- Waldnashorn
-
Waldnashorn 
Schädel
Zeitraum Mittelpleistozän bis Jungpleistozän 600.000 bis 100.000 Jahre Fundorte - Süd-, Mittel- Osteuropa
- West-, Zentral-, Ostasien
Systematik Laurasiatheria Unpaarhufer (Perissodactyla) Nashörner (Rhinocerotidae) Waldnashorn Wissenschaftlicher Name Stephanorhinus kirchbergensis Jaeger, 1839 Das Waldnashorn (Stephanorhinus kirchbergensis), nach seinem Entdecker Carl Heinrich Merck auch Merck´sches Nashorn oder Mercknashorn genannt[1], war eine pleistozäne Nashornart im nördlichen Eurasien. Es lebte zeitgleich mit dem verwandten Steppennashorn (Stephanorhinus hemitoechus). Beide Nashornarten starben im Verlauf des Jungpleistozäns aus.
Inhaltsverzeichnis
Verbreitung und Lebensraum
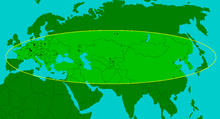 Verbreitungsgebiet des Stephanorhinus kirchbergensis im nördlichen Eurasien. Dargestellt ist die maximale Ausdehnung des Waldnashorns während der Warmzeiten. Die schwarzen Punkte stellen wichtige Fundplätze dar.
Verbreitungsgebiet des Stephanorhinus kirchbergensis im nördlichen Eurasien. Dargestellt ist die maximale Ausdehnung des Waldnashorns während der Warmzeiten. Die schwarzen Punkte stellen wichtige Fundplätze dar.
 Unterkiefer und Beckenknochen eines Waldnashorns, gefunden in Flurlingen bei Schaffhausen.
Unterkiefer und Beckenknochen eines Waldnashorns, gefunden in Flurlingen bei Schaffhausen.Das Waldnashorn war im Mittel- bis Jungpleistozän über weite Teile Eurasiens von Westeuropa bis Südsibirien, Zentralasien und China verbreitet. Sein östlichstes Auftreten ist von der Koreanischen Halbinsel bekannt.[2] Als wärmeliebendes Tier setzte es wenigstens sub-mediterranes Klima voraus und hatte sein Kerngebiet deshalb hauptsächlich im Mittelmeergebiet, in Vorder- und Zentralasien. Bemerkenswert ist allerdings, dass diese Tierart nicht auf der Iberischen Halbinsel vorkam.[3] In den pleistozänen Warmzeiten, in denen die Temperaturen im Klimaoptimum um teilweise bis zu 3° höher waren als heute, verbreitete sich das Waldnashorn auch weit bis in die nordeurasischen Gebiete. Im nordalpinen Europa erreichte es zur Holstein-Warmzeit (vor 340.000 bis 325.000 Jahren) mit England (Illford, Grays und Clacton-on-Sea) hier seinen nördlichsten Punkt. In der Eem-Warmzeit (vor 126.000 bis 115.000 Jahren) konnte es den Ärmelkanal allerdings nicht erneut überwinden und blieb auf das europäische Festland beschränkt.[4][3] Auch in seinem östlichen Verbreitungsgebiet dehnte das Waldnashorn während der Warmzeiten seinen Lebensraum von seinen Kerngebieten, wahrscheinlich Zentralasien und das nördliche China) weit nach Norden bis nach Sibirien aus. Schädel- oder Knochenreste dieser Nashornart wurden hier im Schwemmland von Don und Wolga in der russischen Steppe, ebenso wie im Kusnezker Becken[5] und bei Irkutsk[6] am Baikalsee gefunden. Der nördlichste Fund stammt aus dem Flussgebiet des Wiljui in der Republik Sacha in Jakutien und liegt nahe dem 64. nördlichen Breitengrad. Dieses hohe nördliche Vorkommen ist aber bisher singulär, da sich ansonsten die oberste Verbreitungsgrenze bei 53 bis 54 Grad nördlicher Breite befindet.[2]
