- Webspinnen
-
Webspinnen 
Vierfleckkreuzspinne (Araneus quadratus)
Systematik ohne Rang: Urmünder (Protostomia) Überstamm: Häutungstiere (Ecdysozoa) Stamm: Gliederfüßer (Arthropoda) Unterstamm: Kieferklauenträger (Chelicerata) Klasse: Spinnentiere (Arachnida) Ordnung: Webspinnen Wissenschaftlicher Name Araneae Clerck, 1757 Die Webspinnen (Araneae) sind die bekannteste Ordnung der Spinnentiere (Arachnida), einer Klasse der Gliederfüßer (Arthropoda). Bis einschließlich Dezember 2010 wurden 42.055 Arten in 110 Familien beschrieben[1]. Sie teilen sich weiter in Gliederspinnen (Mesothelae), Vogelspinnenartige (Mygalomorphae) und Echte Webspinnen (Araneomorphae) auf. Die Webspinnen bilden damit nach den Milben (Acari) die artenreichste Ordnung der Spinnentiere.
Inhaltsverzeichnis
Bau der Webspinnen
 Unterseite einer Gartenkreuzspinne
Unterseite einer Gartenkreuzspinne
Webspinnen haben, wie alle Spinnentiere, acht Beine (im Unterschied zu den Insekten (Insecta), die nur sechs Beine haben). Kopf und Brust sind bei ihnen zu einem Stück, dem so genannten Prosoma (Cephalothorax) verschmolzen. Darauf folgt der ungegliederte, gestielte und meist deutlich größere Hinterleib, das Opisthosoma oder Abdomen.
Die vorderen Gliedmaßen des Kopfes, die großen Kieferklauen (Cheliceren), dienen zum Ergreifen der Beutetiere. Sie enden mit einer wie die Klinge eines Taschenmessers einklappbaren Klaue, in deren Spitze der Ausführungsgang einer Giftdrüse mündet. Das Gift fließt in die durch die Klaue geschlagene Wunde und tötet oder betäubt kleinere Beutetiere fast augenblicklich. Als zweite Kopfextremität folgen die Kiefertaster (Pedipalpen); diese sind mehrgliedrige Taster, beim Weibchen von der Form eines verkürzten Beins, beim Männchen mit aufgetriebenem, als Begattungsorgan dienendem Endglied.
Es folgen dann am Prosoma vier Paar Laufbeine. Die Beine bestehen aus je sechs oder sieben Gliedern und sind zusammen mit den Trichobothrien (Becherhaaren) ein wichtiges Sinnesorgan. Die bei den einzelnen Gattungen unterschiedlich langen und verschieden ausgeprägten Beinpaare enden bei den netzbauenden Trionycha mit zwei kammartig gezahnten Klauen (Tarsalklauen) und einer Mittelklaue am Tarsus, durch die der Faden meist mit den hinteren Beinen geführt wird. „Modernen“ Laufspinnen (Dionycha) fehlt diese Mittelklaue. An ihre Stelle treten etwa bei den Springspinnen Scopulahaare in Büscheln, die durch Adhäsionskräfte selbst auf senkrechten Glasscheiben den Beinen sicheren Halt für einen katapultartigen Absprung geben.
Der Hinterleib ist durch einen dünnen Stiel (Petiolus, gebildet aus dem ersten Hinterleibssegment) mit dem Prosoma verbunden. Er trägt keine Beine. An der Bauchseite des Opisthosomas liegt die Geschlechtsöffnung, und seitlich von dieser befinden sich die beiden Stigmen (Spaltöffnungen) der so genannten Lungensäckchen, öfters auch noch ein zweites Stigmenpaar. Den After umgeben am Ende des Hinterleibs vier oder sechs Spinnwarzen, aus denen die Absonderungen der Spinndrüsen hervortreten. Letztere sind birnenförmige, zylindrische oder gelappte Schläuche. Ihr proteinhaltiges Sekret gelangt durch Hunderte feiner Röhrchen nach außen, erhärtet an der Luft schnell zu einem Spinnfaden und wird unter Mithilfe der Fußklauen und manchmal der Spinnspulen zu einem Netz oder Gespinst verwebt.
Sinnesorgane
 Mildes Dornfingerspinne (Cheiracanthium mildei) Kopf und Vorderkörper mit den Punktaugen, Pedipalpus und Kieferklauen
Mildes Dornfingerspinne (Cheiracanthium mildei) Kopf und Vorderkörper mit den Punktaugen, Pedipalpus und KieferklauenDas Nervensystem besteht aus dem Gehirn und einer Brustganglienmasse.
Hinter dem Stirnrand stehen acht Punktaugen in einer nach den Gattungen und Arten verschiedenen Anordnung. Die Hauptaugen (das Paar der vorderen Mittelaugen, VMA) mit starrem Glaskörper befinden sich auf der Oberseite des Prosoma. Die Netzhaut der vorderen Mittelaugen ist durch ein bis sechs Muskeln seitlich verschiebbar, so dass das Gesichtsfeld erweitert wird. Eine Akkommodation findet nicht statt. Die VMA sind relativ gleichförmig gebaut. Sie verfügen wie etwa bei den Trichterspinnen über nur wenige bis hin zu vielen Sehzellen bei Springspinnen und Krabbenspinnen. Den sechsäugigen Spinnen (Dysderidae, Sicariidae, Oonopidae) fehlen die VMA.
