- Stammesgeschichte des Menschen
-
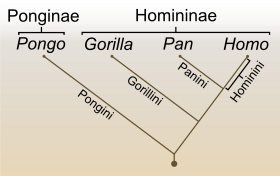 Kladogramm der Familie der Menschenaffen (Hominidae) und ihrer Unterfamilien Ponginae und Homininae sowie den heute noch lebenden Gattungen Pongo (Orang-Utans), Gorilla, Pan (Schimpansen) und Homo
Kladogramm der Familie der Menschenaffen (Hominidae) und ihrer Unterfamilien Ponginae und Homininae sowie den heute noch lebenden Gattungen Pongo (Orang-Utans), Gorilla, Pan (Schimpansen) und Homo
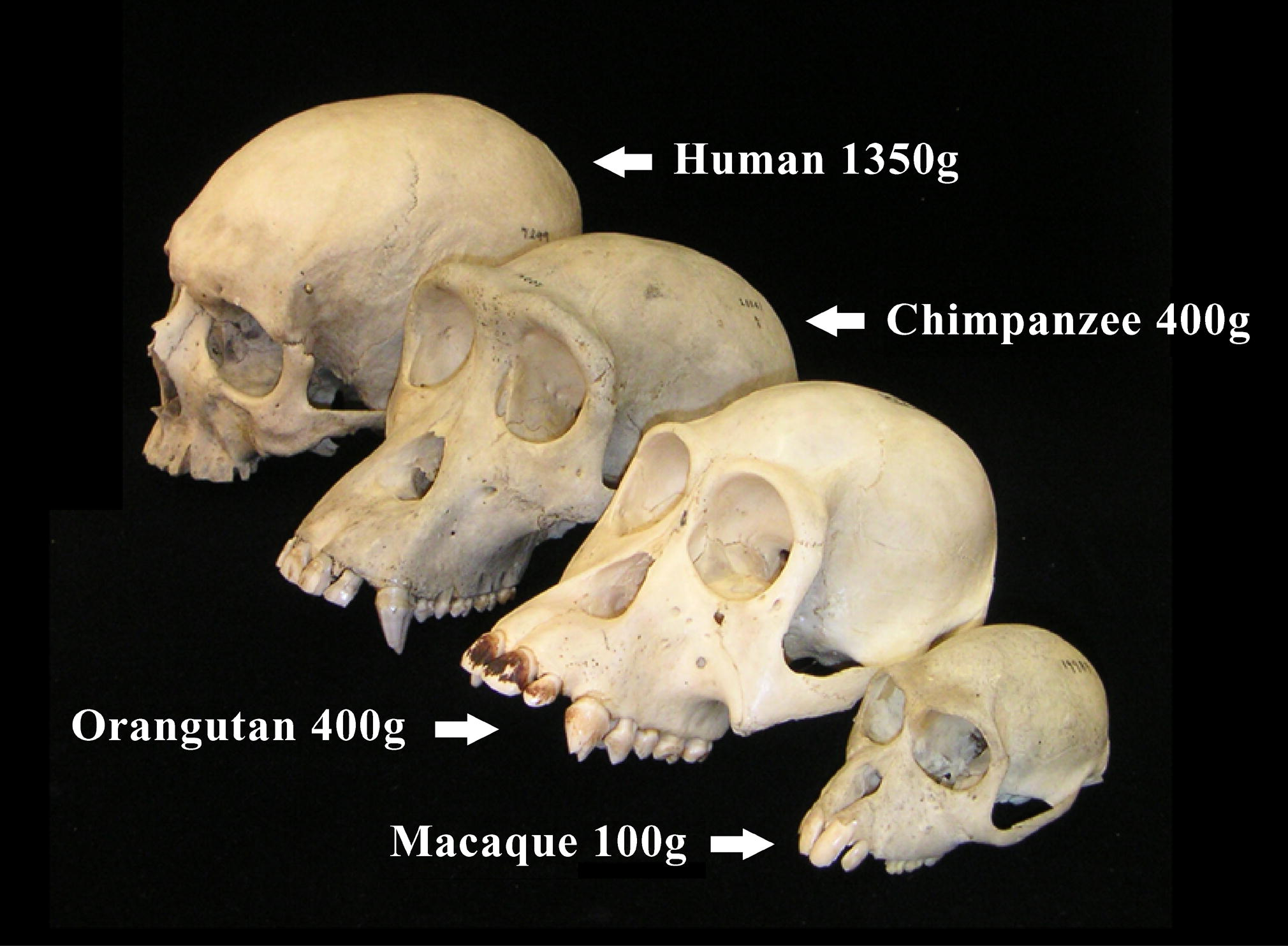
Als Stammesgeschichte des Menschen wird das durch Evolution bedingte Hervorgehen des modernen Menschen (Homo sapiens) und seiner nächsten Verwandten aus gemeinsamen Vorfahren bezeichnet. Die Stammesgeschichte des Menschen begann nach heutiger Auffassung mit der Aufspaltung der letzten gemeinsamen Vorfahrenpopulation der Schimpansen und des Menschen. Diejenige der beiden Teilpopulationen, aus der die Menschen hervorgingen, sowie alle ihre ausgestorbenen und noch lebenden Nachfahren werden als Hominini bezeichnet. Die Erforschung der Stammesgeschichte „zieht im Rahmen der von T. H. Huxley bereits 1863 begründeten Lehre von den Primaten, der Primatenkunde oder Primatologie, alle fossilen und heutigen Lebensformen der Affen in die Betrachtung ein“.[1]
Eine stetig wachsende Anzahl gut erhaltener Fossilien aus unterschiedlichen Regionen Afrikas, Asiens und Europas hat zwar die Erkenntnisse über die Vorfahren des Menschen erweitert, aber zu keiner Einigkeit über ihre Lebensdauer und Verbreitungsgebiete geführt. Dies ist auch dadurch bedingt, dass die systematische Zuordnung zahlreicher Einzelfunde und die Verwandtschaftsverhältnisse vieler vorzeitlicher Arten umstritten sind,[2] weil sich die Forschung derzeit noch „an den Grenzen oder sogar jenseits des analytischen Leistungsvermögens des Datenmaterials und der verfügbaren Methoden“ bewegt.[3]
Häufig werden die Arten der Hominini wie folgt bezeichnet: die Australopithecinen als Vormenschen; Homo habilis und Homo rudolfensis als Urmenschen; alle späteren Arten der Gattung Homo (außer Homo sapiens) als Frühmenschen; Homo sapiens als Jetztmensch oder moderner Mensch.[4] Den Vormenschen zeitlich vorangestellt wird gelegentlich – vor rund 20 Millionen Jahren, im oberen Miozän – ein sogenanntes Tier-Mensch-Übergangsfeld.
Inhaltsverzeichnis
Molekularbiologische Befunde zur Herkunft der Menschenartigen
Anhand einzelner Fossilienfunde kann die Existenz voneinander unterscheidbarer Arten belegt werden. Der genaue Zeitpunkt, ab dem sich der Körperbau der Individuen einer bestimmten Art durch Evolution so stark verändert hat, dass ihre Populationen einer zeitlich jüngeren Art zugeordnet werden können, lässt sich hingegen anhand einzelner Fossilien allenfalls grob abschätzen. Dies liegt daran, dass ein gleitender Übergang von den ursprünglichen Merkmalen zu den neu erworbenen Merkmalen zu erwarten ist. Eine scharfe Grenzziehung erweist sich hier als schwierig und wegen der Lückenhaftigkeit der fossilen Belege häufig als völlig unmöglich. Die sogenannte molekulare Uhr erwies sich als ein wichtiges Hilfsmittel, um die Zeitpunkte der Aufspaltung von Arten näherungsweise zu bestimmen, und zwar mit Hilfe von DNA-Analysen bei Individuen heute lebender Arten. Die Ergebnisse solcher Abschätzungen sind – nicht nur für die Frühphase der Primaten-Entwicklung – in Fachkreisen jedoch umstritten, denn die exakte Ganggeschwindigkeit der molekularen Uhr, das heißt: die Häufigkeit von Mutationen in vergangenen Epochen, ist unbekannt und ihre Kalibrierung ist auf Fossilienfunde angewiesen. So behielt die Ende der 1960er Jahre erstmals berechnete Zeitskala für die Evolution der Primaten[5] bis heute zwar grundsätzlich ihre Gültigkeit, erwies sich jedoch im Detail als zu gespreizt. Die anhand der molekularen Uhr errechneten Zeitpunkte und die durch Fossilienfunde erschlossenen Zeitpunkte weichen zudem häufig um mehrere Millionen Jahre voneinander ab.