Das Waldnashorn war ein typisches warmzeitliches Faunenelement. Seine Präsenz an Fundstätten wird als Maß für den Wechsel der Vegetationsgürtel im Verlauf von Kalt- und Warmzeiten genutzt.[7] Im westlichen Eurasien war es Bestandteil des sogenannten Elephas antiquus-Faunenkomplexes, dem neben dem namengebenden Europäischen Waldelefanten auch der Auerochse, der Rot- und Damhirsch, das Reh oder das Flusspferd angehören.[8] Ebenso war das Steppennashorn, mit dem es ko-existierte, ein typischer Vertreter dieser Faunengemeinschaft. Anders als sein gleichzeitig auftretender Verwandter kam das Waldnashorn aber nur während des Klimaoptimums vor. Bei Abkühlung zog es sich sehr rasch nach Süden in wärmerer Gefilde zurück. Da weder in Portugal noch in Spanien bisher Reste des Waldnashorns gefunden wurden, geht man davon aus, dass diese Region nicht zu seinem Kernrefugium gehörte. Vielmehr scheint es jedesmal vom nordalpinen Europa nach Asien oder Südosteuropa zurück gewandert zu sein.[3]
In Zentralasien gehörte das Waldnashorn zudem dem Koshkurgan-Faunenkomplex an, benannt nach einem mittelpleistozänen archäologischen Travertinfundplatz im Süden von Kasachstan, dem aber auch Funde aus Shoktas, ebenfalls Kasachstan, und Lachuti (Tadschikistan) zugerechnet werden. Hier war es mit dem Südelefanten, dem Mosbacher Pferd, dem Schoetensack-Bison und dem Elasmotherium vergesellschaftet.[9] In Südsibirien, so z. B. im Kusnezker Becken war das Waldnashorn Bestandteil des ebenfalls mittelpleistozänen Tatarischen Faunenkomplexes, zu dem wiederum der Rothirsch, der Riesenhirsch aber auch das Mammut zu zählen sind.[2] In Ostasien stellte diese Tierart - die hier lokal mit dem wissenschaftlichen Namen Stephanorhinus (Dicerorhinus) choukoutienensis bezeichnet wird[10] - einen Begleiter des Equus-Euctenoceros/Megaloceros-Faunenkomplexes (auch Nord-Tsingling-Faunenkomplex) dar, welcher seine Hauptverbreitung vor allem in Nordchina und in der Mongolei hatte. In diesen Komplex sind u. a. neben dem namengebenden Pferd und dem Riesenhirsch auch Elephas namadicus und einige nördliche Elemente, wie das Murmeltier oder der Moschusochse eingeschlossen. Gelegentlich kommen auch Nashörner der Gattung Coeledonta vor, die allerdings nicht zum typischen Wollnashorn gehören und deshalb auch nicht wie dieses als kaltklimatische Anzeiger gelten. Im hauptsächlich in Südchina und Indochina verbreitetem Stegodon-Ailuropoda-Faunenkomplex (auch Süd-Tsingling-Faunenkomplex genannt), benannt nach dem elefantenartigen Stegodon und dem Großen Pandabären, war das Waldnashorn bisher jedoch nicht nachgewiesen und sollte hier nach bisheriger Meinung durch das heute ebenfalls ausgestorbene Chinesische Nashorn (Rhinoceros sinensis) ersetzt worden sein. [11][12] Neuere Untersuchungen, u. a. aus der Nashorn-Höhle bei Shennongjia (Provinz Hubei), einem sehr fossilreichen Fundplatz, zeigen jedoch, dass das Waldnashorn gelegentlich doch in diesem Faunenkomplex auftritt und sich offensichtlich ökologische Nischen suchte, die seinen nördlichen Refugien entsprachen.[13] Wie weit er dabei nach Süden vorstieß, ist allerdings bisher nicht bekannt.