Die drei Paar kleineren Nebenaugen (vordere Seitenaugen (VSA), hintere Seitenaugen (HSA) und hintere Mittelaugen (HMA)) sind wie bei den Säugetieren inverse Augen, die über ein Tapetum verfügen und daher das Licht reflektieren. Der Aufbau der Augen variiert bei den Familien. Springspinnen und Luchsspinnen haben kein Tapetum. Die Anatomie der Nebenaugen wurde daher auch von Homann zur systematischen Unterscheidung herangezogen.
Spinnen haben durch die Augenanordnung ein weites Gesichtsfeld. Trotzdem sind nur wenige Familien wie zum Beispiel die Springspinnen zum Formensehen befähigt. Wahrscheinlich können sie farbig sehen, und Ultraviolett-Rezeptoren sind, wie bei anderen Gliederfüßern auch, vorhanden.
Zu den Sinnesorganen gehört das lyriforme Organ zur Erkennung von Vibrationen. Das Organ findet sich an den Spinnenbeinen und besteht aus mehreren parallelen und unterschiedlich langen Spalten in der Exokutikula und sind meist in Gelenknähe platziert. Jeder Spalt endet in eine zylindrische Vertiefung, dem Kopplungszylinder, über den mittels eines Dendriten eine Sinneszelle gekoppelt wird. Spinnen können über das lyriforme Organ Netz- und Bodenvibrationen wahrnehmen und sie dienen wenigstens bei einigen Arten als Propriorezeptoren und somit der Orientierung.
Obwohl mit den lyraformen Organen auch Schall wahrgenommen wird, sind die Becherhaare (Trichobothrien) für die akustische Wahrnehmung wichtiger. Das Frequenzoptimum liegt bei 300 bis 700 Hz, bei Luftschwingungen zwischen 100 und 2.500 Hz. Die Vibrationen oder Schallkräfte verengen die Spalten und lösen so einen Reiz auf die Dendritenspitze aus. Schallwellen versetzen die Becherhaare in Schwingung, die auf dendritische Endigungen übertragen werden. Die Bewegungen des Haares werden mit Sinneszellen wahrgenommen, die auf drei verschiedene Auslenkungsrichtungen ansprechen.
Verdauung und Atmung
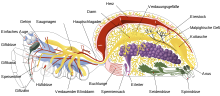 Innere Anatomie einer Webspinne
Innere Anatomie einer WebspinneDer Darmkanal läuft relativ geradlinig vom Mund zum After. Er ist in die Speiseröhre, den Magen mit fünf Paar Blindschläuchen und den Darm untergliedert. In den Darm münden die Lebergänge und zwei verästelte Harnkanäle. Der Lebersaft wirkt ähnlich dem der Bauchspeicheldrüse der höheren Wirbeltiere.
Die Atmungsorgane sind meist eigentümliche so genannte Fächertracheen oder Buchlungen, auch Lungensäckchen genannt. Aber es finden sich außerdem auch noch gewöhnliche Tracheen, in welche die Luft durch Luftlöcher (Stigmen) eintritt.
Das Herz ist ein pulsierendes, im Hinterleib gelegenes Rückengefäß (Arterie). Es besitzt seitliche Spaltöffnungen (Ostien), welche zum Eintritt des Bluts dienen. Bei Kontraktion des Herzens wird das Blut in die Arterie gepresst, während sich die seitlichen Spaltöffnungen wie Ventile schließen. Das Blut fließt durch Arterien zu den Gliedmaßen und zum Kopf, umspült zurückkehrend die Lungensäckchen und tritt durch die seitlichen Spaltöffnungen in das Rückengefäß zurück. Es handelt sich also um einen offenen Blutkreislauf.
Fortpflanzung und Entwicklung
 Zitterspinne (Pholcus sp.) mit Eikokon
Zitterspinne (Pholcus sp.) mit Eikokon geschlüpfte Jungspinnen
geschlüpfte Jungspinnen ein weiteres Spinnennest
ein weiteres SpinnennestDie Männchen, oft durch äußere Merkmale erkennbar und meist kleiner als die Weibchen, haben einen Hinterleib von geringerem Umfang als die Weibchen und besitzen paarige Hodenschläuche, aber in der Regel keine Penisstruktur, so dass mitunter so entfernt gelegene Gliedmaßen wie die Kiefertaster als sekundäre Begattungsorgane die Übertragung der Spermien auf das Weibchen übernehmen. Das verdickte Endglied der Kiefertaster ist dazu löffelförmig ausgehöhlt und enthält einen spiralig gebogenen Faden nebst hervorstreckbaren Anhängen. Bei der Begattung füllt das Männchen dieses Glied mit Spermien und führt es in die weibliche Geschlechtsöffnung ein, wo sich ein besonderes Behältnis zur Aufbewahrung der Spermien (die Samentasche) befindet. Die Weibchen besitzen unpaare oder paarige Eierstöcke, deren Eileiter meist gemeinschaftlich am Anfang des Hinterleibs ausmünden.
