- Knöllchenbakterien
-
Rhizobien Systematik Klassifikation: Lebewesen Domäne: Bakterien Abteilung: Proteobacteria Alphaproteobacteria Ordnung: Rhizobiales Familie: Rhizobien Wissenschaftlicher Name Rhizobiaceae Conn 1938 Als Knöllchenbakterien oder Rhizobien (altgriechisch ῥίζα rhiza = Wurzel und βίος bios = Leben) werden bestimmte Bakterien aus der Familie der Rhizobiaceae bezeichnet. Sie gehören zur Klasse der Alphaproteobacteria. Rhizobien sind gramnegativ, können sich mittels einer polaren oder subpolaren Flagelle oder 2 bis 6 peritrich angeordneten Flagellen aktiv bewegen, sind aerob und chemoorganotroph auf der Basis einer Sauerstoffatmung.
Rhizobien sind häufige und verbreitete Bodenbakterien. Ihre Bedeutung liegt in ihrer Fähigkeit, mit Pflanzen aus der Familie der Hülsenfrüchtler (Fabaceae) eine mutualistische Symbiose einzugehen. Die Lebensgemeinschaft ist sehr eng und führt bei den Rhizobien zu umfassenden morphologischen und physiologischen Veränderungen, bei den Pflanzen zur Ausbildung spezieller Organe.
Der Zusammenhang zwischen Wurzelanschwellungen und dem Befall mit Bakterien wurde 1866 erstmals vom russischen Botaniker Michail Stepanowitsch Woronin am Beispiel der Lupine beschrieben, der auch den Begriff der Knöllchenbakterien prägte. Hermann Hellriegel und Hermann Wilfarth entdeckten 1886 die Symbiose der Leguminosen und Bakterien und deren Fähigkeit, elementaren Luftstickstoff in pflanzenverfügbare Stickstoffverbindungen zu überführen.Rhizobien besitzen die Fähigkeit, elementaren, molekularen Stickstoff zu binden, also zu reduzieren und in Verbindungen zu überführen und damit biologisch verfügbar zu machen. Dies ist ihnen jedoch nur in der Symbiose mit Pflanzen möglich. Unter natürlichen Bedingungen können weder Leguminosen noch Rhizobien allein molekularen Stickstoff fixieren. Die Symbiose ist sowohl von biologischer als auch von wirtschaftlicher Bedeutung.
Inhaltsverzeichnis
Etablierung der Symbiose
Pflanzenwurzeln geben verschiedene organische Verbindungen ab. Diese Exsudate dienen unter anderem der Herausbildung einer speziellen Mikroorganismengesellschaft aus Bakterien und Pilzen in der Rhizosphäre, also der unmittelbaren Umgebung der Wurzel. Die aktiv sich bewegenden Rhizobien werden durch Wurzelexsudate chemotaktisch angelockt. Sie sind unter bestimmten Voraussetzungen in der Lage, in die Wurzel einzudringen („Infektion“). Voraussetzung für eine erfolgreiche Infektion ist stets eine kompatible und hoch spezifische Erkennung zwischen Bakterien und Pflanzenzellen auf molekularer Ebene. Die Infektion beginnt immer an bestimmten Wurzelzellen, den so genannten Wurzelhaaren.
Erkennung, Anheftung und Infektion
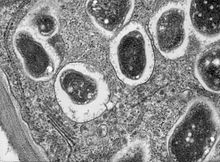 Mikroskopaufnahme eines Wurzelknöllchens der Sojabohne (Glycine max). Es enthält hunderte Millionen Bradyrhizobium-Bakterien.
Mikroskopaufnahme eines Wurzelknöllchens der Sojabohne (Glycine max). Es enthält hunderte Millionen Bradyrhizobium-Bakterien.
 Wurzeln des Sumpf-Hornklees (Lotus pedunculatus)
Wurzeln des Sumpf-Hornklees (Lotus pedunculatus) Wurzeln der Zaun-Wicke (Vicia sepium)
Wurzeln der Zaun-Wicke (Vicia sepium)Die ersten Schritte, Erkennung und Anheftung an die Wurzelhaarzelle, erfolgen durch spezielle Proteine auf der Oberfläche der Bakterien- und Pflanzenzelle. Eines der durch Rhizobien gebildeten Proteine ist Rhicadhesin, das an Calciumverbindungen auf der Pflanzenzelle bindet. Auch pflanzliche Lektine spielen eine Rolle, wie zum Beispiel das durch Weißklee gebildete Trifolin, das an spezielle Kohlenhydrate der äußeren Zellmembran bestimmter Rhizobienstämme bindet.