So wurde 1985 aufgrund von DNA-Analysen eine Studie veröffentlicht, der zufolge vor rund 90 Millionen Jahren aus den Euarchontoglires jene Stammlinien hervorgingen, die einerseits unter anderem zu den heutigen Mäusen, andererseits zu den Menschenartigen, das heißt zu den Menschenaffen und Gibbons, führten.[6] Neuere Analysen bestätigen diese Datierung und geben für die Aufspaltung eine Zeitspanne von 80 bis 116 Millionen Jahre an.[7] Sie nennen ferner für die Abspaltung der zu den Meerkatzenverwandten (zu den Meerkatzen, Pavianen und Makaken) führenden Linie von der zu den Menschenartigen führenden Linie einen Zeitpunkt vor rund 23 Millionen Jahren, am Beginn des Miozäns.[8] Die Menschenartigen trennten sich dieser neueren Datierung zufolge vor rund 15 Millionen Jahren in die Gibbons und die Menschenaffen. Für den Zeitpunkt der Aufspaltung der Menschenaffen in die asiatischen Arten (die Vorfahren der Orang-Utans) und in die afrikanischen Arten wurden 11 Millionen Jahre errechnet, für die Abtrennung der Gorillas von den Schimpansen rund 6,5 Millionen Jahre und für die Abtrennung der Schimpansen von den Hominini schließlich 5,2 ± 1,1 Millionen Jahre.
Wie vorläufig diese Zeitangaben allerdings derzeit noch sind, zeigen folgende Beispiele: Während mit Hilfe der molekularen Uhr das Entstehen der Primaten in die Kreidezeit vor rund 90 Millionen Jahren datiert wird, wurde anhand von Fossilienfunden ein Zeitpunkt vor rund 56 Millionen Jahren – im Paläozän – berechnet.[7] Im Jahr 2007 wurden neu entdeckte, 10 Millionen Jahre alte Fossilien, genannt Chororapithecus, als Vorfahr der Gorillas gedeutet; sie sind somit mehr als drei Millionen Jahre älter als der Zeitpunkt, ab dem man – aufgrund der Schätzungen mit Hilfe der molekularen Uhr – Gorillas als eigenständige Gattung erwarten würde.[9] Auf der Basis von Fossilienfunden datierte Terry Harrison Anfang 2010 die Trennung der Schimpansen von den Hominini in die Zeit vor 7,5 Millionen Jahren,[10][11] C. Owen Lovejoy datierte diese Trennung 2009 hingegen in die Zeitspanne vor etwa 6 bis 5 Millionen Jahren.[12]
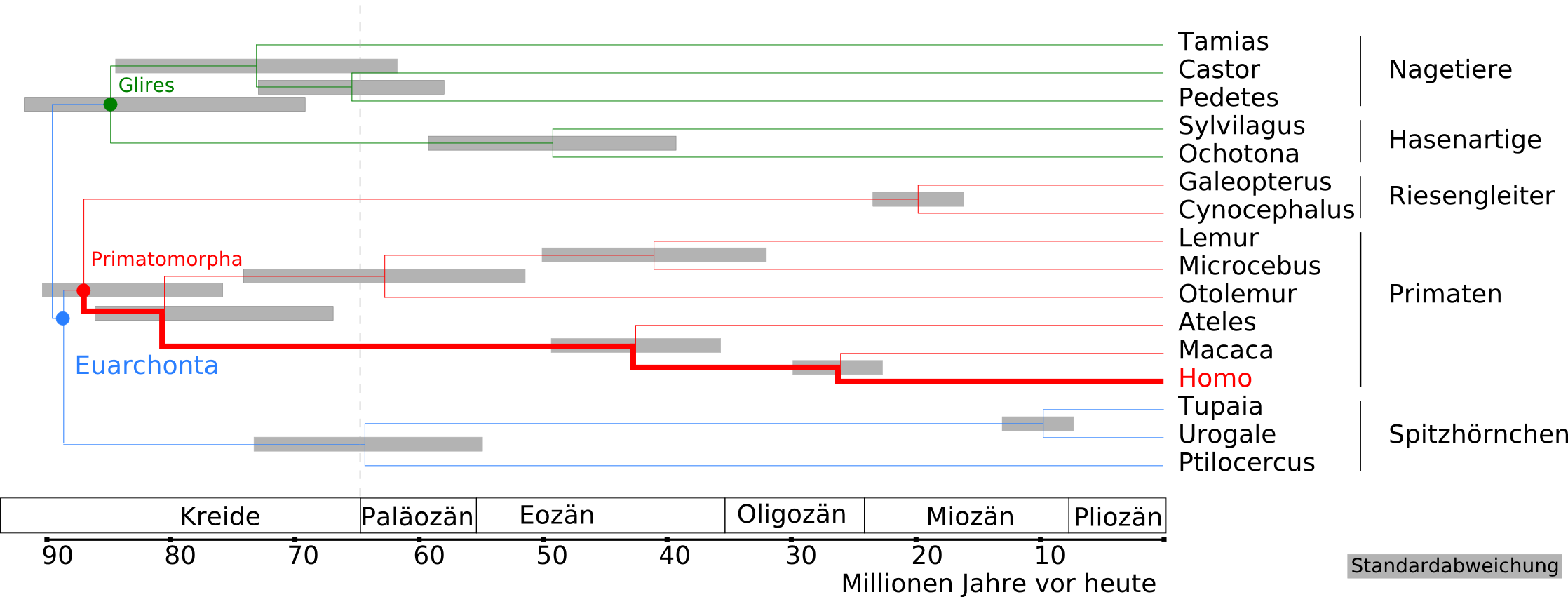
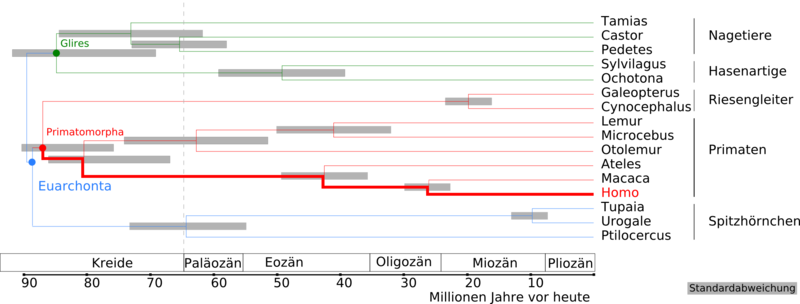 Kladogramm und Zeitspannen der Aufspaltung der Euarchontoglires in Euarchonta und Glires sowie in die Primatomorpha für folgende Taxa (nach Janečka u. a., 2007[13]):
Kladogramm und Zeitspannen der Aufspaltung der Euarchontoglires in Euarchonta und Glires sowie in die Primatomorpha für folgende Taxa (nach Janečka u. a., 2007[13]):
Nagetiere: Tamias (Streifenhörnchen) – Castor (Biber) – Pedetes (Springhasen)
Hasenartige: Sylvilagus (Baumwollschwanzkaninchen) – Ochotona (Pfeifhasen)
Riesengleiter: Galeopterus – Cynocephalus
Primaten: Lemuren – Microcebus (Mausmakis) – Otolemur (Riesengalagos) – Ateles (Klammeraffen) – Macaca (Makaken) – Homo
Spitzhörnchen: Tupaias – Urogale (Philippinen-Spitzhörnchen) – Ptilocercus (Federschwanz-Spitzhörnchen)Weiterentwicklung der Menschenartigen
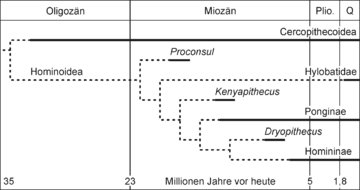 Die Ausdifferenzierung der Menschenartigen:[14] Um die Grenze vom Eozän zum Oligozän trennt sich im Formenkreis der Altweltaffen (Catarrhini) die zu den Cercopithecoidea führende Entwicklungslinie (hierzu gehören die Meerkatzenverwandten) von jener der Menschenartigen (Hominoidea). Zum Taxon der Menschenartigen gehören unter anderem die Gibbons (Hylobatidae), die Orang-Utans (Ponginae) und die afrikanischen Großen Menschenaffen (Homininae: Gorillas, Schimpansen, Mensch).