Bei Webspinnen ist das Verhalten des männlichen Tieres wichtig für eine erfolgreiche Befruchtung: Wenn das Männchen nicht das artspezifische Ritual einhält, kann es vom Weibchen nicht als Geschlechtspartner erkannt werden und wird nicht selten Opfer desselben (Kannibalismus). Alle Webspinnen legen Eier. Die Entwicklung im Ei ist insofern interessant, als der Embryo eine Zeit lang einen deutlich aus zehn bis zwölf Segmenten bestehenden Hinterleib besitzt, an dem sich auch die Anlagen von Gliedmaßen zeigen, die aber im weiteren Verlauf samt der Gliederung wieder verschwinden. Die Weibchen tragen die Jungen häufig bis zu ihrem Ausschlüpfen in einem Eikokon mit sich herum. Die ausschlüpfenden Jungen erfahren keine Metamorphose; haben also meist schon die Form der ausgewachsenen Tiere, bleiben aber bis nach der ersten Häutung im Gespinst der Eihüllen.
Junge Webspinnen verschiedener Arten erzeugen im Herbst lange Fäden (siehe Altweibersommer), mittels welcher sie sich hoch in die Luft erheben, um sich an andere Orte tragen zu lassen.
Die Lebensdauer der Webspinnen ist nicht wie bei den Insekten beschränkt; auch geschlechtsreife Tiere häuten sich in bestimmten Zeiträumen je nach Nahrungsangebot. Die Weibchen einiger Arten sind mehrjährig fortpflanzungsfähig. Manche können monatelang ohne Nahrung existieren. Ihre Regenerationsfähigkeit ist enorm. Verlorene Gliedmaßen können bei frühen Häutungen (nicht mehr bei der Reifehäutung) ersetzt werden.
Wachstum und Häutung
Wie bei allen Gliederfüßern ist das Körperwachstum durch das feste Exoskelett (Außenskelett) nur in sehr beschränktem Maße möglich. Bei Webspinnen kann sich das weichhäutige Abdomen ausdehnen. Extremitäten und Prosoma jedoch sind von einer harten Exocuticula umgeben, so dass sie nur durch eine Häutung wachsen können. Die Anzahl der bis zur Reife benötigten Häutungen ist abhängig von der Größe der Spinnen; kleinere Arten benötigen nur fünf, größere bis zu 10 Häutungen. Manche Vogelspinnen häuten sich auch noch nach der Geschlechtsreife. In den frühen Nymphenstadien ist die Zeit zwischen den Häutungen sehr kurz. Die Frequenz der Häutungen nimmt mit zunehmendem Alter ab und ist vom Nahrungsangebot (Schnelligkeit des Wachstums) abhängig. Tegenaria-Arten zum Beispiel verdoppeln etwa ihr Gewicht bei jeder Häutung.
Die Häutung kündigt sich durch den Rückzug der Spinne in ihren Schlupfwinkel und Nahrungsverweigerung an. Die Beine werden dunkler, und der zurückweichende Abdomen lässt den Petiolus sichtbar werden. Die neue Cuticula liegt dabei schon eingefaltet unter der alten. Haar und Borsten können komplett oder teilweise mit erneuert werden. Die meisten netzbauenden Spinnen lassen sich zur Häutung mit den Hinterbeinen am Häutungsfaden herunterhängen. Vogelspinnen legen sich auf den Rücken.
In der erste Phase klappt der Rückenschild wie ein Deckel ab. Hierbei beschleunigt sich der Herzschlag, und Hämolymphe wird durch das Herz, aber auch durch Kontraktionen der dorso-ventralen Hinterleibsmuskeln, in den Vorderkörper gepumpt, bis das Gewicht desselben sich verdoppelt hat und der Druck auf 200 mbar (150 mmHg) gestiegen ist. Zwar widersteht die Cuticula dem Fünffachen dieses Drucks, aber das Exoskelett wurde bereits durch den Abbau von innen geschwächt. Der Hinterleib schrumpft mit der alten Hülle dabei erheblich und sieht runzlig aus. Schließlich platzt die alte Hülle des Vorderkörpers an den seitlichen Rändern auf.
Danach wird der Hinterleib befreit. Die Risse des abgeplatzten Rückenpanzers erreichen auch den Hinterleib. Der Hinterleib kontrahiert sich wellenförmig und wird herausgezogen. Kurz zuvor heften die Spinnwarzen einen weiteren Faden an die alte Hülle an, so dass sich die Spinne aus der alten Haut abseilen kann.
Gleichzeitig mit dem Abseilen aus der alten Haut werden die Extremitäten herausgezogen. Dies ist der schwierigste Teil des Vorgangs, und hier können die schwerwiegendsten Komplikationen auftreten. Kann die Spinne ihre Beine nicht aus der Exuvie ziehen, stirbt sie. Nützlich ist dabei, dass die neue Cuticula noch nicht ausgehärtet ist und damit leichter durch Engstellen wie Tibia gleitet (besonders wichtig bei den männlichen Bulben, den Palpentarsi). Steigender Hämolymphdruck kann auch ein Ausquellen hervorrufen. Nach unten gerichtete Borsten verhindern ein Zurückgleiten der Beine. Bereits gehäutete Beine bilden ein Widerlager, um die restlichen Beine mit sehr großer Kraftanstrengung aus der alten Hülle zu ziehen. Die Reihenfolge kann artspezifisch sein.
