektiv „cortikal“ – bzw. eingedeutscht „kortikal“ – (eig. „die (gesamte) Hirnrinde betreffend“) fachsprachlich für „die Großhirnrinde betreffend“ benutzt.
Die Großhirnrinde ist je nach Region nur 2 bis 5 Millimeter dick und ein Teil der grauen Substanz (Substantia grisea) des Großhirns. Die Nervenfasern der Neuronen der Großhirnrinde verlaufen unterhalb der Hirnrinde und bilden die weiße Substanz (Substantia alba) des Großhirns. Sie werden analog zur Rinde auch als „Mark“ bezeichnet. Großhirnrinde und Marklager bilden zusammen den Großhirnmantel (Pallium cerebri ). Innerhalb des subcortikalen Marklagers des Großhirns befinden sich weitere Abschnitte grauer Substanz des Großhirns, die Großhirnkerne (Basalganglien, Claustrum und Corpus amygdaloideum).
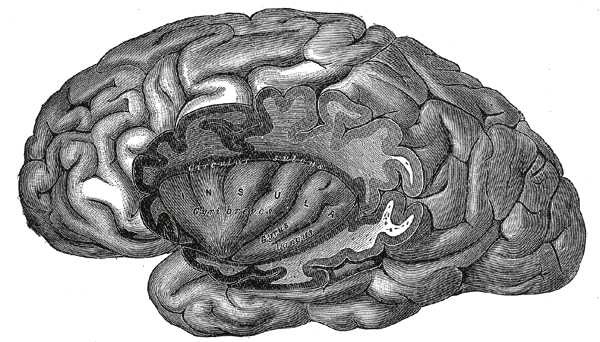
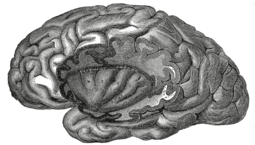 In dieser anatomischen Zeichnung sind Teile des linken Stirn-, Scheitel- und Schläfenlappens entfernt, so dass sich die oberflächliche (dunklere) Rinde und das hellere Marklager unterscheiden lassen.
In dieser anatomischen Zeichnung sind Teile des linken Stirn-, Scheitel- und Schläfenlappens entfernt, so dass sich die oberflächliche (dunklere) Rinde und das hellere Marklager unterscheiden lassen.
Inhaltsverzeichnis
Gliederung
Makroskopische Gliederung
Die noch heute gültige Terminologie der Hirnlappen und -windungen wurde 1869 von Alexander Ecker (1816–1887) vorgeschlagen.
Lappung
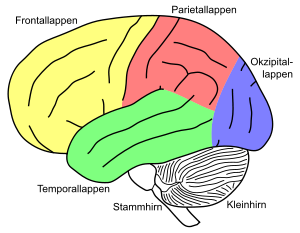 Unterteilung des Großhirns in Hirnlappen (Lobi)
Unterteilung des Großhirns in Hirnlappen (Lobi)Der Cortex lässt sich grob in fünf bis sechs Lappen (Lobi) einteilen, die durch tiefere Spalten (Fissurae) voneinander getrennt sind. Hiervon liegen an der Hirnoberfläche:
- Frontallappen oder Stirnlappen (Lobus frontalis)
- Parietallappen oder Scheitellappen (Lobus parietalis)
- Occipitallappen oder Hinterhauptslappen (Lobus occipitalis)
- Temporallappen oder Schläfenlappen (Lobus temporalis)
Bedeckt von Teilen des Frontal-, Parietal- und Temporallappens liegt seitlich der
- Insellappen (Lobus insularis)
Zusätzlich fassen einige Autoren gewisse entwicklungsgeschichtlich ältere Teile des Cortex (z. B. Gyrus cinguli und Hippocampus) zusammen als sechsten
- Limbischen Lappen (Lobus limbicus)
Die Untergliederung dieser Lappen ist nicht nur morphologisch, sondern auch funktionell von Bedeutung, da jedem Lappen ein spezielles primäres Verarbeitungsareal zukommt:
- Im großen Frontallappen liegen verschiedene Areale, von denen am wichtigsten die motorischen Zentren des Großhirns in und um den Gyrus praecentralis sind. In den rostralen (vorderen) Abschnitten liegt der präfrontale Cortex, der mit Handlungsplanung und -initiierung in Verbindung gebracht wird. Außerdem scheinen grundlegende Merkmale der Persönlichkeit hier lokalisiert zu sein.