Nach der Anheftung beginnen die Rhizobien, in die Wurzelhaarzelle einzudringen. Die Penetration erfolgt immer an der Spitze des Wurzelhaares, das sich daraufhin auf charakteristische Weise einkrümmt und die Bakterienzelle umschließt.
Differenzierung der Bakterien und des infizierten Pflanzengewebes
Phenolische Wurzelexsudate (zum Beispiel Luteolin) aktivieren eine Reihe bakterieller Gene, die in Anlehnung an ihre Funktion als nod-Gene bezeichnet werden. nod steht für Nodulation, das bedeutet Knötchen- oder Knöllchenbildung. Einige der gebildeten Nod-Proteine werden nach außen abgegeben, wirken auf die Pflanzenzellen und sind für die erfolgreiche Interaktion zwischen Pflanzen- und Bakterienzellen notwendig.
Die Bakterien veranlassen die Wurzelhaarzelle, Cellulose zu bilden und induzieren einen Infektionskanal in Richtung Wurzelmitte. Entlang dieses Kanales werden anschließend benachbarte Zellen der Wurzelrinde infiziert. Die verstärkte Ausschüttung der bakteriellen nod-Faktoren veranlasst die benachbarten Zellen, sich zu teilen und sich zu vergrößern, die hierbei entstehenden Wurzelzellen sind außerdem oft polyploid. Dies führt zur Bildung knötchenartiger Wurzelverdickungen, der „Wurzelknöllchen“, in denen sich die Rhizobien befinden.
Zunächst vermehren sich die schlanken, stäbchenförmigen Bakterien. Anschließend beginnen die meisten, sich in verdickte, unförmige und verzweigte Zellen, so genannte Bacteroide oder Bacterioide zu verwandeln. Diese werden von den infizierten Pflanzenzellen in Membranen eingehüllt und bilden Zellorganellen, die als Symbiosomen bezeichnet werden. Es gibt Hinweise darauf, dass jedes Wurzelknöllchen aus der Infektion durch ein einziges Bakterium hervorgeht. Die Bacteroide eines Knöllchens wären in diesem Falle Klone. Den Bacteroiden fehlt die für Proteobakterien typische äußere Zellmembran. Sie sind außerdem nicht mehr in der Lage, sich zu vermehren oder sich in die ursprünglichen, vermehrungsfähigen Zellen zurückzuverwandeln. Mit fortschreitendem Alter nimmt der Verzweigungsgrad der Bacteroide zu.
Fixierung des elementaren, molekularen Stickstoffs (N2)
Die Enzyme für die N2-Fixierung besitzen nur die Rhizobien, nicht die Pflanzen. Das wichtigste Enzym, die molybdänhaltige Nitrogenase, ist jedoch in hohem Maße sauerstoffempfindlich. Bereits geringe Sauerstoffkonzentrationen inaktivieren das Enzym irreversibel. Die Bacteroide sind ihrerseits jedoch nicht in der Lage, völlig ohne Sauerstoff zu leben. Für die erfolgreiche Stickstoff-Fixierung muss die Sauerstoffkonzentration in den Wurzelknöllchen also genau ausbalanciert sein. Diese Funktion übernimmt die Pflanzenzelle, indem sie ein eisenhaltiges Protein in den Knöllchen bildet. Dieses Leghämoglobin bindet überschüssigen Sauerstoff, hält dessen Niveau konstant und erfüllt somit eine Sauerstoffpuffer-Funktion. Das Leghämoglobin ähnelt in seinem Aufbau stark dem gleichfalls sauerstoffbindenden Bluthämoglobin (Es bindet aber lediglich ein Molekül Sauerstoff und nicht vier) und färbt das Gewebe aktiver Wurzelknöllchen blassrosa bis blutrot.