Die Ausdifferenzierung der Menschenartigen:[14] Um die Grenze vom Eozän zum Oligozän trennt sich im Formenkreis der Altweltaffen (Catarrhini) die zu den Cercopithecoidea führende Entwicklungslinie (hierzu gehören die Meerkatzenverwandten) von jener der Menschenartigen (Hominoidea). Zum Taxon der Menschenartigen gehören unter anderem die Gibbons (Hylobatidae), die Orang-Utans (Ponginae) und die afrikanischen Großen Menschenaffen (Homininae: Gorillas, Schimpansen, Mensch). Rekonstruktion des Körperbaus von Proconsul
Rekonstruktion des Körperbaus von Proconsul„Die Wiege der Hominoidea“ – der Überfamilie der Menschenartigen – „liegt im frühen Miozän Ostafrikas“,[15] das heißt in der Zeit vor 23 bis 16 Millionen Jahren.[10] Ihre frühmiozänen Formen werden als archaische oder Stamm-Hominoidea bezeichnet; ihre verwandtschaftlichen Beziehungen untereinander und zu späteren Arten sind wegen der geringen Zahl bekannter Fossilien umstritten. Ferner weist keine der bisher bekannten Arten von miozänen Menschenartigen die für Menschenaffen typischen körperlichen Merkmale des Fortbewegungsapparats auf, „die sich offenbar erst mit zunehmender terrestrischer Lebensweise pliozäner Hominoidea herausgebildet haben“,[16] das heißt vor rund 5 Millionen Jahren.
Vor rund 18 bis 15 Millionen Jahren trennten sich zwei Entwicklungslinien: Die eine führte zu den Gibbons (Hylobatidae), die andere zu einigen bereits im Miozän wieder ausgestorbenen Gattungen – wie Afropithecus, Kenyapithecus und Griphopithecus, Pierolapithecus, Dryopithecus und Oreopithecus – sowie zu den Menschenaffen,[10] das heißt zu den Orang-Utans (Ponginae) und zu den afrikanischen Großen Menschenaffen (Homininae: Gorillas, Schimpansen, Mensch).
Von einem Teil der Fachautoren wird heute angenommen, dass die Menschenaffen aus der in dieser Epoche existierenden Überfamilie der Proconsulartigen (Proconsuloidea) hervorgegangen sein könnten, für die Proconsul Namensgeber war[17] und die den paläontologischen Befunden zufolge „ein außergewöhnlich formenreiches Taxon [waren], das ausschließlich in tropischen Wäldern Afrikas und der Arabischen Halbinsel vorkam“.[10] Andere Fachautoren halten eine Schwestergruppe der Proconsulartigen für wahrscheinlicher oder verzichten völlig auf Spekulationen: „Nach gegenwärtigem Kenntnisstand ist jedoch keines der bekannten miozänen Hominoidea-Taxa in die direkte gemeinsame Vorfahrenschaft von afrikanischen Menschenaffen und der Stammlinie Hominini zu stellen.“[18]
Die Erstbesiedlung Eurasiens
Vor 17 bis 14 Millionen Jahren wurde das Klima in Afrika trockener, zugleich kam es im Jahresverlauf zu stärkeren Temperaturunterschieden. Diese Änderungen der ökologischen Gegebenheiten verringerten dem heutigen Forschungsstand zufolge die Proconsulartigen-Vielfalt; zugleich entwickelten sich frühe Verwandte der Meerkatzenverwandten und der Menschenartigen (beispielsweise Kenyapithecus wickeri, Equatorius africanus und Nacholapithecus) zu den vorherrschenden Gattungen. Nachweisbar ist bei den Fossilien aus dieser Epoche, dass Anpassungen an eine härtere Nahrung entwickelt wurden: u. a. kräftige Kiefer sowie dicke Zahnschmelz-Schichten auf den Backenzähnen[19] und dass vor 16 bis 15 Millionen Jahren erstmals auch Gebiete außerhalb Afrikas – weite Teile Eurasiens – von diesen Arten besiedelt wurden.[20]
Vor rund 16 Millionen Jahren, um die Grenze vom frühen zum mittleren Miozän, trennte sich in Asien die Verwandtschaftsgruppe der Orang-Utans von jener der anderen Menschenaffen; zur Verwandtschaftsgruppe der Orang-Utans zählen auch die fossil überlieferten Gattungen Ramapithecus, Sivapithecus / Ankarapithecus, Lufengpithecus und Khoratpithecus sowie Gigantopithecus.[10] Ramapithecus, dessen erste Fossilien Anfang der 1930er-Jahre in Nordindien gefunden worden waren, galt in den 1960er- und 1970er-Jahren irrtümlich als Vorfahre der Hominini.
 Der Unterkiefer von Dryopithecus fontani
Der Unterkiefer von Dryopithecus fontaniVor 13 bis 9 Millionen Jahren nahm die Zahl der Menschenartigen-Arten in Europa zu, u. a. sind aus dieser Epoche neben den beiden in Spanien entdeckten Funden von Pierolapithecus und Anoiapithecus mindestens vier Dryopithecus-Arten bekannt.[21] Eine vergleichbare Entwicklung vollzog sich in Asien in jenem Formenkreis, dessen einzige Überlebende die Orang-Utans sind.
Vor 9,6 Millionen Jahren führte die sogenannte Vallesium-Krise zu erheblichen Veränderungen der Ökosysteme in Europa:[22] Von der spanischen Mittelmeerregion ausgehend verschwanden in West- und Mitteleuropa die subtropischen, immergrünen Wälder infolge allmählicher Abkühlung; an ihrer Stelle folgten laubabwerfende Bäume und in einigen südlichen Regionen Steppen. Diese Klimaveränderung hatte auch einen tiefgreifenden Wandel in der Tierwelt zur Folge – die meisten europäischen Arten der Menschenartigen dieser Epoche starben aus; einzig Oreopithecus überlebte bis vor ungefähr 7 oder sogar 6 Millionen Jahren auf einer Insel in der Region von Sardinien / Korsika. Vor 8 bis 7 Millionen Jahren bewirkte die Anhebung des Tibetischen Hochlands eine Steigerung der Intensität des Monsuns in Asien. Die Folge war – auch in Europa – eine Verringerung der Niederschlagshäufigkeit, nachweisbar u. a. durch eine Ausbreitung von C4-Gräsern. Diese Klimaveränderungen führten zu einem völligen Aussterben der Menschenartigen in Europa[23] und zu einem Rückgang der Artenvielfalt in Asien, von dem letztlich nur die Orang-Utans und die Gibbons verschont blieben.
Die Entwicklung in Afrika
Betroffen von der Vallesium-Krise war vor 9,6 Millionen Jahren auch der Norden und Osten Afrikas.[24] Allerdings sind aus diesen Regionen bisher nur wenige Funde bekannt, die in die Zeit von vor 13 bis 7 Millionen Jahren datiert wurden. Dies hatte zeitweise zu der Vermutung geführt, dass die Menschenaffen sich in Asien entwickelt hatten und nach Afrika zurückgewandert waren.[25] Tatsächlich waren jedoch die schlechteren Voraussetzungen für das Entstehen von Fossilien – das feuchtere Klima – eine der Hauptursachen für die geringe Anzahl an Funden. Mit dem 2007 erstbeschriebenen, knapp 10 Millionen Jahre alten Nakalipithecus nakayamai aus Kenia,[26] dem schon länger aus Kenia bekannten, etwas älteren Samburupithecus kiptalami sowie dem in Äthiopien geborgenen, ebenfalls etwas älteren Chororapithecus kann inzwischen jedoch belegt werden, dass auch Afrika im mittleren und späten Miozän von einer Vielzahl von Menschenartigen-Arten bevölkert war. Im späten Miozän – vor 8 bis 6 Millionen Jahren – trennten sich in Afrika auch die Entwicklungslinien der Gorillas und der Schimpansen von jener der Hominini.[10]
Eine im Sommer 2006 in der Zeitschrift Nature veröffentlichte Genanalyse[27] des Broad Institute, eines gemeinsamen Instituts des Massachusetts Institute of Technology und der Harvard University, wurde dahingehend interpretiert, dass der Entwicklungsgang der frühesten Hominini ungewöhnlicher verlief, als zumeist angenommen wird. In dieser Studie waren 20 Millionen Basenpaare von menschlicher DNA sowie von Schimpansen- und Gorilla-DNA mit Hilfe der molekularen Uhr miteinander verglichen worden. Laut Studie separierte sich eine frühe Menschenaffenart bereits vor ca. 10 Millionen Jahren von den Vorfahren der Hominini. Diese beiden Populationen vereinigten sich jedoch einige Jahrtausende später wieder und bildeten eine Mischpopulation, in der es zu Kreuzungen kam.