- Nach hinten schließt sich der Parietallappen an, in dem das primäre sensible Zentrum liegt (Gyrus postcentralis).
- Am Pol des Occipitallappens liegt das Sehzentrum (Area striata)
- Auf der Innenseite des Temporallappens befindet sich das Hörzentrum (Area temporalis granulosa) in den sogenannten Heschlschen Querwindungen (Gyri temporales transversi)
- Die Inselrinde ist am wenigsten erforscht. Hier befindet sich unter anderem der primäre Geschmackscortex. Man vermutet hier auch das primäre Zentrum für basale Viszerosensibilität (Informationen aus den Eingeweiden).
- Im Limbischen Lappen (auch Limbisches System) werden alte Verschaltungsmuster prozessiert, deren prominenteste Vertreter Gedächtnisfunktionen und emotionale Prozesse sind.
Faltung (Gyrierung)
Die Großhirnrinde zeichnet sich bei vielen Säugetieren durch zahlreiche Windungen (lat. Griech. Gyri, Singular Gyrus), Spalten (lat. Fissurae, Sing. Fissura) und Furchen (lat. Sulci, Sing. Sulcus) aus. Die Faltung dient der Vergrößerung der Oberfläche: beim Menschen beträgt diese etwa 1800 cm². Bei der Furchung des Cortex unterscheidet man eine Primärfurchung, die bei allen Individuen annähernd gleich ist, von einer Sekundär- und Tertiärfurchung, die so individuell wie ein Fingerabdruck sein können.
Gefurchte Gehirne bezeichnet man als gyrenzephal. Bei einigen Kleinsäugern (beispielsweise Nagetiere, Igel) und bei Vögeln besitzt der Cortex keine Furchen (lissenzephales Gehirn).
Die Lobi und Gyri werden durch die Fissuren und Sulci voneinander getrennt. Deren wichtigste Vertreter sind:
- Fissura longitudinalis (auch Mantelkante), die den Spalt zwischen den beiden Hemisphären bildet. In die Fissura longitudinalis ragt die Falx cerebri.
- Sulcus centralis trennt den Frontal- und Parietallappen (Gyrus praecentralis beziehungsweise Gyrus postcentralis) und somit das primär motorische vom primär sensiblen Rindenfeld
- Sulcus lateralis (auch Sylvische Fissur) liegt oberhalb der Insula und trennt den Temporallappen von den darüber liegenden Frontal- und Parietallappen
- Sulcus parietooccipitalis zwischen Parietal- und Occipitallappen
- Sulcus calcarinus teilt innerhalb des Occipitallappens die primäre Sehrinde in einen oberen und unteren Anteil, welcher das jeweils gegenüberliegende Gesichtsfeld repräsentiert, das heißt oberhalb des Sulcus calcarinus das untere Gesichtsfeld, unterhalb des Sulcus calcarinus das obere.
Histologische Gliederung
Der Cortex kann nach zwei Gesichtspunkten unterteilt werden. Zum einen aufgrund seines histologischen Feinbaus in einen sechsschichtigen Isocortex und einen drei- bis fünfschichtigen Allocortex. Innerhalb der Cortexformen lassen sich Variationen im histologischen Feinbau feststellen, nach denen die Großhirnrinde des Menschen 1909 von Korbinian Brodmann in 52 Areale unterteilt (Brodmann-Areale oder -Felder) wurde. Ein anderer Gesichtspunkt ist das stammesgeschichtliche Alter der Hirnrinde, nachdem der Cortex in einen neueren Neocortex und die älteren Archicortex und Palaeocortex unterteilt wird.
Im Folgenden wird der histologische Aufbau des Isocortex beschrieben. Informationen zum Archicortex finden sich z. B. unter Hippocampus.
Zelltypen der Großhirnrinde
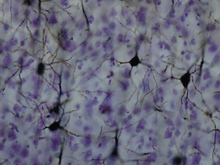 Sternzellen
SternzellenDie sechs Schichten der Großhirnrinde sind durch das Vorkommen bestimmter Zelltypen definiert. Viele dieser Zellen sind Interneurone (Calbindin-positive Zellen), die mit ihren Fortsätzen den Cortex nie verlassen, sondern nur zwischen den anderen Neuronen verschalten (meist GABAerg). Zwei Zelltypen sind jedoch für den Cortex spezifisch. Beide sind histologisch verwandt (Calmodulin-Kinase II (CaMK II)-positive Zellen) und entstehen vermutlich aus der gleichen Vorläuferzelle, weshalb sie im deutschen Sprachraum auch teilweise (verwirrend) als Pyramidenzellen und modifizierte Pyramidenzellen bezeichnet werden. Im angloamerikanischen Gebrauch werden letztere als bedornte Sternzellen bezeichnet.