Fossilien dieser Art werden im Vergleich zu anderen pleistozänen Großsäugern relativ selten gefunden. Gründe dafür können entweder die schlechten Fossilisationsbedingungen sein oder die Art war tatsächlich relativ selten.[14][2] Generell ist sie im westlichen Eurasien häufiger nachgewiesen als im östlichen.[15] In Deutschland ist das Waldnashorn von mehr als einem Dutzend Fundstellen bekannt. Ein fast vollständiges Skelett wurde u. a. in den pleistozänen Deckschichten des Braunkohletagebaus von Neumark-Nord im Geiseltal (Sachsen-Anhalt) geborgen und gehört der Eem-Warmzeit an.[3] Außerdem wurde das Waldnashorn in warmzeitlichen Ablagerungen der wichtigen, bis zu 600.000 Jahre alten Fossilienfundstelle Mosbacher Sande im Stadtgebiet von Wiesbaden (Hessen) sowie aus dem etwa 300.000 Jahre alten frühmenschlichen Fundplatz Steinheim an der Murr (Baden-Württemberg) bekannt.[16] Weitere wichtige Fundstellen hier sind der rund 350.000 Jahre alte Homo erectus-Fundplatz Bilzingsleben (Thüringen),[17], der Fundort der etwa gleichalten Schöninger Speere (Niedersachsen)[18] und die auf ca. 220.000, möglicherweise auch nur auf 120.000 Jahre datierte Neandertaler-Station Ehringsdorf (ebenfalls in Thüringen).[19]
Körperbau und Ernährungsweise
 Schematische Darstellung der Kopfhaltung der drei im Mittel- und Jungpleistozän Europas vorkommenden Nashornarten,. Oben: Waldnashorn (Stephanorhinus kirchbergensis), Mitte: Steppennashorn (Stephanorhinus hemitoechus), unten: Wollnashorn (Coelodonta antiquitatis). Ohne Unterkiefer, Darstellung jeweils in linker Seitenansicht.
Schematische Darstellung der Kopfhaltung der drei im Mittel- und Jungpleistozän Europas vorkommenden Nashornarten,. Oben: Waldnashorn (Stephanorhinus kirchbergensis), Mitte: Steppennashorn (Stephanorhinus hemitoechus), unten: Wollnashorn (Coelodonta antiquitatis). Ohne Unterkiefer, Darstellung jeweils in linker Seitenansicht.Da im Gegensatz zum ebenfalls mittel- und jungpleistozänen Wollnashorn (Coelodonta antiquitatis) keine Höhlenzeichnungen oder gefrorenen Überreste vom Waldnashorn vorliegen, ist über sein Aussehen weit weniger bekannt als über das seines nördlichen Verwandten. Anatomische Untersuchungen zeigen, dass es einen kräftigen Körperbau und sehr lange und schmalen Vorder- und Hinterbeine hatte, deren Gelenke stark ausgeprägt waren. Vor allem die letzten beiden Merkmale zeigen, dass sich das Waldnashorn in Arealen bewegte, die eine eher geschlossene Vegetationsdecke aufwiesen.[10] Innerhalb der Stephanorhinus-Linie war das Waldnashorn deren größter Vertreter.[2] Rekonstruierte Körpergewichte schwanken zwischen 1,6 t[20] und maximal 2,9 t[21] und liegen damit in der Variationsbreite des Indischen Panzernashorns.
Der Kopf ist zwischen 70 und 80 cm lang und wirkt aufgrund der enormen Körpergröße relativ klein[3], was in der Vergangenheit häufig zur Diskussion über die tatsächliche Körpergröße geführt hat.[14] Markant am Schädel ist das verkürzte und dadurch eher rechtwinklige Hinterhaupt, welches bewirkte, dass das Waldnashorn seinen Kopf deutlich aufrecht hielt. Dadurch ähnelt es den früheren Stephanorhinus-Arten, weicht aber markant von seinem Verwandten, dem Steppennashorn ab, das den Kopf viel weiter nach unten hielt. In diesem Merkmal gleicht das Waldnashornauch auch dem heutigen Spitzmaulnashorn (Diceros bicornis) oder seinem nächsten heute noch lebenden Verwandten, dem Sumatranashorn (Dicerorhinus sumatrensis).[14][4] Wie die anderen Vertreter von Stephanorhinus trug es zwei Hörner auf der Nase und auf dem mittleren Schädel, deren Ansatzstellen an den Knochenoberflächen blumenkohlartig geraut sind. Das vordere Horn war aufgrund dieser gerauten Oberflächen größer und stieg steil nach oben.[3] Vermutlich war es aber nicht so groß wie beim Steppennashorn, worauf neben der geringeren Ausdehnung und Ausformung der Ansatzflächen beim Waldnashorn auch die Nasenscheidewand hinweist, die im Gegensatz zu seinem nahen Verwandten nur im vorderen Drittel verknöchert ist. Ein weiteres Merkmal ist der äußerst massiv gebaute Unterkiefer, der an Robustizität die seiner Verwandten weit übertrifft.[14]