Zum Schluss hängt eine frische Spinne schlaff an der alten Haut. Daraufhin beginnt sie mit einer Gymnastik, um während der Aushärtung die Gelenke beweglich zu halten. Der Häutungsvorgang kann von zehn Minuten bei kleinen Spinnen bis zu mehreren Stunden bei Vogelspinnen dauern. Nach der Gymnastik folgt häufig eine Körperreinigung, bei der die Laufbeine und Palpen durch die Cheliceren und Mundöffnung gezogen und aneinander gerieben werden.
Lebensweise der Webspinnen
 Eine Kreuzspinne spinnt ihre Beute ein
Eine Kreuzspinne spinnt ihre Beute einWebspinnen leben zumeist räuberisch und ernähren sich überwiegend von erbeuteten Gliedertieren, besonders Insekten, die sie aussaugen. Hierzu werden die Beutetiere zunächst mit einem enzymhaltigen Verdauungssaft aufgelöst, welchen die Spinne in ihr mit den Kieferklauen getötetes Opfer einbringt (siehe extraintestinale Verdauung). Viele Arten bauen Netze, um ihre Beute zu fangen.
Neben der aktiven Jagd gibt es zwei Ausnahmen in der Ernährungsweise. Erstens fressen nahezu alle Spinnen auch frische, tote Tiere und sind somit auch Aasfresser. Die zweite Ausnahme bildet die Springspinne Bagheera kiplingi, die sich fast ausschließlich vegetarisch von Belt'schen Körperchen einiger Akazien ernährt; dies sind spezielle Futterkörperchen direkt an den Blättchen, die diese Akazien für die beherbergten Ameisen insbesondere als Lock- und Nahrungsmittel ausbilden.
Die Gespinste der Webspinnen differieren je nach Gattung und Art im Aussehen. Oft halten sich die Tiere in der Nähe der Netze in röhren- oder trichterartigen Verstecken auf. Daneben gibt es auch viele Arten, die vagabundieren und ihre Beute im Lauf oder Sprung überwältigen.
Fast alle Webspinnen sind Landtiere. Sie sind über den ganzen Erdkreis verbreitet, doch finden sich in den heißeren Zonen die meisten und größten Arten. Die einzige Spinnenart, welche im Wasser lebt, ist die Wasserspinne (Argyroneta aquatica). Allerdings gibt es eine Reihe von Arten, die auf der Wasseroberfläche jagen.
Feinde
 Wegwespe mit erbeuteter Spinne
Wegwespe mit erbeuteter SpinneAls Fressfeinde von Spinnen spielen unter den Wirbeltieren vor allem Vögel eine wichtige Rolle. Amphibien, Reptilien (Geckos, Leguane, Salamander etc.) und Fledermäuse erbeuten seltener Spinnen. Weitere wichtige Fressfeinde sind andere Spinnen, so z. B. die Mimetidae, die sich ausschließlich von Spinnen ernähren. Manche Insekten wie zum Beispiel einige tropische Libellen und verschiedene Ameisen fressen Spinnen. Alle Wegwespen (Pompilidae) und einige Grabwespen fangen Spinnen für ihren Nachwuchs. Sie erbeuten eine Spinne, die sie mit ihrem Giftstachel betäuben, und bringen sie dann in ihr Nest ein, wo die Wespenlarve die Spinne auffrisst. Einige Schlupfwespen legen ihre Eier in lebende, häufig vorher betäubte Spinnenkörper, die sich entwickelnden Larven leben dann als Parasitoide in diesen Wirten. Auch die Kugelfliegen entwickeln sich auf diese Weise in Spinnen. Hinzu kommen verschiedene Parasiten wie etwa Fadenwürmer und Milben.
Systematik und Evolution der Webspinnen
Die weltweit rund 42.000 Arten der Webspinnen werden aktuell in 110 Familien aufgeteilt. In Mitteleuropa sind 43 Familien der echten Webspinnen und eine Familie der Vogelspinnenartigen, nämlich 3 Arten der Tapezierspinnen (Atypidae), heimisch.[2]
Anzahl Familien Anzahl Arten Belgien 38 701 Niederlande 40 621 Deutschland 38 1.004 Schweiz 41 945 Österreich 40 984 Tschechien 38 841 Schweden 38 906 Polen 37 792 Mitteleuropa gesamt 43 1.313 weltweit 110 42.473 Die systematische Einteilung erfolgt unter anderem aufgrund der Form und Größe der Spinndrüsen, der Anordnung der Augen, dem Bau der Cheliceren und der Pedipalpen sowie dem Vorhandensein eines Cribellum; in neuere Zeit aber immer häufiger aufgrund genetischer Analysen. Auf diese Weise ergeben sich drei als Unterordnungen eingestufte Gruppen:
Webspinnen (Araneae) Opisthothelae Vogelspinnenartige (Mygalomorphae, ca. 2500 Arten)
Echte Webspinnen (Araneomorphae, ca. 36.000 Arten)
Gliederspinnen (Mesothelae)
Die Gliederspinnen weisen als einzige rezente Webspinnen eine deutliche Segmentierung auf, ein urtümliches (plesiomorphes) Merkmal, das auch bei anderen Cheliceraten verbreitet ist. Die äußerliche Segmentierung spiegelt sich bei den Gliederspinnen auch in der inneren Organisation wider. Die Abdominalmuskulatur verbindet die Segmente jeweils zwischen den Sterniten und Tergiten und die dorso-ventrale Muskulatur verläuft von Tergit zu Sternit. Das Herz ist ebenfalls segmental gegliedert. Die Gliederspinne Liphistius besitzt vier Paare aktive Spinnwarzen, die sich aus den Segmenten zehn und elf (je Segment zwei Paar) auf der Ventralseite (Bauchseite) aus Extremitätenanlagen bilden; bei allen anderen Spinnen sind eine oder zwei Spinnwarzenpaare sekundär reduziert worden. Das vierte, innere Paar am zehnten Segment ist auch bei Liphistius funktionslos.