- Pyramidenzellen sind die größten Zellen des Cortex. Sie werden benannt durch ihren pyramidalen Zelleib, dessen Spitze immer zur Cortexoberfläche zeigt. Sie haben einen apikalen und mehrere basale bedornte Dendriten, sowie ein basales Axon. Die Pyramidenzelle ist die efferente Zelle des Großhirns. Sie ist CaMK II-positiv und benutzt Glutamat als Neurotransmitter. Ihr Axon kann unterschiedlich lang sein. Im Falle der besonders großen Betz'schen Riesenzellen des Motorcortex, reicht es bis in das Rückenmark. Nach ihr ist dementsprechend die Pyramidenbahn benannt.
- Körnerzellen (Bedornte Sternzellen oder modifizierte Pyramidenzellen) haben einen etwas abgerundetereren Zelleib und viele bedornte Dendriten, von denen einer apikal aus der Zelle hervorgeht und die anderen quasi überall am Zelleib beginnen, weshalb sie im histologischen Bild an einen Stern erinnert. Sie sind ebenfalls glutamaterg und CaMK II-positiv und stellen die afferenten Zellen des Cortex dar, die Informationen aus anderen Hirnarealen und vor allem aus dem Thalamus empfangen.
- Interneurone sind in der Großhirnrinde zahlreich. Sie haben unterschiedliche Formen und sind meistens GABAerg und Calbindin-positiv. Ihre Fortsätze verlassen den Cortex nicht, da die Interneurone ausschließlich zwischen den anderen Zellen des Cortex kommunizieren. Sie werden in Doppelbuschzellen, Candelaberzellen, unbedornte Sternzellen, Fusiforme Zellen, Marinotti-Zellen, Horizontalzellen und Bipolare Zellen unterschieden.
Zusätzlich zu den Nervenzellen befinden sich im Cortex auch eine Vielzahl an Gliazellen. Sie bilden die Bindesubstanz zwischen den Neuronen und kommen verschiedenen Sonderaufgaben nach, für die sie jeweils spezialisiert sind:
- Oligodendrocyten bilden die Myelinscheiden um die Axone
- protoplasmatische und fibrilläre Astrozyten stützen das neuronale Gewebe, bilden die Blut-Hirn-Schranke (Membrana gliae limitans perivascularis), ebenso die Blut-Liquor-Schranke an der Hirnoberfläche (Membrana gliae limitans superficialis) und übernehmen zahlreiche Funktionen für die Ernährung und Erhaltung der Neurone
- Mikroglia sind die Abwehrzellen des Zentralnervensystem
Eine nennenswerte interzelluläre Matrix gibt es im Gehirn nicht, das gilt auch für die Großhirnrinde. Der Spalt zwischen Nerven- und Gliazellen ist nur 10 bis 50 Nanometer breit.
Laminierung
Durch das Vorhandensein der verschiedenen Zelltypen lässt sich der Cortex in verschiedene Schichten untergliedern
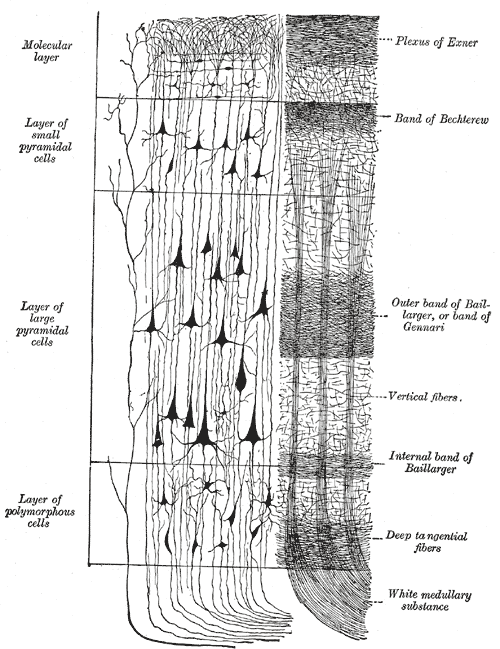
 Rindenschichten - links Zellfärbung, rechts Darstellung der Fasern.
Rindenschichten - links Zellfärbung, rechts Darstellung der Fasern.