Die Spaltung der Stickstoff-Dreifachbindung zur anschließenden Fixierung erfordert sehr viel Energie. Um ein Molekül Stickstoff zu spalten, werden 16 ATP und 4 NADH benötigt. Um 16 ATP zu generieren, müssen mindestens vier Moleküle Sauerstoff in der Atmungskette reduziert werden. Das heißt, eine optimale Fixierung bräuchte ein Gasgemisch aus vier Teilen Sauerstoff und einem Teil Stickstoff. Bekanntermaßen ist das Verhältnis dieser beiden Gase in der Atmosphäre aber umgekehrt. Sauerstoff ist in den Zellen also "Mangelware". Tatsächlich erhöht das Leghämoglobin den Sauerstofffluss zu den Bakterioiden aufgrund der deutlich besseren Löslichkeit des Leghämoglobin-Sauerstoff-Komplexes gegenüber dem einfachen Sauerstoffmolekül und das trotz der herabgesetzten Diffusionsgeschwindigkeit aufgrund des enormen Zuwachses an Volumen und Masse.
Das sauerstoffempfindliche, stickstofffixierende Enzym, die Nitrogenase, wird von Bakterioiden selbst geschützt, da die Atmungskette, in der das ATP gebildet wird, in seiner Membran, also außen, lokalisiert ist. Dort wird der Sauerstoff verbraucht und kann so der Nitrogenase im Inneren der Bakterioiden nicht schaden, es sei denn, es kann aus irgendwelchen Gründen nicht ausreichend Sauerstoff an der Atmungskette verbraucht werden.
Bis vor Kurzem nahm man an, dass die Komponenten für die Bildung des funktionsfähigen Hämoglobins kooperativ hergestellt würden: Die Proteinkomponente, also das Globin, würde durch die Pflanzenzelle synthetisiert, während der Häm-Anteil mit dem Porphyrinring durch die Bacteroide gebildet und in die Pflanzenzelle exportiert würde. In der Pflanzenzelle erfolge dann unter Einbau des Eisen-Ions die Assemblierung des vollständigen Leghämoglobins. Neuere Erkenntnisse zeigen jedoch, dass sich der Bauplan für Leghämoglobin ausschließlich auf der DNA der Pflanze befindet und das Leghämoglobin auch nur dort synthetisiert wird.
Stoffaustausch
Die Bacteroide sind ernährungsphysiologisch von der Pflanze abhängig. Die Pflanze stellt zur Deckung des Energiebedarfes für die Stickstoff-Fixierung organische Kohlenstoffverbindungen wie Succinat, Malat und Fumarat, also Zwischenprodukte des Citratzyklus zur Verfügung. Diese Stoffe stammen ursprünglich aus der Photosynthese der Pflanze. Durch den Abbau der Verbindungen gewinnen die Bacteroide Energie in Form von ATP und Reduktionsmittel, in diesem Falle Pyruvat, zur Spaltung und Reduktion des Stickstoffmoleküls. Diese Reaktion ist außerordentlich energieaufwändig. Zur Umwandlung eines einzigen N2-Moleküls sind mindestens 16 Moleküle ATP notwendig. Die Bacteroide liefern im Gegenzug an die Pflanzenzelle hauptsächlich Ammoniak (NH3) als erstes stabiles Produkt der Stickstoff-Fixierung, das im wässrigen Milieu zu Ammoniumionen (NH4+) umgesetzt wird. Ammoniak ist jedoch ein starkes Zellgift. Um eine Ammoniak-Anreicherung in den Pflanzenzellen zu vermeiden, wird dieser sofort zur Synthese von Glutamin und Glutaminsäure weiterverwendet (Ammoniak-Assimilation).
Genetik und Kompatibilitätsgruppen
Die bakteriellen Gene für die Etablierung der Symbiose und auch für die Stickstoff-Fixierung befinden sich in der Regel nicht im bakteriellen Genom, sondern auf einem Plasmid, dem sym-Plasmid.
Die Gene, die für die Knöllchenbildung notwendig sind und vorrangig auf die Pflanzenzellen wirken, werden als nod-Gene bezeichnet (nod steht für Nodulation – Knötchenbildung). Die Gene für die Stickstoff-Fixierung werden nif-Gene genannt (nif steht für nitrogen fixation – Stickstoff-Fixierung).