Die Segmentierung lässt sich bei den Vogelspinnenartigen und bei den Echten Webspinnen nur noch ansatzweise erkennen, zum Beispiel an Sterniten, Reliefierung und Musterung des Hinterleibes. Bei diesen Unterordnungen verschmolzen die sechs Extremitäten tragenden Segmente zum Prosoma (Vorderkörper); das siebte wurde zum Petiolus. Das sackförmige Opisthosoma (Hinterleib) der Webspinnen bildete sich aus den übrigen Segmenten, wie anhand ihrer Embryonalentwicklung zu sehen ist. Das zweite Segment trägt die Epigastralfurche (Geschlechtsöffnung). Zwischen dem zweiten und dritten Sternit befindet sich die Atemöffnungen zur Buchlunge.
 Die Rindenspringspinne Marpissa muscosa ist ein Vertreter moderner Spinnen
Die Rindenspringspinne Marpissa muscosa ist ein Vertreter moderner SpinnenDurch eine starke Streckung des dritten Sternits und eine Verkürzung der Tergite 13 bis 17 wanderten die Spinnwarzen nach hinten, wo sie heute bei den Mygalomorphen und Araneomorphen direkt unter der Kloake liegen. Zwischen dem dritten und vierten Sternit liegt direkt vor den Spinnwarzen, zusätzlich zu der nach außen gewanderten Atemöffnung der Buchlunge zwischen dem zweiten und dritten Hinterleibsternit, eine weitere Atemöffnung zum Röhren-Tracheensystem. Die Geschlechtsöffnung wanderte ebenso an eine günstigere Stelle auf der Bauchseite nach hinten. Beide Unterordnungen verfügen über ein Endoskelett, an dem die Saugmagenmuskulatur ansetzt.
Die Mygalomorphae besitzen meist drei Paar Spinnwarzen. Bei Angehörigen der Tapezierspinnen (Atypidae) sind drei Paar Spinnwarzen nur im Juvenilstadium aktiv, bei adulten Tieren bleibt das dritte Paar passiv. Bei anderen Vogelspinnenartigen sind nur noch die zwei Paare des elften Segmentes vorhanden. Die Echten Webspinnen besitzen in der Regel drei Paar Spinnwarzen. Das vierte Paar bildet sich bei ihnen teilweise zu weiteren Organen um. Bei den "Cribellaten" Spinnen bildete sich diese Paar zum Cribellum, auf denen die Spinnspulen im Ruhezustand eingeklappt sitzen. Entgegen der traditionellen Auffassung nimmt man heute in der Regel an, dass das Cribellum ein plesiomorphes Stamm-Merkmal ist und vielfach unabhängig voneinander verloren ging. Das homologe Organ bei einigen "Ecribellaten" ist der Colulus (ein Hügel mit unklarer Funktion, vermutlich funktionslos); bei anderen "Ecribellaten" fehlt dieses vierte Paar vollständig.
Ferner unterscheiden sich die Vogelspinnenartigen durch die Stellung der Kieferklauen; wegen dieser auffälligen Unterscheidung war dies früher namensgebend für die Unterordnungen. Bei den Echten Webspinnen arbeiten sie gegeneinander und sind auch als multifunktionales Werkzeug einsetzbar. Im Gegensatz dazu klappen die kräftigeren Cheliceren der Vogelspinnenartigen wie ein Taschenmesser auf das Kiefergrundglied.