In dieser Abbildung sind die Schichten II und III sowie IV und V zusammengefasst.Von außen nach innen sind dies im Isocortex:
- Lamina I (Stratum moleculare): Sie ist ein zellarmer Anteil des Cortex, in dem hauptsächlich Fasern und vereinzelte Interneurone zu finden sind. In der Embryonalentwicklung entsteht diese Schicht als erste, indem sich hier die ersten Neurone (Cajal-Retzius-Zellen) einlagern, die jedoch im späteren Entwicklungsverlauf Apoptose eingehen. Die anderen Schichten entwickeln sich invers: unter die Schicht I legt sich die Schicht VI, durch die dann die Neurone der Schicht V wandern, danach Schicht IV und so weiter, bis zum Schluss die Schicht II angelegt wird. In der Lamina I liegt ein ausgeprägtes Faserbündel, der Exner-Streifen.
- Lamina II (Stratum granulosum externum): Hier finden sich vor allem kleinere bedornte Sternzellen.
- Lamina III (Stratum pyramidale externum): Hier liegen kleinere Pyramidenzellen, sowie ein Faserzug, der Kaes-Bechterew-Streifen.
- Lamina IV (Stratum granulosum internum): In dieser Schicht liegen die größeren primärafferenten bedornten Sternzellen, die Projektionen aus anderen Hirnarealen enthalten. Dementsprechend ist diese Schicht in der Hör-, Seh- und sensiblen Rinde besonders stark ausgeprägt (granulärer Cortex), während sie in motorischen Rindenfeldern praktisch völlig fehlt (agranulärer Cortex). In der Lamina IV liegt ein weiteres Fasergeflecht, das äußere Band von Baillarger. In der Sehrinde ist dieses Band derart stark ausgebildet, dass es auch makroskopisch mit bloßem Auge sichtbar ist. Diesem hier als Gennari- oder Vicq d'Azyr-Streifen benannten Streifen verdankt die Sehrinde ihren Namen als Area striata.
- Lamina V (Stratum pyramidale internum): Sie ist der Sitz der großen Pyramidenzellen, die aus der Hirnrinde herausprojizieren. Im Motorcortex finden sich hier besonders große Pyramidenzellen (Betzsche Riesenzellen), während die Schicht im granulären Cortex vollkommen fehlt. Des Weiteren liegt hier der tiefste aller Fasergeflechte des Cortex, das innere Band von Baillarger.
- Lamina VI (Stratum multiforme): Hier liegen viele verschiedene Pyramiden- und bedornte Sternzellen, sowie zahlreiche Interneurone.
Neben den horizontalen Schichten ist der Cortex oftmals vertikal in Säulen organisiert. Diese kortikalen Säulen sind vor allem in den primären sensorischen Arealen ausgeprägt und zeichnen sich durch eine starke Konnektivität innerhalb einer Säule aus. Sie stellen damit sozusagen die elementaren Verarbeitungseinheiten (Module) der Großhirnrinde dar.
In einer großangelegten Studie untersuchten Chen et al. (2008) die Modularität eines kortikalen Netzwerks bestehend aus insgesamt 54 Hirnregionen. Im Zuge der statistischen Auswertung der erhobenen Daten konnte gezeigt werden, dass eine Vielzahl von Verknüpfungen zwischen den untersuchten Regionen besteht, und dass kortikale Dicke alleine die notwendige Information über die Modularität des Gehirns liefern kann.[1]
Interne Organisation
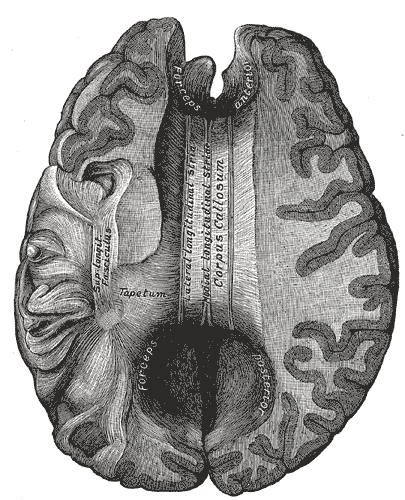
 Faserpräparat
FaserpräparatMan unterscheidet drei verschiedene Verlaufsformen der Axone:
- Assoziationsfasern verbinden verschiedene Areale innerhalb einer Hemisphäre. Man unterscheidet kurze Assoziationsfasern, die einen Gyrus mit dem nächsten verbinden (Fibrae arcuatae), mittellange (u. a. Fasciculus arcuatus und Fasciculus uncinatus) und lange Assoziationsfasern (vor allem Capsula externa, Fasciculus longitudinalis superior und Fasciculus longitudinalis inferior)
- Kommissurenfasern verbinden korrespondierende Areale gegenüberliegender Hemisphären. Wichtige Kommissuren sind das Corpus callosum, die Commissura anterior, Commissura posterior und die Fornix (Commissura fornicis).