Wie bei allen Stephanorhinus-Arten fehlten im Gebiss die Schneidezähne, weiterhin traten je Kiefernbogen drei Vorbackenzähne und drei Backenzähne auf. Von allen Arten der Stephanorhinus-Linie hat das Waldnashorn im Verhältnis zur Kopfgröße die größten Zähne, wobei die Dimensionen der Zähne von vorne nach hinten zunahm. Hier sticht besonders der dritte Molar hervor, der außerordentlich voluminös ausgebildet war. Dieses Merkmal weist darauf hin, dass diese Tierart vor allem große Mengen an pflanzlicher Nahrung zu sich genommen hatte, die aber insgesamt wenig Nährstoffe bot.[3] Die Zähne sind relativ hochkronig, wobei die Kronenhöhe von den Prämolaren zu den Molaren hin abnimmt, erreichen aber nicht die Werte des Steppennashorns.[14][4] Allerdings besitzen sie weniger Zahnzement, was charakteristisch ist für Tierarten, die hauptsächlich weiche Nahrung, wie Blätter, Blüten, Beeren oder Früchte verzehren. Darauf weisen zusätzlich auch die in der Mitte stets trogartig ausgekolkten Zahnoberflächen hin, was durch Abrieb beim Zerkauen der Nahrung entsteht.[22] Untersuchungen an Nahrungsresten, die an Zähnen vom Waldnashorn gefunden wurden, zeigen, dass sich diese Tierart zumindest in Mitteleuropa hauptsächlich von Pflanzenarten wie Birken, Rosen, Pappeln, Eichen, Weiß- und Feuerdorn aber auch von Seerosen und mitunter auch von Gräsern ernährte.[23][22]
Da es keine überlieferten Weichteile gibt, ist über das weitere Aussehen des Waldnashorns nichts bekannt. So ist auch unklar, ob dieses Nashorn ein Fell hatte, weiterhin fehlen Informationen über die Beschaffenheit der Haut, der eventuellen Ausbildung von Hautfalten oder die Größe und Form der Ohren.[24] Darüber hinaus gibt es auch über die Lebensweise dieser Tierart nur wenige Hinweise. Der Skelettbau und die Ernährungsweise lassen an einen Waldbewohner denken. Unklar ist dabei, ob das Waldnashorn tatsächlich in dichten Wäldern lebte oder eher im Übergangsbereich von den Waldrändern zu den offenen Landschaften.[14]
Stammesgeschichte
Das Waldnashorn war einer der letzten Vertreter der Gattung Stephanorhinus, als sein Zeitgenosse überlebte auch das Steppennashorn (Stephanorhinus hemitoechus) bis ins späte Pleistozän. Das mit ihm verwandte Sumatranashorn (Dicerorhinus sumatraensis), das ebenfalls zu den Dicerorhini gehört, hält sich bis heute in den Wäldern Südostasiens, obwohl es durch menschliche Einflüsse bis auf wenige hundert Tiere dezimiert wurde.
Über den Ursprung ist, ähnlich wie beim Steppennashorn wenig bekannt. Vermutlich stammt es von Stephanorhinus magarhinus ab und bildet mit ihm zusammen eine Schwesterklade der Linie Stephanorhinus etruscus-Stephanorhinus hundsheimensis-Stephanorhinus hemitoechus.[25][10] Wann die Tierart genau entstanden ist, ist bisher unklar. Auch hier wird ein Ursprung in Asien angeommen. In Europa erschien sie erstmals vor rund 600.000 Jahren, u. a. in den Mosbacher Sanden (Wiesbaden, Hessen; genauer Mosbach 2).[3] und Ponte Molle (Italien).[26] Ältere Fundstellen, wie z. B. Soleilhac (Frankreich) oder Tegelen (Niederlande], sind vermutlich Verwechslungen mit anderen Vertretern der Stephanorhinus-Linie.[14] In Ostasien trat das Waldnashorn sicher belegt vor rund 570.000 bis 580.000 Jahren an der berühmten Frühmenschen-Fundstelle Zhoukoudian (China, Lokalität 1) auf. Zwar wurde es auch von Gongwangling, einer mehr als 700.000 Jahre alten Fundstelle, beschrieben, doch sind die Überreste dort zu spärlich.[11][27] In Zentralasien lässt sich das Waldnashorn erst vor 500.000 bis 550.000 Jahren anhand der Fundstellen von Lachuti (Tadschikistan) und Koshkurgan (Kasachstan) nachweisen.[2][9]