Fossile Überlieferung
Die ältesten Fossilien von Araneae[3] stammen aus dem Karbon. Sie weisen wie die rezenten Mesothelae eine deutlich erkennbare opistosomale Segmentierung auf. Devonische, spinnenähnliche Fossilien, als Attercopus fimbriunguis beschrieben, werden nicht mehr als zu den eigentlichen Webspinnen gehörig betrachtet, sie werden heute in eigene Ordnung Uraraneida gestellt, die Wurzel- oder Schwestergruppe zu den eigentlichen Webspinnen sein könnte. Die Uraraneida besaßen bereits Spinnvermögen, aber noch keine Spinnwarzen[4]. Ebenso wie die heutigen Mesothelae besaßen sie keine Giftdrüse an den Chelicerenklauen. Den moderneren Unterordnungen Mygalomorphae und Araneomorphae sicher zuzuordnende Fossilien stammen erst aus dem Mesozoikum. Die Radiation der Spinnen scheint mit einer gewissen Verzögerung derjenigen der Insekten, ihrer wichtigsten Beutegruppe, zu folgen[5]. Die Spinnenfossilien der Trias, z.B. aus Südafrika und Nordamerika (Virginia) sind überwiegend bereits modernen Familien zuzuordnen. Am Ende der Kreide sind fast alle modernen Familien fossil nachgewiesen, rezente Formen weisen ihnen gegenüber kaum noch irgendwelche Veränderungen auf[6]. Die meisten und aussagekräftigsten fossilen Spinnen liegen als Einschlüsse in Bernstein vor. Neben dem bekannten baltischen Bernstein sind ältere Lagerstätten bedeutsam, so konnte z.B. die bisher nur wenig fossil dokumentierte Familie Linyphiidae durch Funde im libanesischem Bernstein auf die Kreide zurückdatiert werden[7]. Die Ursprünge der artenreichsten Familie, der Springspinnen, sind nach molekularbiologischen Analysen ebenfalls in die Kreide zu datieren, auch wenn hier noch keine Fossilien vorliegen[8]
 Spinnennetz
SpinnennetzSpinnen und Menschen
Spinnen als Ekeltier, Delikatesse oder Gottheit
In den Gesellschaften der westlichen Industrieländer herrscht eine irrationale Abneigung gegen diese Tiergruppe vor, die bis zur krankhaften Arachnophobie gehen kann, obwohl hier, mit Ausnahme von Australien, kaum humanpathogene Spinnen vorkommen. Unter den rund 40.000 Arten sind weniger als ein dutzend Arten für den Menschen gefährlich; wobei diese „Gefährlichkeit“ nur in den seltensten Fällen ernsthafte Folgen hat. Hingegen werden sie in anderen Gesellschaften, in deren Umwelt gefährliche Spinnen häufiger sind, toleriert, als Delikatesse verspeist oder gar als Gottheit verehrt.
In Kambodscha werden Vogelspinnen gesammelt. Ihnen werden die Kieferklauen herausgerissen, danach werden sie bei lebendigem Leibe frittiert und in den Städten frisch verkauft. In vielen Kulturen Asiens werden Spinnen in der Nähe des Menschen toleriert, da sie nützliche Insektenvertilger sind. In Westafrika wird die Spinne Anansi als hohe Gottheit verehrt. Anansi gilt hier als Urheber des Wissens und der Klugheit, Erfinder des Ackerbaus, Regen- und Wettergott.
In Japan gibt es die Sage um die Riesenspinne Tsuchigumo.
Gefährliche Spinnen
Unterschieden werden muss zwischen neural wirkenden (Neurotoxinen) und nekrotisch, also zellzersetzend wirkenden Giften. Das Gift der in Amerika und Afrika beheimateten Loxosceles-Arten wirkt zusätzlich hämolytisch. Dabei sind die meisten Gifte der Spinnen nicht zum Töten gedacht, sondern zum Betäuben, um die Beute lebend als Vorrat zu konservieren. Erst der Verzehr oder ein Tötungsbiss tötet die Beute.
Nur wenige in Mitteleuropa beheimatete Spinnen sind aufgrund ihrer Größe überhaupt in der Lage, die menschliche Haut zu durchdringen. Die durch die Öffnung der Cheliceren injizierte Dosis an Gift ist minimal, auch wenn die Gifte hochwirksam sind. Insgesamt ist die Wahrscheinlichkeit, von einer Spinne gebissen zu werden, verschwindend gering, denn Spinnen ergreifen zunächst die Flucht und stellen sich tot (Schreckstarre).
Die einzige auch im deutschsprachigen Raum heimische, obschon hier nur regional anzutreffende Spinnenart, deren Biss eine gewisse medizinische Relevanz hat, ist der Ammen-Dornfinger (Cheiracanthium punctorium). Sein Biss ist in erster Linie recht schmerzhaft. In seltenen Fällen wurde von Übelkeit, Kopfschmerzen und Erbrechen berichtet, seltener auch von Fieber und Schüttelfrost. Die Symptome klingen nach etwa drei Tagen ab. Die Bissstelle kann aber noch längere Zeit geschwollen oder gerötet sein. Die in Amerika gefürchtete und dort eingeschleppte Feldwinkelspinne (Tegenaria agrestis) bringt es immerhin auf leichte lokale Symptome wie Taubheitsgefühl, die nach kurzer Zeit wieder verschwinden - einzig im Falle einer Allergie gegen das Gift kann ein Biss mit weiteren Komplikationen einhergehen. Die Feldwinkelspinne ist auch in Mitteleuropa verbreitet, wo sich ihr Lebensraum allerdings überwiegend auf die freie und von Menschen eher gering frequentierte Natur beschränkt; innerhalb von Häusern ist sie nur selten anzutreffen, dies nicht zuletzt, da sie dort in direkter Konkurrenz stünde, mit den in Nordamerika wiederum fehlenden, in Mitteleuropa jedoch häufigen und relativ größeren, für den Menschen aber allesamt harmlosen, weiteren Arten der Gattung Tegenaria (so in etwa T. atrica, die Große Winkelspinne). Mit diesen Spinnen in Zusammenhang stehende Bissunfälle sind in Europa dementsprechend unbekannt.