- Projektionsfasern verbinden den Cortex mit tiefer liegenden Arealen. Die prominentesten Projektionsbahnen sind die Capsula interna als Teil der Pyramidenbahn und der Lemniscus medialis.
In der Makroskopie sind diese verschiedenen Bahnen deutlich sichtbar im Marklager des Großhirns organisiert. Von außen nach innen erkennt man hier kurze Assoziationsfasern (Capsula extrema), lange Assoziationsbahnen (Capsula externa) und ganz innen die Projektionsfasern der Capsula interna. Die gleiche Anordnung nach Länge und Typ haben die Fasern im Cortex. Wenn man sich klar macht, dass die Schichten II und IV afferent und die Schichten III und V efferent sind, ist es recht logisch nachvollziehbar, wie der Cortex intern organisiert ist:
- Kurze und mittellange Assoziationsfasern beginnen in Schicht III und enden in Schicht II
- Kurze Kommisurenfasern beginnen in Schicht III und enden in Schicht II
- Lange Assoziations- und Kommissurenfasern beginnen in Schicht V und enden in Schicht IV
- Lange afferente Projektionen enden in Schicht IV
- Lange efferente Projektionen beginnen in Schicht V
- Verbindungen aus allocorticalen Arealen (Riechhirn, Claustrum, Amygdala etc.) sowie Projektionen aus unspezifischen "Thalamuskernen enden in Schicht VI und teilweise in Schicht I.
Funktionelle Gliederung
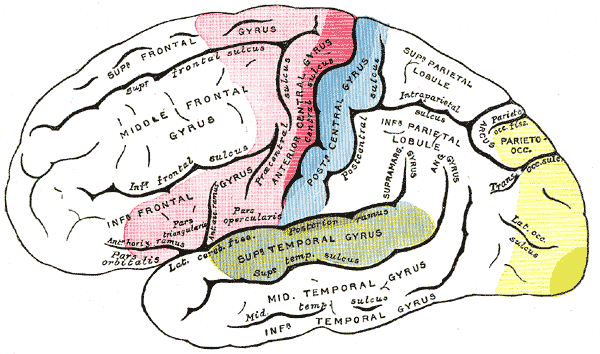
 Funktionelle Organisation der Großhirnrinde
Funktionelle Organisation der GroßhirnrindePrimär-motorisches Areal Prä/Supplementär-motorische Areale Primär-sensible Areale Sensible Assoziationsareale Sehfelder Hörfelder Weiterhin befinden sich in der Großhirnrinde die so genannten funktionellen Zentren, die in engem Zusammenhang mit den Brodmann-Arealen stehen. Die wichtigsten funktionellen Zentren sind die primären sensorischen und primären motorischen Areale. Das primäre motorische Areal, das Bestandteil des Motorcortex ist, liegt im Gyrus praecentralis (Brodmann-Areal 4). Gleich daneben liegt der primäre somatosensible Cortex im Gyrus postcentralis (Areale 1 bis 3). Im occipitalen Lappen formt der primäre visuelle Cortex in Area 17 den caudalsten (hintersten) Pol des Gehirns. Der primäre akustische Cortex findet sich in den Gyri temporales transversi (Areal 41). Zu den primären Arealen kommt meist eine ganze Reihe sekundärer Areale, die ebenfalls ausschließlich Informationen aus einer Modalität (Sehen, Hören, Motorik) bearbeiten. Diese Cortex-Regionen nehmen eine zentrale Stellung in der Verarbeitung und Bewusstwerdung neuronaler Impulse ein, dürfen jedoch nicht isoliert betrachtet werden, da das gesamte Nervensystem ein vielfach verschaltetes Netzwerk darstellt. Der Rest der Großhirnrinde wird vom Assoziationscortex eingenommen, also Arealen, die multimodalen Input bekommen und oftmals weder eindeutig sensorische oder eindeutig motorische Aufgaben haben. Heute weiß man, dass komplexe Fähigkeiten wie Motivation, Aufmerksamkeit, Kreativität, Spontaneität und beispielsweise auch die Verinnerlichung sozialer Normen von ihnen abhängen.