Die frühesten Waldnashörner hatten dabei noch einen recht kleinen dritten Backenzahn, der erst im Laufe der Zeit an Größe zunahm. Im späten Mittel- und im frühen Jungpleistozän erreichte er dann seine größten Dimensionen. Diese Veränderungen am Gebiss werden häufig als biostratigraphische Anzeiger genommen.[28] Im Gegensatz zum Steppennashorn, nahm es aber nicht an Körpergröße zu, im Gegensatz entwickelten zahlreiche Populationen, vor allem in Südeuropa, kleinere Formen.[29]
In den kühleren Phasen der ausgehenden Eem-Warmzeit zog sich das Waldnashorn, wie bei den vorherigen Klimawechseln auch, aus den nördlichen Refugien nach Süden zurück. Nachweise der Tierart aus dieser späten Zeit stammen aus der Kůlna-Höhle im Mährischen Karst (Tschechien, Schicht 9b), aus der Veternica Höhle (Schicht j) und der Vindija-Höhle (Schicht K) (beide Kroatien) bzw. aus Grimaldi (Italien).[30][25] Doch schon kurz nach Beginn der letzten Vereisungsperiode (Weichsel-Kaltzeit) muss sie ausgestorben sein, da sie sich im weiteren Verlauf des Jungpleistozäns nicht mehr nachweisen lässt. Dasselbe Schicksal, zeitlich allerdings später, ereilte auch den Europäischen Waldelefanten und das Steppennashorn.[3][26] (vgl. Quartäre Aussterbewelle).
Forschungsgeschichte
Als fossile Art wurde das Waldnashorn erstmals 1839 durch den Stuttgarter Arzt und Paläontologen Georg Friedrich Jäger (1785–1866) wissenschaftlich beschrieben.[31] Für den von Jäger eingeführten wissenschaftlichen Namen Rhinoceros kirchbergensis lagen Zahnfunde von Kirchberg an der Jagst in Württemberg zugrunde. Der später genutzte Gattungsname Dicerorhinus wurde 1942 durch die vom ungarischen Paläontologen Miklós Kretzoi (1907–2005) eingeführte Bezeichnung Stephanorhinus ersetzt, Dies begründete er mit zahlreichen bestehenden anatomischen Unterschieden zum heute noch lebenden Sumatra-Nashorn als letzten Vertreter der Gattung Dicerorhinus.[32]
Im Lauf der Zeit wurde das Waldnashorn unter verschiedenen wissenschaftlichen Namen geführt:[2]
- Rhinoceros incisivus Merck 1784
- Rhinoceros megarhinus de Christol 1834 [33]
- Rhinoceros leptorhinus Cuvier 1836
- Rhinoceros kirchbergensis Jäger 1839
- Rhinoceros Merckii (merckii, mercki, merki, Mercki) Kaup 1841 [1]
- Dicerorhinus mercki Kaup 1841
- Rhinoceros leptorhinus Owen 1850 [34]
- Rhinoceros (Tichorhinus) Merckii Brandt 1877 [35]
- Rhinoceros Mercki (Merckii) var. Brachycephala Schroeder 1903 [36]
- Coelodonta merckii Abel 1919
- Dicerorhinus kirchbergensis Hooijer 1947 [37]
- Dicerorhinus mercki (kirchbergensis) (Jäger) var. Brachycephalus Schroeder
- vel Dicerorhinus merckii Mayer 1971 [38]
Literatur
- Paul S. Martin, Richard G. Klein (Hrsg.): Quaternary Extinctions. A Prehistoric Revolution. The University of Arizona Press, Tucson AZ 1984, ISBN 0-8165-1100-4.
- Arno Hermann Müller: Lehrbuch der Paläozoologie. Band 3: Vertebraten. Teil 3: Mammalia. 2. überarbeitete und erweiterte Auflage. Gustav Fischer Verlag, Jena 1989, ISBN 3-334-00223-3.
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Theiss-Verlag, Stuttgart 2002, ISBN 3-8062-1734-3.