Große Exemplare der Gartenkreuzspinne (Araneus diadematus) bringen es immerhin bis zu einem folgenlosen, manchmal schmerzhaften Zwicken, das nur in größter Not angewendet wird und sehr selten ist. Ähnlich wirkt das Gift der Wasserspinne (Argyroneta aquatica), die allerdings aufgrund ihrer Lebensweise mit dem Menschen kaum in Berührung kommen wird. Ein europäischer Vertreter der Schwarzen Witwen ist die in den Mittelmeerländern vorkommende Latrodectus tredecimguttatus. Die auch irreführend und falsch „Malmignatte“ genannte Spinne baute ihr Netz früher oft unter Toilettendeckeln einfacher Latrinen aufgrund des hohen Insektenaufkommens, das sich dort einstellte; bei der Latrinenbenutzung ist es gelegentlich zu Giftbissen gekommen.
Einige Vertreter der Gattung Echte Witwen (Latrodectus) wie beispielsweise die in Nordamerika beheimatete Schwarze Witwe (L. mactans) oder die australische Rotrückenspinne (L. hasselti) besitzen starke Gifte, so dass deren Biss für kranke Menschen, zum Beispiel für Allergiker, lebensgefährlich werden kann. Vom eigentlichen Biss einer Schwarzen Witwe ist zunächst nichts zu spüren, daher bleibt der Biss oft unbemerkt.
Selbst Bisse der in Australien in der Region um Sydney beheimateten Sydney-Trichternetzspinne (Atrax robustus) verlaufen meist glimpflich. Da diese Art ein ausgesprochener Kulturfolger ist, kommt es häufig zu Begegnungen mit ihr. Trotzdem wurden seit 1927 nur 13 Todesopfer gezählt.
Früher kam es bei der Einfuhr von Südfrüchten auch gelegentlich zu Bissunfällen durch Kammspinnen (Phoneutria spp.). Nachdem in den 1980er und 1990er Jahren kaum Einschleppereignisse dokumentiert werden konnten, sind diese zu Anfang des 21. Jahrhunderts wieder angestiegen. So wurde beispielsweise erst 2009 wieder eine mit einer Bananenkiste eingeschleppte Phoneutria boliviensis in Hessen entdeckt.[9] Phoneutria keyserlingi und Phoneutria nigriventer aus der Familie der südamerikanischen Kammspinnen besitzen das Neurotoxin mit der höchsten Letalität. Der lebensgefährliche Biss dieser zum Teil ungewöhnlich aggressiven Spinnen muss stets mit einem Antiserum behandelt werden.
Größtenteils ungefährlich sind die gemeinhin gefürchteten Vogelspinnen. Nur wenige Arten wie z. B. Poecilotheria spp. sind in der Lage, eine für gesunde Menschen stärkere Vergiftung mit Schüttelfrost, Lähmungen und Krämpfen auszulösen. Allergiker müssen sich allerdings auch vor harmloseren Vogelspinnen in Acht nehmen, deren Bisswirkung oft mit einem Bienenstich verglichen wird. Der primäre Schmerz selbst, welcher durch den reinen Biss entsteht, ist allerdings nicht zu unterschätzen, da die Beißwerkzeuge (Cheliceren) von großen Arten wie Theraphosa blondi eine Länge bis zu 2,5 cm erreichen können. Neuweltliche Arten der Unterfamilien Theraphosinae und Aviculariinae besitzen Brennhaare, die sie zum Teil auf Aggressoren schleudern können. Diese speziellen Haare, die mit Widerhaken versehen sind und sich in Augen und Schleimhäuten festsetzen können, werden nur bei neuweltlichen Gattungen wie beispielsweise Brachypelma, Theraphosa und Avicularia durch das sogenannten Bombardieren verteilt. Hierbei streift die Spinne mit ihrem letzten Beinpaar in schnellen ruckartigen Bewegungen über ihren Hinterleib (Opisthosoma) und „bürstet“ die leicht abbrechenden Haare dem potentiellen Angreifer entgegen (Bertani et al. 2003). Diese Brennhaare führen zu starken Reizungen und allergischen Reaktionen.