Verschaltung
Die Großhirnrinde erhält ihre zuführenden (Afferenz) Informationen vom Thalamus. Diese Informationen umfassen Sinneswahrnehmungen der verschiedenen Sinnesorgane. Bereiche, welche solche Informationen erhalten, werden als Sinnesbereiche oder Projektionszentren bezeichnet, z. B. der visuelle Cortex. Die beiden Hemisphären (linke und rechte) erhalten die Informationen der jeweils anderen Hälfte des Körpers, da die zuführenden Bahnen im Verlauf auf die Gegenseite kreuzen. Die Teile der Großhirnrinde, die Informationen über den Thalamus beziehen, werden als primäre Sinnesbereiche bezeichnet.
Weitere Bereiche erhalten Impulse von den primären Sinnesbereichen und kombinieren die Informationen unterschiedlicher Sinnesorgane. Diese assoziativen Bereiche nehmen bei allen Primaten, besonders beim Menschen, viel Raum ein.
Schließlich leiten die Assoziationsareale Informationen an die motorischen Bereiche weiter. Dort entstehen die Befehle für alle willkürlich steuerbaren Körperfunktionen und werden über die Pyramidenbahn als hauptsächlichem Output des Großhirns an die Peripherie weitergeleitet. Teile des Motorcortex sind eng mit den Basalganglien und dem Kleinhirn verschaltet.
Neben den Informationen, die von den Sinnesorganen über den Thalamus die Rinde erreichen, erhalten alle Bereiche des Cortex zusätzliche „unspezifische“ Erregungen aus den thalamischen Kerngebieten der Formatio reticularis. Diese Erregungen des aufsteigenden retikulären Aktivierungssystems sind rhythmisch, wobei ihre Frequenz mit dem Grad der Wachheit (Vigilance) veränderlich ist. Das Spektrum reicht von etwa 3 Hz im Tiefschlaf und Narkose bis ca. 40 Hz bei hellwacher Anspannung, z. B. beim Lesen.
Die Oszillationen des ARAS werden in einer schleifenförmigen Leitung zwischen Thalamus und den Basalganglien (Stratum, Pallidum, Nucleus caudatus, Putamen) erzeugt, sie bilden den natürlichen „Hirnschrittmacher“. Elektronische Hirnschrittmacher, die in letzten Jahren zur Behandlung der Parkinson-Krankheit entwickelt wurden, versuchen diese aktivierende und hemmende Funktion des ARAS zu ersetzen.
Die Evolution und Funktion des Großhirns
 Größenvergleich der Gehirne verschiedener Säugetiere. Man beachte auch die mit der evolutionären Entwicklungsstufe zunehmende Oberflächenfaltung (Gyrierung).
Größenvergleich der Gehirne verschiedener Säugetiere. Man beachte auch die mit der evolutionären Entwicklungsstufe zunehmende Oberflächenfaltung (Gyrierung).
Quelle: University of Wisconsin and Michigan State Comparative Mammalian Brain Collections and the National Museum of Health and Medicine.Das menschliche Gehirn ist keine Neuentwicklung der Natur. Es hat sich wie alle anderen Organe aus einfachen Formen entwickelt. Das Nervensystem entwickelt sich aus einer sehr einfachen Struktur, dem äußeren Keimblatt (Ektoderm). Dass ein Organ der Informationsverarbeitung aus der äußeren Grenzschicht entsteht, ist leicht verständlich, weil hier die Reize aus der Umwelt auftreffen. Erst im Lauf der Evolution wurden die empfindlichen Nervenverbände in die Tiefe des Neuralrohrs verlegt, weil sie dort besser geschützt sind. Die Verbindungen zur Außenwelt blieben über die nun spezialisierten Sinnesorgane bestehen.
Mit der Entstehung spezialisierter Sinnesorgane ist die Bildung einer Nervenzentrale verbunden, die den ganzen Körper einheitlich nach den Sinneseindrücken steuern kann. Weil sich schon früh in der Geschichte der Wirbeltiere Augen, Ohren und chemische Sinne (Geschmack, Geruch) ausbilden, ist das Gehirn aller Wirbeltiere in gleicher Art zur zentralen Integration dieser Sinne konstruiert.