Weblinks
 Commons: Stephanorhinus – Sammlung von Bildern, Videos und Audiodateien
Commons: Stephanorhinus – Sammlung von Bildern, Videos und AudiodateienEinzelnachweise
- ↑ a b Johann Jakob Kaup: Akten der Urwelt oder Osteologie der urweltlichen Säugethiere und Amphibien. Darmstadt, Verlag des Herausgebers, 1841
- ↑ a b c d e f g Emmanuel M. E. Billia: Revision of the fossil material attributed to Stephanorhinus kirchbergensis (Jäger 1839) (Mammalia, Rhinocerotidae) preserved in the museum collections of the Russian Federation. Quaternary International, Volume 179, Issue 1, March 2008, S. 25-37
- ↑ a b c d e f g h i Jan van der Made: The rhinos from the Middle Pleistocene of Neumark-Nord (Saxony-Anhalt). In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseums für Vorgeschichte 62. Halle/Saale 2010, S. 433–527
- ↑ a b c Mikael Fortelius, Paul Mazza und Benedetto Sala: Stephanorhinus (Mammalia: Rhinocerotidae) of the Western European Pleistocene, with a revision of S. etruscus (Falconer, 1868). Palaeontographia Italica, 80, 1993, S. 63–155
- ↑ Emmanuel M. E. Billia: First records of Stephanorhinus kirchbergensis (Jäger, 1839) (Mammalia, Rhinocerotidae) from the Kuznetsk Basin (Kemerovo region, Kuzbass area, South-East of Western Siberia). Bollettino della Società Paleontologica Italiana, 46 (2-3), 2007, S. 95-100
- ↑ Emmanuel M. E. Billia: The skull of Stephanorhinus kirchbergensis (Jaeger 1839) (Mammalia, Rhinocerotidae) from the Irkutsk region (Southwest Eastern Siberia). Quaternary International, Volume 179, Issue 1, March 2008, S. 20-24
- ↑ Donald R. Prothero,Robert M. Schoch: Horns, tusks, and flippers. The evolution of hoofed mammals. Johns Hopkins Univ Pr, Baltimore 2003, ISBN 0-8018-7135-2, S. 272.
- ↑ Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart, 2002, S. 153-155
- ↑ a b Anatoly P. Derevianko, Valery T. Petrin, Zhaken. K. Taimagambetov und Marcel Otte: Early Palaeolithic assemblages in trevertine, Southern Kazakhstan (a variant of an adaptation model. Anthropologie 36, 1998, S. 137-164
- ↑ a b c Frederic Lacombat: Phylogeny of the genus Stephanorhinus in the Plio-Pleistocene of Europe. Hallesches Jahrbuch für Geowissenschaften 23, 2007, S. 63–65
- ↑ a b Hans-Dietrich Kahlke: Zur chronologischen Stellung der Choukoutien-Kultur. Alt-Thüringen 6, 1963, S. 22-41
- ↑ Jean S. Aigner: Archaeological remains in Pleistocene China. AVA-Forschungen 1, Bonn, 1981
- ↑ Tong HaoWen und WU XianZhu: Stephanorhinus kirchbergensis (Rhinocerotidae, Mammalia) from the Rhino Cave in Shennongjia, Hubei. Chinese Science Bulletin 55 (4), 2010, S. 1157-1168
- ↑ a b c d e f g H.K. Loose: Pleistocene Rhinocerotidae of Western Europe with reference to the recent two-horned species of Africa and S.E. Asia. Proefschrift [Dissertation], Leiden, Scripta Geologica 33, 1975, S. 1–59
- ↑ Emmanuel M. E. Billia: Occurences of Stephanorhinus kirchbergensis (JÄGER, 1839) in Eurasia - An account. Acta Palaeontologica Romaniae 7, 2011,S. 17-40
- ↑ Karl-Dietrich Adam: Der Urmensch von Steinheim an de Murr und seine Umwelt. Ein Lebensbild aus der Zeit vor einer viertel Million Jahre. Jahrbuch des Römisch-Germanischen Zentralmuseums 35, 1988, S. 1–23
- ↑ Jan van der Made: A preliminary note on the rhinos from Bilzingsleben. Praehistoria Thuringica 4, 2000, S. 41–64
- ↑ Hartmut Thieme (Hrsg.): Die Schöninger Speere. Mensch und Jagd vor 400000 Jahren. Ausstellungskatalog. Stuttgart, 2007
- ↑ Walter Steiner: Der Travertin von Ehringsdorf und seine Fossilien. Lutherstadt Wittenberg 1981