Daneben sind Gene vorhanden, die für die hoch spezifische Erkennung zwischen Bakterien- und Pflanzenzelle notwendig sind: Rhizobien gehören immer einer speziellen Kompatibilitätsgruppe an. Ein bestimmter Bakterienstamm kann unter natürlichen Bedingungen meist nur mit einer bestimmten Pflanzenart eine Symbiose eingehen. Bakterien, die sich zum Beispiel auf Klee (Trifolium sp.) spezialisiert haben, können mit anderen Leguminosen keine erfolgreiche Symbiose etablieren. Man spricht in diesem Falle von „Biovaren“: Rhizobium leguminosarum biovar trifolii ist ein Stamm, den man nur an Klee findet. Überträgt man jedoch die entsprechenden Kompatibilitätsgene, können auch andere Pflanzen als Symbionten dienen.
Ökologie
Stickstoff ist für alle Organismen essentiell. Er ist ein wesentlicher Bestandteil von Aminosäuren und damit von Proteinen, aber auch von Nucleinsäuren, also DNA und RNA.
Tiere gewinnen ihren Stickstoff meist durch Aufnahme kompletter Aminosäuren, indem sie sich von anderen Organismen ernähren. Sie verfügen daher über ausreichende Stickstoffquellen.
Die meisten Pflanzen und viele Mikroorganismen können hingegen nur anorganischen Stickstoff assimilieren. Gebundener, also in Verbindungen vorliegender und damit biologisch verfügbarer Stickstoff kommt als Nitrat, Harnstoff oder Ammonium in Gewässern und Böden vor. Solche Stickstoffverbindungen sind unter natürlichen Bedingungen aber meist knapp und limitierend für das Wachstum dieser Organismen. Der größte Stickstoff-Vorrat befindet sich in elementarer, molekularer Form (N2) in der Luft (78 Volumenprozent) und gelöst im Wasser, ist in dieser Form für Tiere, Pflanzen, Pilze und die meisten Mikroorganismen aber nicht nutzbar. Nur einige Bakterien und Archaeen verfügen über die Enzyme, mit deren Hilfe N2 reduziert und in eine Form umgewandelt werden kann, die für andere Organismen nutzbar ist (Stickstoff-Fixierung). Die bakterielle Stickstoff-Fixierung ist damit von grundlegender Bedeutung für das Leben und den globalen Stickstoffkreislauf.
Durch die Symbiose mit Rhizobien wird für die Pflanzen indirekt N2 verfügbar. Leguminosen haben deshalb auf stickstoffarmen Böden einen klaren Selektionsvorteil. Viele Schmetterlingsblütler spielen außerdem eine wichtige Rolle als Pionierpflanzen auf Sand und Schutt, Halden und Kahlschlägen.
Bedeutung der Symbiose
Man schätzt die Menge des jährlich fixierten Stickstoffs auf über 120 Millionen Tonnen. Pflanzen, die auf diese Weise auf weniger wertvollen Böden gedeihen, haben großen Anteil an der Landwirtschaft und unserer Nahrungsgrundlage. Leguminosen und deren Früchte sind häufig proteinreich, was sicher auf die gute Versorgung mit Stickstoff zurückzuführen ist. Daneben werden Leguminosen als Gründüngung zur natürlichen Anreicherung des Bodens mit Stickstoff kultiviert.
Beispiele für Pflanzengattungen, die eine Symbiose mit Rhizobien eingehen können:
Nutzpflanzen (Auswahl) Soja (Glycine sp.)
Erbse (Pisum sp.)
Linse (Lens sp.)
Kichererbse (Cicer sp.)
Bohne (Phaseolus sp.)
Ackerbohne (Vicia faba)
Klee (Trifolium sp. )
Erdnuss (Arachis sp.)
Luzerne (Medicago sp.) Sojabohne (Glycine max), erntereife Pflanzen
Sojabohne (Glycine max), erntereife Pflanzen Ackerbohne (Vicia faba)
Ackerbohne (Vicia faba) Weiß-Klee (Trifolium repens)
Weiß-Klee (Trifolium repens) Luzerne (Medicago sativa)
Luzerne (Medicago sativa)Wild-, Heil- und Zierpflanzen Wicke (Vicia sp.)
Wundklee (Anthyllis sp.)
Hornklee (Lotus sp.)