Das Endhirn war zunächst Verarbeitungszentrum für das Geruchsorgan. Weil der Geruchssinn ein allgemeines Warn- und Reizsystem hoher Empfindlichkeit ist, aber wenig über die räumliche Situation bzw. den Ort der Reizquelle aussagt, ist für das Riechhirn eine Verbindung mit den optischen und akustischen Zentren des Mittelhirns notwendig, mit der alle Sinnesqualitäten auf einer gemeinsamen Ebene vereinigt werden.
Diese gemeinsame Ebene entsteht schon bei den Reptilien aus einer Erweiterung des Endhirns als Telencephalon oder rudimentärer Cortex. Bereits bei Fröschen und Salamandern ist diese Hirnstruktur für die Integration der verschiedenartigen Reize angelegt. Für die Umschaltung der Seh-, Tast- und Hörwelt vom Mittelhirn auf das Endhirn entwickelt sich ein Teil des Vorderhirns, das Zwischenhirn. Aus ihm entsteht der Thalamus, der aus mehreren Kerngruppen die spezifischen Signale des Mittelhirns zu spezifischen Regionen der Großhirnrinde sendet. Man bezeichnet diese Anordnung als ein Projektionssystem, die Anatomen nannten den Thalamus das „Tor zum Bewusstsein“.
Mit dem Wegfall des Schuppenkleides der Fische bzw. der Hornschuppen der Reptilien wurde bei den Säugetieren die ganze Haut zu einem empfindlichen Sinnesorgan, das ebenso über Projektionsbahnen im Cortex mit den übrigen Sinnesqualitäten in ganzheitliche Verbindung gelangt.
Eine Nervenzentrale, in der alle Qualitäten der Umweltsignale zusammengeführt werden, wäre nicht sinnvoll, wenn in ihr keine Befehle für die Reaktionen des Organismus gebildet und an die ausführenden Organe geleitet werden könnten. Weil das Geruchsorgan von Anfang an einen steuernden Zugriff auf komplexe Verhaltensweisen hat, kann das zum Integrationszentrum aller Sinne erweiterte Riechhirn auf diese Steuerungsbahnen zurückgreifen, um aus der Vereinheitlichung aller Empfindungen ganzheitliche Verhaltensschritte zu entwickeln.
Diese Integrationsleistung des Neocortex, die alle Sinne zu einem Ganzen verbindet und sinnvolle Verhaltensmuster daraus herstellt, ermöglicht bereits Ratten, Katzen usw. ein intelligentes Verhalten, das bei Insekten oder einfachen Organismen so nicht vorkommt. Dabei zeigt sich, dass schon Vögel und Mäuse ihr integratives Zentrum, die Hirnrinde, nicht nur als Kommandozentrale, sondern auch als besonders leistungsfähigen Informationsspeicher (Gedächtnis) nutzen können. Eine Fliege lernt es nie, den Zusammenstoß mit einer Fensterscheibe zu vermeiden, während ein Vogel nach einigen Erfahrungen einen vorsichtigen Umgang mit der durchsichtigen Wand lernt.
Nur Tiere, die über einen Cortex verfügen, können auch dressiert werden, das heißt, sie entwickeln ein Gedächtnis für sprachliche Anweisungen, die auch über die angeborenen Verhaltensmuster dominieren können. Deutlich ist diese Lernfähigkeit bei den Delphinen, die als Säugetiere mit einem mächtigen Cortex ausgestattet und gut dressierbar sind, während die relativ großhirnarmen Haie zur Dressur wenig geeignet sind.
Mit der Entwicklung des Cortex kommt zunehmend eine spielerische Phase der Jungtiere zum Vorschein, die als Lernphase der Hirnrinde zu verstehen ist und uns den Eindruck vermittelt, dass diese Tiere (z. B. Hunde, Katzen usw.) ähnliche geistige Zustände wie die Menschen empfinden.
Eine mächtige Entwicklung der Großhirnrinde wurde bei den Affen durch die Sonderstellung der Hände ausgelöst. Als bei den Säugetieren noch alle vier Extremitäten ausschließlich zur Fortbewegung dienten, genügten einfache Reflexmuster auf Rückenmarksebene dazu, den harmonischen Laufrhythmus zu steuern. Bei den Primaten geschieht ein Wandel der Fortbewegung, vom Vierfüßler zum Klettertier. Damit kommt es zu einer Umkonstruktion der vorderen Extremitäten, die zu Greifinstrumenten werden. Das alte Bewegungsmuster der Vierfüßler ist damit überfordert, aber die Großhirnrinde kann sich durch massives Wachstum den neuen Anforderungen der Handmotorik anpassen.