- ↑ Mikael Fortelius (coordinator). Neogene of the Old World Database of Fossil Mammals (NOW). University of Helsinki, 2003 http://www.helsinki.fi/science/now/
- ↑ Andreas Wagner: Rekonstruktion von Körpermassen pleistozäner Rhinocerotidae in der Sammlung von Königswald. Frankfurt am Main, 2007
- ↑ a b Jan van der Made und René Grube: The rhinoceroses from Neumark-Nord and their nutrition. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 382–394
- ↑ René Grube: Pflanzliche Nahrungsreste der fossilen Elefanten und Nashörner aus dem Interglazial von Neumark-Nord. In: Jan Michal Burdukiewicz, Lutz Fiedler, Wolf-Dieter Heinrich, Antje Justus und Enrico Brühl (Hrsg.): Erkenntnisjäger. Festschrift für Dietrich Mania. Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle 57. Halle/Saale 2003, S. 221–236
- ↑ F. E. Zeuner: New reconstructions of the whooly rhinoceros and Merck's rhinoceros. Proceedings of the Linnean Society of London 156 (3), 1945, S. 183-195
- ↑ a b Claude Guérin: Les Rhinocerotidae (Mammalia, Perissodactyla) du Miocène terminal au Pléistocène supérieur d’Europe occidentale comparés aux espèces actuelles: tendances évolutives et relations phylogénétiques. Géobios 15, 1982, S. 599–605
- ↑ a b Emmanuel M. E. Billia und Carmelo Petronio: Selected records of Stephanorhinus kirchbergensis (Jäger, 1839) (Mammalia, Rhinocerotidae) in Italy. Bollettino della Società Paleontologica Italiana, 48, 2009, S. 1-12.
- ↑ Xinzhi Wu und Frank E. Poirier: Human evolution in China. New York, Oxford 1995
- ↑ Jan van der Made: Biostratigraphy - "Large mammals". In: Harald Meller (Hrsg.): Elefantenreich - Eine Fossilwelt in Europa. Halle/Saale, S. 83-92
- ↑ Frederic Lacombat: Pleistocene Rhinoceroses in Mediterranean Europe and Massif Central (France). Courier des Forschungs-Instituts Senckenberg 256, 2006, S. 57-69
- ↑ Rudolf Musil: The environmemt of the Middle Palaeolithic sites in Central and Eastern Europe. In: Jan Michał Burdukiewicz und Andrzej Wiśniewski (Hrsg.): Middle Palaeolithic human activity and palaeoecology: New discoveries and ideas. Studia Archeologiczne 41, Wrocław 2010, S. 121-180
- ↑ Georg Friedrich Jäger: Über die fossilen Säugetiere welche in Württemberg in verschiedenen Formationen aufgefunden worden sind, nebst geognostischen Bemerkungen über diese Formationen. C. Erhard Verlag, Stuttgart, 1835–39
- ↑ Miklós Kretzoi: Bemerkungen zum System der nachmiozänen Nashorn-Gattungen. Földtany Közlöny 72, 1942, S. 309-323
- ↑ Christol, J. de: Recherches sur les grandes espèces de rhinocéros fossiles. Montpellier, J. Martel,1834, 170 Seiten, 31 Abb.
- ↑ Owen, R.: A History of British fossil mammals and birds. London, Joh Van Voorst: IXLVII
- ↑ Brandt, J. F., Versuch einer Monographie der Tichorhinen Nashörner nebst Bemerkungen über Rhinoceros leptorhinus Cuv. u.s.w. Mém. Ac. Imp. Sc. St. Pétersb., Sér. VII, XXIV, 4, 1877
- ↑ H. Schroeder: Die Wirbelthierfauna des Mosbacher Sandes, I Gattung Rhinoceros. Abh. Kön. Preuss. Geol.L andesanst., N.F. 18, 1903, S. 1143
- ↑ Hooijer, D. Α.: Notes on some fossil mammals of the Netherlands. Arch. Mus. Teyler, 1947, Ser. 3, 10, 1
- ↑ Mayer, G.: Beiträge zur Geschichte der Badischen Landessammlungen für Naturkunde in Karlsruhe, III. Der Schädel des Dicerorhinus mercki (kirchbergensis) (Jaeger) var. brachycephalus Schroeder von Daxlanden und seine Geschichte. Beitr. naturk. Forsch. Südw. Dtl., 30/2, 1971, S. 157-163
Kategorien:- Unpaarhufer
- Ausgestorbenes Säugetier
- Fauna des Pleistozäns



Wikimedia Foundation.