Platterbse (Lathyrus sp.)
Goldregen (Laburnum sp.)
Ginster (Genista sp., Cytisus sp.)
Robinie (Robinia sp.)
Lupine (Lupinus sp.) Zaunwicke (Vicia sepium)
Zaunwicke (Vicia sepium) Gemeiner Goldregen (Laburnum anagyroides)
Gemeiner Goldregen (Laburnum anagyroides) Gewöhnliche Robinie (Robinia pseudoacacia)
Gewöhnliche Robinie (Robinia pseudoacacia) Lupine (Lupinus sp.)
Lupine (Lupinus sp.)und andere, auch viele tropisch verbreitete Gattungen.
Die bekanntesten Pflanzenarten, die mit Rhizobien in Symbiose leben, gehören zu den Schmetterlingsblütlern (Faboideae), aber auch innerhalb der mit diesen eng verwandten Johannisbrotgewächsen (Caesalpinioideae) und Mimosengewächsen (Mimosoideae) findet man Symbiosen mit Rhizobien. Wichtige Rhizobien-Arten sind Rhizobium leguminosarum, R. tropici, R. loti, R. trifolii, R. meliloti, R. fredii, Bradyrhizobium japonicum, B. elkanii, Azorhizobium caulinodans (tropisch, bildet Stängelknöllchen).
Die Symbiose zwischen Pflanzen und Stickstoff fixierenden Bakterien ist aber nicht auf Leguminosen und Rhizobien beschränkt. Enge Lebensgemeinschaften findet man zum Beispiel auch zwischen Erlen und Aktinomyzeten der Gattung Frankia oder dem tropischen Schwimmfarn Azolla und Cyanobakterien der Gattung Anabaena.
Ansammlungen stickstoffbindender Bakterien konnten auch in der Rhizosphäre verschiedener Gräser nachgewiesen werden. Kohlenhydrate, die die Pflanzenwurzeln absondern, dienen hier als Energiequelle.
Es existieren auch frei lebende Bakterien, die - ohne eine Symbiose einzugehen - N2 nur für den Eigenbedarf assimilieren (zum Beispiel Azotobacter und einige Cyanobakterien). Diese Art der Stickstoff-Assimilation, die „Stickstoff-Fixierung“, ist aufgrund des hohen Energiebedarfes jedoch streng reguliert und findet nur statt, wenn keine andere Stickstoffquelle verfügbar ist.
Literatur
- M. S. Woronin: Über die bei der Schwarzerle (Alnus glutinosa) und bei der gewöhnlichen Gartenlupine (Lupinus mutabilis) auftretenden Wurzelanschwellungen. Mémoires de l'Academie Impériale des Sciences de St. Pétersbourg, VII Series, vol. X. 1866.
- Wolfgang Böhm: Die Fixierung von elementarem Stickstoff durch die Wurzelknöllchen der Leguminosen. Zur Erinnerung an Hermann Hellriegels epochemachende Entdeckung im Jahre 1886. In: Angewandte Botanik Bd. 60, 1986, S. 1-5 (mit Bild).
- Lincoln Taiz, Eduardo Zeiger: Plant Physiology - das Original mit Übersetzungshilfen. 4. Auflage. Spektrum Akademischer Verlag / Springer-Verlag, Heidelberg u. a. O. 2007, ISBN 3-8274-1865-8.
- Gerhard Richter: Stoffwechselphysiologie der Pflanzen. 6.Auflage, Thieme Verlag, Stuttgart 1998, ISBN 3-13-442006-6.
- Hans W. Heldt und Birgit Piechulla: Pflanzenbiochemie. 4. Auflage, Spektrum Akademischer Verlag, Heidelberg 2008. ISBN 978-3-8274-1961-3; S. 295–309
- Bob B. Buchanan, Gruissem, Jones: Biochemistry& Molecular Biology of Plants 6. Auflage, American Society of Plant Physiologists, Rockville 2006, ISBN 0-943088-39-9.
Siehe auch
Weblinks
 Commons: Wurzelknöllchen – Sammlung von Bildern, Videos und Audiodateien
Commons: Wurzelknöllchen – Sammlung von Bildern, Videos und Audiodateien











Wikimedia Foundation.