Zusätzlich ist bei den Säugern das Kleinhirn in Verbindung mit dem Gleichgewichtsorgan für die Ausführung komplizierter Bewegungsabläufe in das motorische System integriert. Aufrechtes Laufen auf zwei Beinen ist ohne diese Hirnstruktur nicht möglich. Die Zusammenarbeit zwischen Cortex und Kleinhirn lässt sich am Beispiel des Radfahrens so erklären: Die Entscheidung über Rechtskurve oder Bremsvorgang trifft der Cortex, während die Feinarbeit der Gewichtsverlagerung und viele automatische Bewegungsimpulse im Kleinhirn bearbeitet werden.
Bei den Affen hat sich die Stellung der Augen im Gesichtsfeld so geändert, dass immer ein räumliches Bild der Umwelt gesehen wird. Für die zentrale Auswertung der binokularen Bilder müssen neue Analysatoren in das System integriert werden, und auch dabei erweist sich die Großhirnrinde als anpassungsfähiges Integrationszentrum mit riesigem Speichervermögen für komplexe Information.
Mit dieser Ausstattung war spätestens Homo erectus für den aufrechten Gang in der Savanne gut gerüstet und konnte den Geruchssinn zu Gunsten der Fernsinne (Augen und Ohren) vernachlässigen. Der Cortex passte sich seinen neuen Anforderungen an, indem er seine Fläche durch Faltenbildung vergrößerte.
So weit ist das biologische Standardwissen detailliert erforscht und beweist, dass die Großhirnrinde von Anfang an für die Herstellung einer ganzheitlich vereinigten Projektion aller Umweltsignale und einer daraus basierenden Verhaltenssteuerung spezialisiert war und diese Aufgabe in der Evolution immer stärker ausdehnen konnte. Ein bisher noch unverstandener Speichermechanismus ist verantwortlich für die Gedächtnisfunktion dieser Integrationszentrale, die den Lebewesen neben der starren, genetischen Anpassung eine flexible Anpassung an beliebige neue Situationen ermöglicht.
Die ersten Menschen hatten mit diesem Gedächtnisorgan und einem verbesserten Kehlkopf die Grundlage für die Verfeinerung der äffischen Laut- und Gebärdensprache. Die veränderte Daumenstellung erleichterte den Gebrauch von Werkzeugen und sorgte für weitere Ausdehnung der Hirnrindentätigkeit.
Schon bei der Herstellung von Faustkeilen mit scharfen Klingen ergab sich eine Aufgabenteilung für die beiden Hände, indem eine Hand zum Festhalten und die zweite Hand für gestaltende Feinarbeiten bevorzugt wurden. Viele Tätigkeiten mit Werkzeugen fördern eine differenzierte Spezialisierung der Hände, und spätestens beim systematischen Training des Schreibens ist eine dominante Hand kaum noch zu vermeiden.
Dementsprechend unterscheiden sich die beiden Seiten der Hirnrinde im Lauf der Evolution und der individuellen Entwicklung zunehmend, und nur auf der Seite der schreibenden Hand wird zusammen mit den Buchstabenverbindungen auch die Artikulation der Sprache gründlich trainiert. Weil die Nervenbahn des rechten, schreibenden Armes im linken Cortex beginnt, liegen auch die Sprachzentren im linken Großhirn, das deshalb als die dominante Hemisphäre bezeichnet wird.
Die Evolution des Cortex ist nachvollziehbar. Es fehlt nur noch eine wissenschaftlich einleuchtende Erklärung für die Leistungsfähigkeit, die sich in den grauen Falten unter der Schädeldecke als Gedächtnis und Bewusstsein erleben und in Sprache ausdrücken lässt.
Siehe auch
- Blue Brain Projekt, dessen Ziel eine Computersimulation des Neocortex ist.
Literatur
- Wilder Penfield, Theodore Rasmussen: The Cerebral Cortex of Man. A Clinical Study of Localization of Function. The Macmillan Comp., New York 1950.
- Otto Detlev Creutzfeldt: Cortex cerebri. Springer 1983.
Einzelnachweise
- ↑ Zhang J. Chen, Yong He, Pedro Rosa-Neto, Jurgen Germann, Alan C. Evans: Revealing Modular Architecture of Human Brain Structural Networks by Using Cortical Thickness from MRI. In: Cerebral Cortex, 18(10) 2008, S. 2374–2381.


Wikimedia Foundation.