 Das Fossil „Ardi“, eines Ardipithecus ramidus: Zeichnung nach der Rekonstruktionszeichnung in Science
Das Fossil „Ardi“, eines Ardipithecus ramidus: Zeichnung nach der Rekonstruktionszeichnung in ScienceDen Interpretationen der Forscher zufolge gab es über vier Millionen Jahre hinweg eine Abfolge von Kreuzungen und sich auseinander entwickelnden Gruppen, bis eine letztmalige, dauerhafte Trennung der Schimpansenvorfahren und der Hominini vor ca. 6,3 bis 5,4 Millionen Jahren erfolgte. Dieser letzte Genaustausch sei durch das durchgängig sehr geringe Alter der X-Chromosomen belegt, die sich erst zu diesem späten Zeitpunkt in der für Menschen charakteristischen Form herausbildeten und den X-Chromosomen des Schimpansen sehr ähnelten. Dieses Szenario blieb allerdings nicht unwidersprochen.
Die Auffächerung der Hominini
Die publizierten Daten zur Dauer der Existenz der Hominini-Gattungen und -Arten sind Schätzwerte, die abgeleitet wurden aus den geologischen (stratigraphischen) Untersuchungen der Fundstellen von Fossilien, das heißt: aus der Altersbestimmung einzelner Fundstellen. Die sogenannte molekulare Uhr kann für die Gattungen der Hominini nicht genutzt werden, da der moderne Mensch die einzige überlebende Art ist und nur von ihm und dem eng mit ihm verwandten Neandertaler genetisches Vergleichsmaterial vorhanden ist.
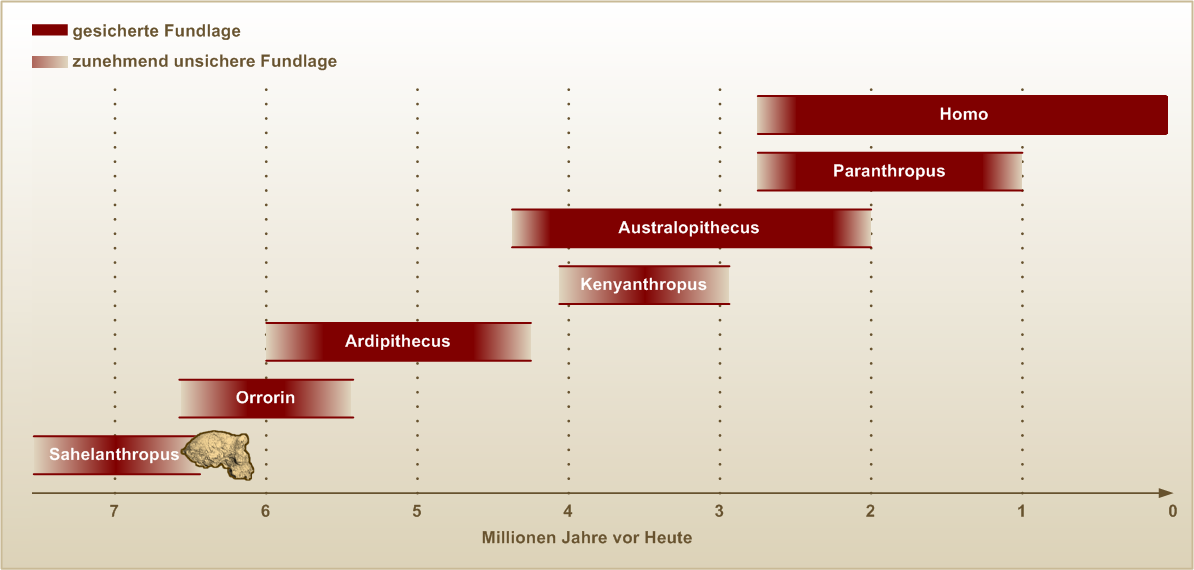
Völlig unklar ist aufgrund der wenigen Funde bislang, welche afrikanischen Vorfahren den rund 7 bis 5 Millionen Jahre alten Gattungen Sahelanthropus, Orrorin und Ardipithecus unmittelbar vorausgingen und ob diese drei Gattungen von ihren Entdeckern zu Recht in die direkte Ahnenreihe der Gattung Homo gestellt wurden.
Die Frühzeit der Hominini

Nach dem Fund des rund 6 Millionen Jahre alten Orrorin tugenensis im Jahr 2000 und des 6 bis 7 Millionen Jahre alten Sahelanthropus tchadensis im Jahr 2001 wurden beide Arten als bereits aufrecht gehende, älteste bisher bekannte Arten der Hominini ausgewiesen und somit als direkte Vorfahren des Menschen. Allerdings steht diese Deutung in Widerspruch zu den anhand der molekularen Uhr errechneten Befunden, denen zufolge die Trennung der zu Homo und zu den Schimpansen führenden Entwicklungslinien erst vor 5 bis 6 Millionen Jahren erfolgte.
Bereits 1994 waren in Äthiopien Fossilien von Ardipithecus ramidus entdeckt worden. Sie sind 4,4 Millionen Jahre alt und werden von vielen Forschern ebenfalls zu den direkten Vorfahren des Menschen gestellt. Auch die Individuen dieser Art konnten, wie vor allem das Fossil Ardi zu belegen scheint, auf dem Boden zweibeinig gehen. Die verwandtschaftlichen Beziehungen zwischen Sahelanthropus, Orrorin und Ardipithecus sowie die Beziehungen dieser drei Arten zu den späteren Hominini sind jedoch umstritten.
Diese Funde stammen aus dem Übergang von Messinium zum Zancleum, einer erdgeschichtlichen Epoche, in der die Weltmitteltemperatur ungefähr 4 °C wärmer war als heute.[28] Einer 2011 publizierten Studie zufolge waren die Lebensräume der frühen Hominini vor 6 Millionen Jahren Savannen mit allenfalls 40 Prozent Bedeckung durch Baumkronen; um 3,6 Mio. Jahre vor heute betrug die Baumbedeckung 40 bis 60 Prozent, und im Übergang vom Pliozän zum Pleistozän nahm sie wieder ab, so dass vor 1,9 Millionen Jahren Flächen mit einer Baumbedeckung von mehr als 50 Prozent nahezu verschwunden waren.[29]
Australopithecinen
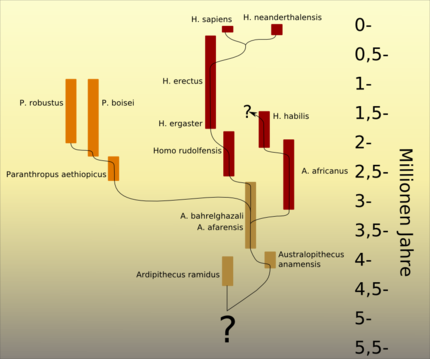 Hypothese zur Evolution der Australopithecinen, wie sie aufgrund der gegenwärtigen Fundlage beispielsweise durch Friedemann Schrenk vertreten wird.
Hypothese zur Evolution der Australopithecinen, wie sie aufgrund der gegenwärtigen Fundlage beispielsweise durch Friedemann Schrenk vertreten wird.Zu den durch Fossilienfunde relativ gut bekannten, bereits aufrecht gehenden Vorfahren des Menschen gehören die Australopithecinen, speziell die Gattung Australopithecus. In welchen verwandtschaftlichen Beziehungen ihre Arten zueinander und zu den Arten der Gattung Homo stehen, ist allerdings noch immer unklar. Australopithecus anamensis gilt heute als „die früheste unzweifelhafte Hominini-Art“;[10] eine der bedeutendsten Australopithecus-Fundstätten in Südafrika gilt daher als eine Wiege der Menschheit. Besonders bekannte Einzelfunde sind das Kind von Taung und das jugendliche, weibliche Skelett DIK 1-1 (beide gehören zu Australopithecus africanus) sowie Lucy, ein Australopithecus afarensis. Bei Laetoli in der Olduvai-Schlucht in Tansania wurden fossile Fußspuren von mehreren Australopithecus-afarensis-Individuen gefunden, die eindeutig belegen, dass Vertreter dieser Art aufrecht gingen. Umstritten ist hingegen, ob der 1999 von Meave Leakey in Kenia entdeckte Kenyanthropus platyops als eigenständige Gattung anzusehen oder zur Gattung Australopithecus zu stellen ist.
Ebenfalls zu den Australopithecinen wird die Gattung Paranthropus gestellt, deren Vertreter ein extrem robustes Gebiss aufweisen. Als Ursache dieser Anpassung gilt eine Klimaveränderung (Abkühlung) vor rund 2,5 Millionen Jahren; ausgelöst wurde sie zum einen durch die vor 2,7 Millionen Jahren beginnende Vergletscherung der Arktis, zum anderen durch die plattentektonische Hebung Ostafrikas. Beides bewirkte eine Verringerung der Niederschlagsmengen und in der Folge eine weitgehende Versteppung des angestammten Lebensraums der Vorfahren von Paranthropus.[30] Eine solche savannenartige Landschaft bot in erster Linie Nahrung für grasfressende Paarhufer und Wiederkäuer, die es vorher schon, meist in kleineren Formen, als laub-äsende Waldbewohner gab. Diese traten nun bald in großen Herden auf, und weil sie zahlreicher wurden, konnten sich auch Raubtiere und Aasfresser vermehren. So differenzierten sich zwei Typen von Hominini.