Siehe auch: Arachnologie, Spinne des Jahres
Literatur
- Peter Ax: Das System der Metazoa. Ein Lehrbuch der phylogenetischen Systematik. Bd 2. Gustav Fischer, Stuttgart 2001. ISBN 3-8274-1179-3
- Heiko Bellmann: Spinnen, beobachten – bestimmen. Naturbuch, Augsburg 1992. ISBN 3-894-40064-1
- Heiko Bellmann: Kosmos-Atlas der Spinnentiere Europas. Kosmos, Stuttgart 2006. ISBN 3-440-10746-9
- Richard C. Brusca, G. J. Brusca: Invertebrates. Kap. 19. Sinauer, Sunderland Mass 1990, S. 661 (2.Aufl.). ISBN 0-87893-097-3
- Rainer F. Foelix: Biologie der Spinnen. Thieme, Stuttgart 1992. ISBN 3-13-575802-8
- Stefan Heimer, W. Nentwig: Spinnen Mitteleuropas. Parey, Hamburg-Berlin 1991. ISBN 3-489-53534-0
- Dick Jones: Der Kosmos-Spinnenführer. Franckh-Kosmos, Stuttgart 1996. ISBN 3-440-06141-8
- Ernst Kullmann, Horst Stern: Leben am seidenen Faden. Franckh-Kosmos, Stuttgart 1981, 1996. ISBN 3-570-00597-6
- Peter Klaas: Vogelspinnen im Terrarium. Lebensweise, Haltung und Zucht. Ulmer, 2003. ISBN 3-8001-7933-4
- Gerhard Neuweiler, G. Heldmaier: Vergleichende Tierphysiologie. Bd 1. Neuro- und Sinnesphysiologie. Springer, Berlin 2003, S.164. ISBN 3-540-44283-9
- Rainar Nitzsche: Spinnen-Spiegelungen in Menschen-Augen. Nitzsche, Kaiserslautern 2004, 2005. ISBN 978-3-930304-65-3
- Franz Renner: Spinnen ungeheuer - sympathisch. Nitzsche, Kaiserslautern 1990, 2001. ISBN 978-3-980210201
- Michael J. Roberts: Field Guide. Spiders of Britain and Northern Europe. Harper Collins, London 1995. ISBN 0-00-219981-5
- Edward E. Ruppert, R. S. Fox, R. P. Barnes: Invertebrate Zoology - A functional evolutionary approach. Kap. 18. Brooks/Cole, Southbank 2004, S.571 (7.Aufl.). ISBN 0-03-025982-7
- Günter Schmidt: Giftige und gefährliche Spinnentiere. Westarp Wissenschaften, 2000. ISBN 3-89432-405-8
- Boris F. Striffler: Die Martinique-Baumvogelspinne. Avicularia versicolor. Natur und Tier Verlag, 2004. ISBN 3-937285-42-3
- Peter Weygoldt: Chelicerata - Spinnentiere. in: W. Westheide: Spezielle Zoologie. Teil 1. Einzeller und Wirbellose Tiere Gustav Fischer, Stuttgart-Jena 1996, 2004. ISBN 3-437-20515-3
Weblinks
 Commons: Webspinnen – Sammlung von Bildern, Videos und Audiodateien
Commons: Webspinnen – Sammlung von Bildern, Videos und Audiodateien Wiktionary: Webspinne – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen
Wiktionary: Webspinne – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen- Datenbank und Bestimmungsschlüssel der Spinnen Europas
- Norman I. Platnick, 2011: The World Spider Catalog, Version 12.0, American Museum of Natural History
- Checklist der Arachnologischen Gesellschaft e.V. - ARAGES
- Die Spinnen des Kaukasus (caucasus-spiders.info)
- Spiders of North-West Europe
- A proposal to standardise the ordinal names amongst the Arachnida
- Gifttier-Informationsdienst der Toxikologischen Abteilung Klinikum Rechts der Isar München
Quellen
- ↑ Norman I. Platnick, 2011. The World Spider Catalog, Version 11.5. American Museum of Natural History.
- ↑ Die Checklisten der Spinnentiere der AraGes
- ↑ ein Übersicht in: Paul A. Selden & DavidPenney (2010): Fossil spiders. Biological Revues 85: 171–206. doi:10.1111/j.1469-185X.2009.00099.x
- ↑ Paul A. Seldena, William A. Shear, Mark D. Sutton (2008): Fossil evidence for the origin of spider spinnerets, and a proposed arachnid order. Proceedings of the National Academy of Science USA vol. 105 no. 52: 20781-20785. doi:10.1073/pnas.0809174106
- ↑ D. Penney (2004): Does the fossil record of spiders track that of their principal prey, the insects? Transactions of the Royal Society of Edinburgh, Earth Sciences 94: 275–281.
- ↑ Jason A. Dunlop & Paul A. Selden (2009): Calibrating the chelicerate clock: a paleontological reply to Jeyaprakash and Hoy. Experimental & Applied Acarology 48: 183–197. doi:10.1007/s10493-009-9247-1
- ↑ David Penney & Paul A. Selden (2002): The oldest linyphiid spider in lower cretaceous lebanese amber. Journal of Arachnology 30: 487–493. doi:10.1636/0161-8202(2002)030[0487
- ↑ David E. Hill & David B. Richman (2009): The evolution of jumping spiders (Araneae: Salticidae): a review. Peckhamia 75(1): 1-7. PDF
- ↑ Peter Jäger, Theo Blick (2009): Zur Identifikation einer nach Deutschland eingeschleppten Kammspinnenart. Arachnologische Mitteilungen 38:33-36.(Online)


Dieser Artikel wurde am 19. April 2004 in dieser Version in die Liste der exzellenten Artikel aufgenommen. Kategorien:- Wikipedia:Exzellent
- Webspinnen











Wikimedia Foundation.