Der eine Typus entstand als Folge einer Anpassung an eine nunmehr – im Vergleich zum Laub der Wälder – hartfaserige Nahrung in der Savanne.[30] Paranthropus boisei, Paranthropus robustus und Paranthropus aethiopicus entwickelten in dieser ökologischen Nische eine gewaltige Kaumuskulatur und entsprechend mächtige Backenzähne. Ihre Kaumuskeln setzten an dem hohen Scheitelkamm des Schädels an.
Der zweite Typus fing die Folgen des Klimawandels ab, indem er zu einer Ernährungsweise überging, die mehr und mehr auch Fleisch als Nahrung einbezog.[30] Da diese Individuen weder die Fähigkeit besaßen, als Raubtier größere Beutetiere zu stellen noch über Klauen oder Zähne verfügten, die geeignet gewesen wären, ein großes Beutetier zu töten oder aufzubrechen, dürfte sich ihre Nahrung auf Aas und Beuteraub beschränkt haben. Sehr wahrscheinlich kam es hier zum ersten Gebrauch von Steinwerkzeugen, indem Steine dazu benutzt wurden, das Mark erbeuteter Röhrenknochen freizulegen.
Umstritten ist, welcher Gattung und welchen Arten diese ältesten Steinwerkzeuge zuzuordnen sind: „Von einigen Forschern wird die Ansicht vertreten, dass die Herstellung von Oldowan-Geräten dem Australopithecus zuzuschreiben sei und dass der Gebrauch von Werkzeugen nicht als ausschließlicher Anhaltspunkt für die menschliche Art angesehen werden dürfe. Es ist möglich, dass auch die Australopithecinen zu einer groben Steinbearbeitung fähig waren.“[31] Ein breites Formenspektrum von Steinwerkzeugen „und ihre systematische Herstellung mit Hilfe anderer Werkzeuge, also mit künstlich erschaffenen Geräten“ sei allerdings erst den Arten der Gattung Homo zuzuschreiben.
Die Gattung Homo
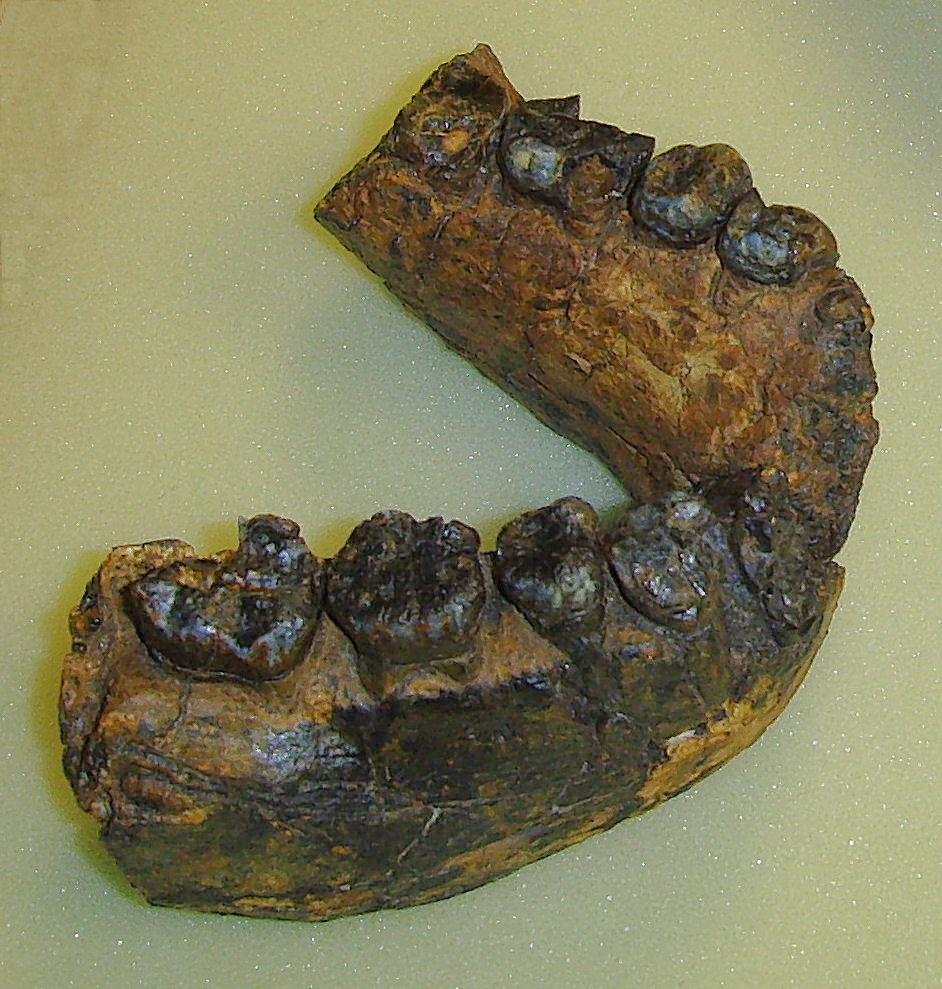
 Der Unterkiefer UR 501 von Homo rudolfensis ist mit 2,4 Millionen Jahren das älteste bekannte Fossil der Gattung Homo (Original, Sammlung Schrenk im Naturmuseum Senckenberg)
Der Unterkiefer UR 501 von Homo rudolfensis ist mit 2,4 Millionen Jahren das älteste bekannte Fossil der Gattung Homo (Original, Sammlung Schrenk im Naturmuseum Senckenberg)
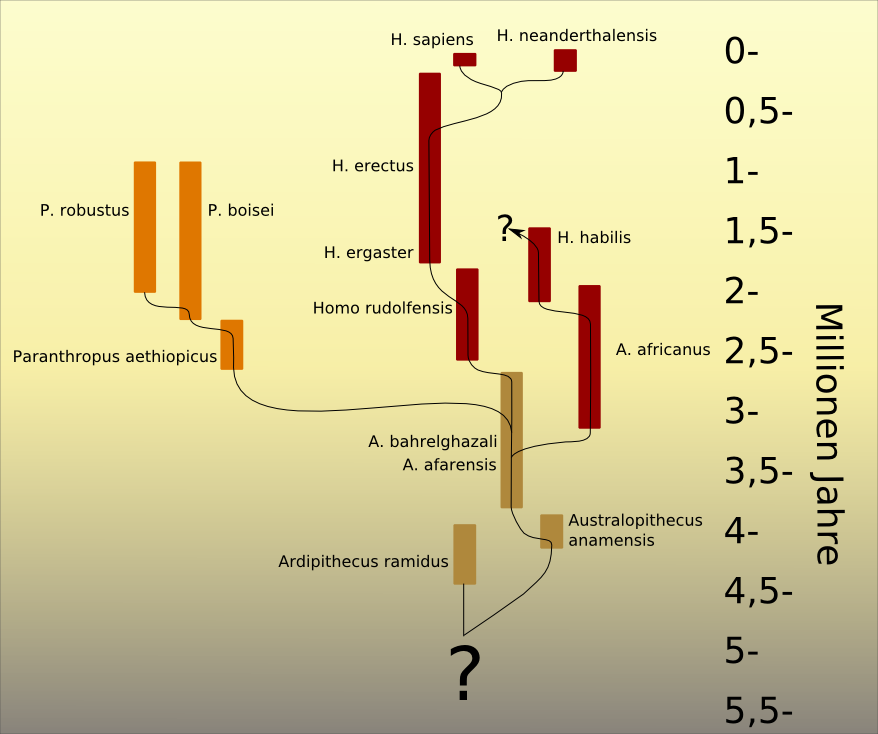
Aus einer Art der Gattung Australopithecus entwickelten sich vor drei bis zwei Millionen Jahren die ersten Vertreter der Gattung Homo, deren Fossilien insbesondere aufgrund von Werkzeugfunden zu Homo gestellt wurden.
Von Homo rudolfensis zu Homo erectus
Die beiden ältesten Homo-Arten sind Homo rudolfensis, der nach dem Rudolf-See – heute Turkana-See – in Kenia benannt wurde und Homo habilis. Die Verwandtschaftsbeziehungen beider Arten zueinander sowie ihre Verwandtschaft zu vorhergehenden und nachfolgenden Arten der Hominini sind bislang umstritten. Ihre noch sehr große anatomische Nähe zu Australopithecus veranlasste Bernard Wood 1999 sogar zu dem Vorschlag, beide Arten in Australopithecus rudolfensis und Australopithecus habilis umzubenennen.[32] Die anatomischen Merkmale der beiden Arten werden dennoch in der Regel in Abgrenzung von älteren Australopithecus-Arten definiert.
So ist das Gesicht von Homo habilis und Homo rudolfensis leichter gebaut als bei Australopithecus, aber die Augen stehen bei Homo habilis noch weiter auseinander als bei den späteren Homo-Arten wie beispielsweise Homo erectus. Der Überaugenwulst, das ist der durchgehende Querwulst des Stirnbeins oberhalb der Nasenwurzel, ist hingegen weniger stark ausgeprägt als bei Homo erectus. Der Schädel von Homo habilis und Homo rudolfensis „verengt sich hinter den Augenhöhlen nicht so stark wie bei Australopithecus oder Paranthropus, sodass sich ein größeres Gehirnvolumen ergibt“.[33]
Ober- und Unterkiefer beider Arten sind zudem kleiner als bei Australopithecus, entsprechend sind die Ansätze der Kaumuskulatur am Schädel weniger stark ausgeprägt.[34] Unterscheidungsmerkmale für Homo habilis und Homo rudolfensis sind laut Friedemann Schrenk: das bei Homo rudolfensis größere Gehirnvolumen; die oberen Vorbackenzähne von Homo rudolfensis haben 3 Wurzeln (Homo habilis: 2), die unteren 2 Wurzeln (Homo habilis: 1); die Weisheitszähne von Homo rudolfensis sind im Vergleich zu Australopithecus verkleinert (bei Homo habilis nicht verkleinert), Oberschenkel und Fuß von Homo rudolfensis sind menschenähnlich, bei Homo habilis ähneln sie Australopithecus.[35]
Vor etwa zwei Millionen Jahren entwickelte sich Homo ergaster. Sein Rang als abgrenzbare Art ist allerdings umstritten; diverse Forscher ordnen dessen Fossilien als frühe Exemplare der Art Homo erectus zu. Homo erectus war die erste Art der Gattung Homo, die Afrika verließ und sich über den Vorderen Orient nach Asien und Europa ausbreitete. Der erste Nachweis der Gattung Homo außerhalb Afrikas stammt allerdings von den homininen Fossilien von Dmanisi in Georgien, die auf 1,8 Millionen Jahre datiert wurden und deren Anbindung an den Stammbaum der Gattung Homo ungeklärt ist; möglicherweise gehören die Dmanisi-Fossilien zu Homo habilis.
Umstritten ist ferner, ob die von ihren Entdeckern als Homo antecessor bezeichneten, bis zu 1,2 Millionen Jahre alten spanischen Fossilien zu Recht als eigene Art ausgewiesen wurden oder als lokale Variante von Homo erectus einzuordnen sind.
Homo floresiensis, scherzhaft auch „Hobbit“ genannt, dessen Überreste 2003 entdeckt wurden, wird heute überwiegend als eine späte Zwergform des Homo erectus interpretiert. Angehörige dieser Art lebten noch bis vor 12.000 Jahren auf der indonesischen Insel Flores.
Von Homo erectus zum Neandertaler
Vor ca. 800.000 Jahren entwickelte sich aus Homo erectus eine Form mit größerem Gehirn, die als Homo heidelbergensis bezeichnet wird. Es ist umstritten, ob sie, wie in der angloamerikanischen Paläoanthropologie, als eigene Art oder, wie in der europäischen Paläoanthropologie, als eine Unterart des Homo erectus zu klassifizieren ist. Aus Homo heidelbergensis bzw. Homo erectus heidelbergensis entwickelten sich in Europa die Neandertaler (Homo neanderthalensis), während zur selben Zeit aus den in Afrika verbliebenen Populationen des Homo erectus der Homo sapiens hervorging, der heutige Mensch.
Neben dem Neandertaler gab es vor rund 40.000 Jahren zudem im Altai-Gebirge noch eine Schwestergruppe des Neandertalers, die so genannten Denisova-Menschen. Von ihnen ist bisher nur ein Backenzahn, ein Finger- und ein Zehenknochen aus der Denissowa-Höhle wissenschaftlich beschrieben worden.[36][37][38] Die Studien kamen aufgrund der Analyse von mtDNA und von Zellkern-DNA zu dem Ergebnis, dass die Denisova-Menschen zu einer homininen Gruppe gehören, die am engsten mit den Neandertalern verwandt ist, aber eine möglicherweise 250.000 Jahre lange eigenständige Populations-Geschichte neben den Neandertalern aufweist. Demnach hat es im Altai und neben Homo sapiens und dem Neandertaler noch eine dritte, unabhängig von diesen beiden Arten dorthin eingewanderte Population der Gattung Homo gegeben.
Die vier Arten der Gattung Homo – Homo erectus, Neandertaler, Homo floresiensis und Mensch – sowie die Denisova-Menschen besiedelten über gewisse Zeitspannen hinweg gleichzeitig Eurasien, denn noch aus der Zeit von vor ca. 50.000 Jahren wurden in abgelegeneren Gebieten, so auf Java, Fossilien des Homo erectus gefunden.
Ursprung des Homo sapiens
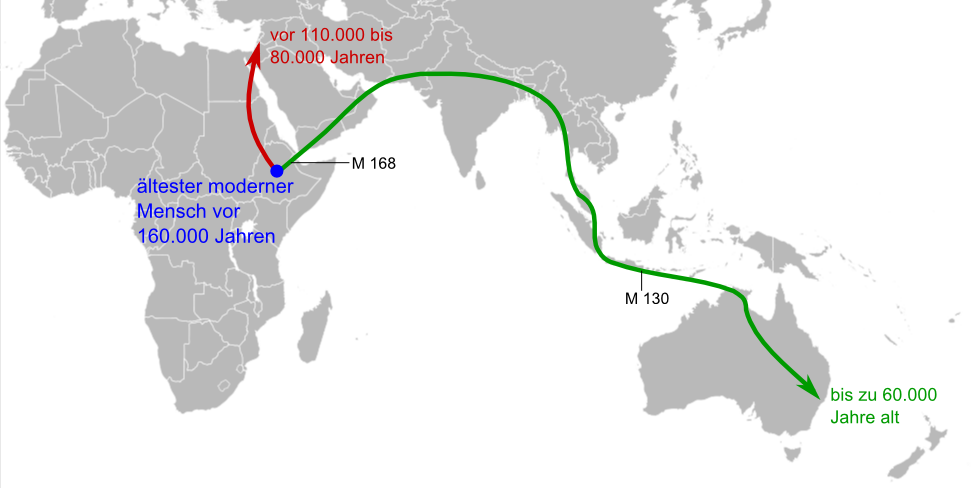
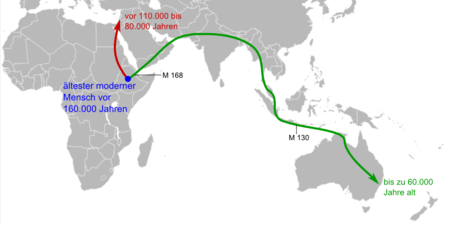 Die ersten Wanderungen des modernen Menschen außerhalb Afrikas gingen in den Nahen Osten und nach Australien. M 168 und M 130 bezeichnen genetische Marker im Y-Chromosom heute lebender Menschen.[39]
Die ersten Wanderungen des modernen Menschen außerhalb Afrikas gingen in den Nahen Osten und nach Australien. M 168 und M 130 bezeichnen genetische Marker im Y-Chromosom heute lebender Menschen.[39] Vier Nachkommen der ersten Auswanderungswelle des Homo sapiens:
Vier Nachkommen der ersten Auswanderungswelle des Homo sapiens:
Negrito-Frau (Philippinen); Aborigine-Mann (Australien);
Papua-Mann (Neuguinea); Melanesier-Junge (Vanuatu)Über die Ursprünge des Homo sapiens gingen die Meinungen lange Zeit auseinander. Von den Paläoanthropologen weitgehend akzeptiert ist heute die auf Fossilfunden basierende Out-of-Africa-Theorie. Ihr zufolge entstand der archaische Homo sapiens in der Zeitspanne zwischen 200.000 und 100.000 Jahren vor heute[40] in Afrika; die Ausbreitung des modernen Menschen in die anderen Regionen der Erde begann vor etwa 60.000 bis 70.000 Jahren über den Nahen Osten, der damals ökologisch zu Afrika gehörte. Vertreter der Hypothese vom multiregionalen Ursprung des modernen Menschen vertreten hingegen die Ansicht, dass sich lokale Populationen des Homo erectus, die viel früher aus Afrika auswanderten, mehrfach unabhängig voneinander in verschiedenen Regionen der Welt zum modernen Menschen entwickelten. Genetische Analysen des Y-Chromosoms und der Mitochondrien des Menschen stützen inzwischen die Out-of-Africa-Theorie (siehe dazu auch: Adam des Y-Chromosoms und Mitochondriale Eva). Eine Vermischung zwischen Homo sapiens und den späten Vertretern von Homo erectus in Asien ist unbelegt, die Vermischung zwischen Homo sapiens und dem Neandertaler war mit allenfalls 1 bis 4 Prozent eher gering und schlug sich nach den bisherigen Analysen nicht in äußerlich sichtbaren Merkmalen nieder.
Die ältesten bislang bekannten Funde des modernen Menschen in Afrika sind rund 195.000 Jahre (Omo 1 und Omo 2) bzw. etwa 160.000 Jahre alt („Homo sapiens idaltu“) und stammen aus dem Nordosten des Kontinents; ob Homo sapiens auch in dieser Region entstand oder ob sein Ursprung im südlichen Afrika liegt, ist derzeit Gegenstand der wissenschaftlichen Diskussion.[41] Einige Nukleotidsequenz-Varianten (Haplotypen) bei den afrikanischen Ethnien San, Mandinka und Aka wurden dahingehend interpretiert, dass es bei ihnen vor rund 35.000 Jahren zu einer „Vermischung“ mit einer Homo-Population gekommen sein könnte, die sich vor rund 700.000 Jahren von der zum modernen Menschen führenden Entwicklungslinie getrennt hatte;[42] wahrscheinlichster Kandidat hierfür sei – abgeleitet von den bislang bekannten Fossilien – Homo erectus.
Homo sapiens ist die einzige Menschenart, die Australien (vor etwa 60.000 Jahren) und Amerika besiedelt hat (vor etwa 11.500 bis 15.000 Jahren, nach mancher Ansicht wesentlich früher). Homo sapiens ist zugleich der letzte Überlebende der Gattung Homo.
Forschungsgeschichte
Unter Bezug auf Plinius hatte Andreas Vesalius (1514–1564), der Begründer der neuzeitlichen Anatomie und des morphologischen Denkens, bereits im 16. Jahrhundert eine Theorie der Abstammung des Menschen vom Affen über Pygmäen entwickelt, und der englische Anatom Edward Tyson hatte 1699 die Gemeinsamkeiten von Schimpansen- und Menschengehirn beschrieben[43]. Offenbar war man sich schon im 17. Jahrhundert in gelehrten Kreisen der großen Ähnlichkeit von Mensch und Menschenaffe bewusst. Funde menschlicher Fossilien im 18. und frühen 19. erwiesen sich jedoch entweder als Irrtümer, wie ein als Homo diluvii testis[44] bezeichneter tertiärer Riesensalamander, oder ihr „diluvialer“ Befundzusammenhang wurde nicht anerkannt.
Erst mit der Entdeckung des Neandertalers im Jahre 1856 – drei Jahre vor dem Erscheinen von Darwins Hauptwerk über Die Entstehung der Arten – waren Indizien dafür zutage getreten, dass es neben dem Homo sapiens noch einen weiteren Menschentyp gegeben haben könnte. Steinbrucharbeiter hatten in einem heute dem Kalkabbau zum Opfer gefallenen Abschnitt des Neandertals 16 Knochenfragmente freigelegt. 1864 schrieb sie der irische Geologe William King einer vom modernen Menschen unterscheidbaren Art zu, dem „Homo Neanderthalensis King“.[45] Gleichwohl blieb es bis zum Ende des 19. Jahrhunderts umstritten, ob der Fund aus dem Neandertal ein „Urmensch“ oder ein deformierter moderner Mensch sei.
Unabhängig von diesem Forscherstreit hatte Charles Darwin 1871 in seinem Werk Die Abstammung des Menschen und die geschlechtliche Zuchtwahl argumentiert, der Mensch habe sich vermutlich in Afrika entwickelt, da seine nächsten Verwandten – Schimpansen und Gorillas – dort beheimatet sind.[46] Auch die genauen anatomischen Studien, die Thomas Henry Huxley 1863 in seiner Schrift Evidence as to Man’s Place in Nature vorgelegt hatte, legten das Entstehen des Menschen in Afrika nahe.
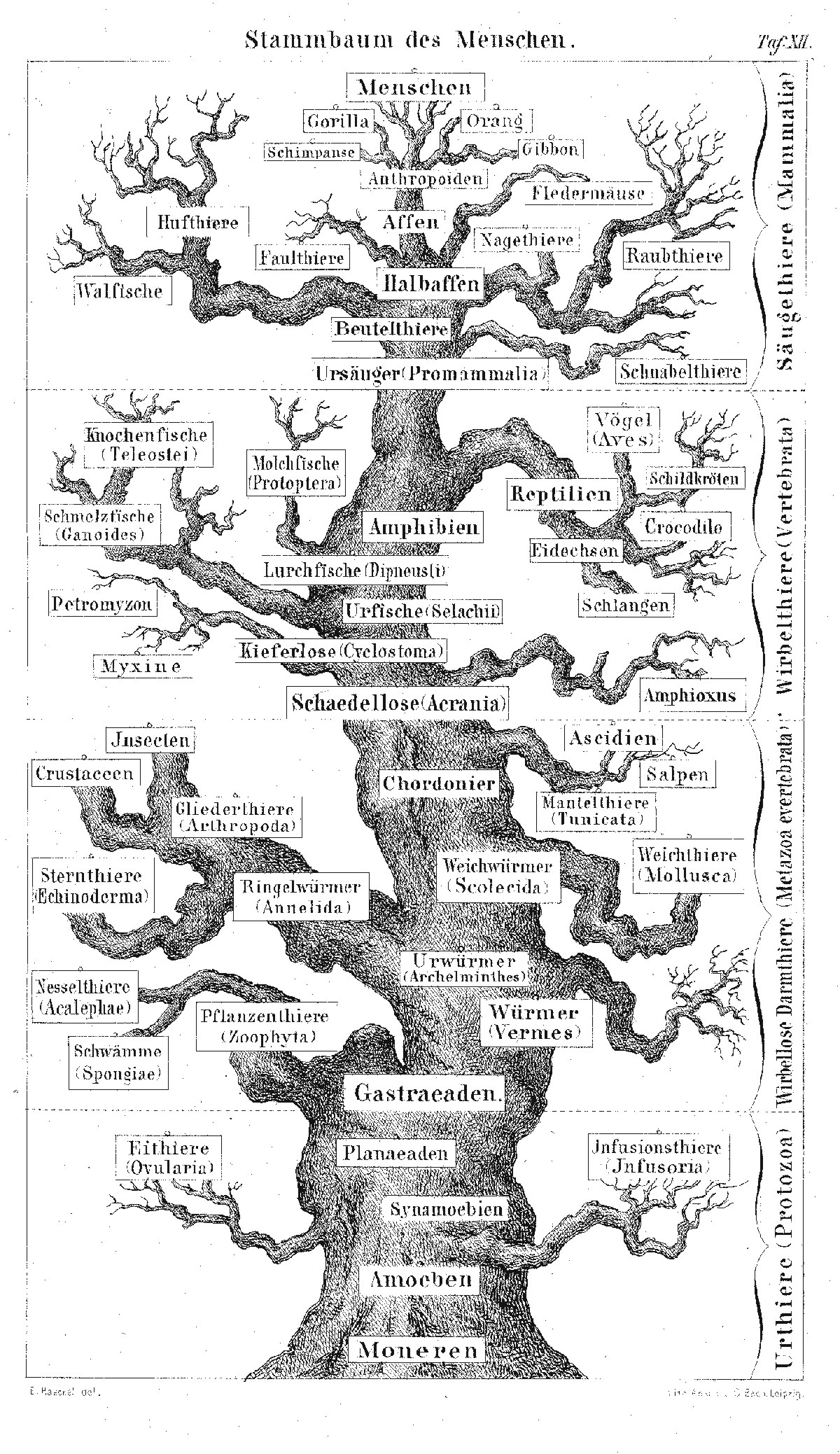
 Der Stammbaum des Menschen nach Ernst Haeckel
Der Stammbaum des Menschen nach Ernst HaeckelEinen gänzlich anderen Ansatz vertrat hingegen Ernst Haeckel. Schon 1868 hatte er in seiner Natürlichen Schöpfungsgeschichte geschrieben, dass „die meisten Anzeichen auf das südliche Asien“ hindeuteten.[47] Haeckel stützte seine Mutmaßung vor allem auf den Vergleich von Behaarung, Hautfarbe und Schädelform der damals als primitiv geltenden, heute als indigen bezeichneten Völker Afrikas und Asiens mit den Menschenaffen. Haeckel räumte jedoch zugleich ein: „Vielleicht war aber auch das östliche Afrika der Ort, an welchem zuerst die Entstehung des Urmenschen aus den menschenähnlichen Affen erfolgte; vielleicht auch ein jetzt unter den Spiegel des indischen Oceans versunkener Kontinent, welcher sich im Süden des jetzigen Asiens einerseits östlich bis nach den Sunda-Inseln, andrerseits westlich bis nach Madagaskar und Afrika erstreckte.“
Haeckels Hypothese, die Sunda-Inseln seien der Rest eines versunkenen Kontinents, auf dem sich vorzeitliche Menschenaffen zu den Vorfahren des Menschen und der anderen jetztzeitlichen Menschenaffen entwickelten, faszinierte den jungen holländischen Militärarzt Eugène Dubois. Er ließ sich deshalb 1877 nach Sumatra versetzen, um im Gebiet des indonesischen Archipels nach Fossilien zu suchen. In seinem Buch Die Frühzeit des Menschen beschreibt Friedemann Schrenk Dubois' Vorgehensweise wie folgt: „Besessen von seiner Idee, begann er an einer Stelle in Java zu graben, die nach heutigen Vorstellungen als völlig aussichtslos gelten würde. Er grub in einem Gebiet, wo im Umkreis von Tausenden von Kilometern noch nie zuvor auch nur die kleinste Andeutung von Resten eines Urmenschen gefunden wurde – und er grub auf den Zentimeter genau an der richtigen Stelle.“[48] Dubois kannte allerdings Hinweise von Bauern, die dort Tierfossilien gefunden hatten.[49]
Haeckels anatomische Vergleiche, Dubois’ Fossil des Java-Menschen von 1891 und die Entdeckung des Peking-Menschen in den 1920er-Jahren führten dazu, dass Darwins Hinweis auf Afrika als die Wiege der Menschheit nicht weiter beachtet wurde – stattdessen galt Asien unter den führenden Forschern als jene Region, in der sich der moderne Mensch entwickelt hatte. Daran konnte zunächst auch der Fund eines rund zwei Millionen Jahre alten, gut erhaltenen fossilen Schädels nichts ändern, der 1924 in der heute zu Südafrika gehörigen Ortschaft Taung entdeckt wurde. Raymond Dart, ein Anatom der University of the Witwatersrand in Johannesburg, erkannte die Bedeutung des Fundes und publizierte ihn Anfang 1925 in der Fachzeitschrift Nature unter dem neuen Art- und Gattungsnamen Australopithecus africanus.[50]
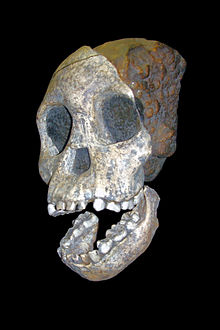 Das Kind von Taung
Das Kind von TaungDank seiner ärztlichen, neuroanatomischen Ausbildung hatte Raymond Dart erkannt, dass das so genannte Kind von Taung zwar ein affenähnliches Gesicht besaß; sein Gehirn und die Bezahnung waren jedoch menschenähnlich. Dart argumentierte daher, dass wichtige Merkmale des kleinen Schädels stärker menschenähnlich als affenähnlich seien: „Die Wangenbeine, die Jochbögen, Oberkiefer und Unterkiefer lassen zarte, menschenähnliche Charakteristika erkennen.“ Ähnliches gelte für das Gehirn, das gleichfalls mehr menschenähnliche als affenähnliche Merkmale aufweise: Das Kind von Taung sei daher einzuordnen als Mosaikform, das heißt als Mitglied „eines ausgestorbenen Geschlechts von Affen, das ein Zwischenglied darstellt zwischen den Menschenaffen der Gegenwart und dem Menschen“.[51] Darts Interpretation des Fossils wurde allerdings bis in die 1940er-Jahre nicht ernst genommen.
Ein wesentlicher Grund hierfür war die Ablehnung durch den damals führenden US-amerikanischen Paläontologen Henry Fairfield Osborn, der als Kritiker von Darwins Evolutionstheorie das theoretische Konzept eines so genannten Dawn-Man (Frühzeit-Mensch oder Mensch der Morgendämmerung) popularisiert hatte. Osborn behauptete unter anderem, das Gehirn des modernen Menschen sei derart komplex, dass zwei oder drei Millionen Jahre nicht hätten ausreichen können, um es aus einem affenähnlichen Gehirn hervorzubringen. Nahezu alle Forscher schlossen sich in den 1920er-Jahren dieser These an und unterstellten eine Zeitspanne von mindestens 20 bis 25 Millionen Jahren, während der die modernen Menschen und die modernen Menschenaffen sich getrennt entwickelt hätten – eine These, die keineswegs neu war, denn schon Rudolf Virchow hatte aus ähnlichen Überlegungen heraus den Neandertaler als fossile Art verkannt. Diese heute völlig willkürlich wirkende Annahme fand in den 1920er-Jahren jedoch eine plausible Stütze im Piltdown-Menschen, dessen Knochen ab 1912 in Sussex aufgesammelt worden waren[52] und der erst 1953 als Fälschung entlarvt wurde. Sein Kopf bestand aus dem Schädel eines modernen Menschen, dem der Unterkiefer eines Affen angepasst worden war. Die Fälschung kam der herrschenden Auffassung dieser Zeit entgegen, nach der die Vorfahren des Menschen bereits seit langer Zeit ein besonders großes Gehirn besaßen und versperrte so den damals führenden britischen und US-amerikanischen Paläontologen den Blick auf die tatsächlichen Gegebenheiten.
Die Wende zugunsten einer Anerkennung der Australopithecinen als Vormenschen trat erst ein, als die Aussagekraft anderer Fossilien neu bewertet wurde. So waren immer mehr Neandertaler entdeckt worden, deren Körperbau einheitlich war und die somit die These unhaltbar machten, das zuerst gefundene Exemplar sei ein deformierter, moderner Mensch gewesen. Ab 1936 waren zudem weitere Australopithecus-Fossilien in Afrika entdeckt worden, die von einer nachgewachsenen Forschergeneration nicht mehr in erster Linie vor dem Hintergrund der älteren Lehrmeinungen zur Dauer der Evolution des Gehirns gedeutet wurden.
Besonders aussagekräftige Fossilien wurden ab Ende der 1950er- / Anfang der 1960er-Jahre durch das Ehepaar Louis Leakey und Mary Leakey und später von deren Sohn Richard Leakey und seiner Ehefrau Meave Leakey in der Olduvai-Schlucht im heutigen Tansania sowie am Turkana-See in Kenia entdeckt. Seit den 1970er-Jahren bargen Forscher um Yves Coppens, Donald Johanson und Tim White zahlreiche Fossilien im Afar-Dreieck in Äthiopien. 1991 entdeckte der deutsche Forscher Friedemann Schrenk in Malawi das bislang älteste der Gattung Homo zugeschriebene Fossil, den Unterkiefer UR 501 von Homo rudolfensis. Das vollständigste bisher entdeckte Skelett eines frühen Vertreters der Hominini ist das von Ronald J. Clarke bearbeitete Fossil Little Foot aus Sterkfontein (Südafrika), an dessen Freilegung seit 1997 gearbeitet wird.
 Ausgrabungsarbeiten in der Höhle von Arago
Ausgrabungsarbeiten in der Höhle von AragoWichtige Fundstellen in Europa sind die Sierra de Atapuerca in Spanien, wo eine Besiedelung seit 1,2 Millionen Jahren gesichert zu sein scheint, und zwar beginnend bei Homo erectus / Homo antecessor über Homo heidelbergensis bis zum Neandertaler und zu Homo sapiens; die Höhle von Arago in Südfrankreich, in der 450.000 Jahre alte Homo erectus-Fossilien geborgen wurden, hier genannt Mensch von Tautavel; Peștera cu Oase in Rumänien (der Fundort der ältesten Fossilien von Homo sapiens in Europa) sowie Cro-Magnon in Frankreich, der Namensgeber des Cro-Magnon-Menschen.
Siehe auch
Literatur
- Robert Boyd, Joan B. Silk: How Humans Evolved. 5th edition. W. W. Norton & Company, New York NY u. a. 2009, ISBN 978-0-393-93271-3.
- Jared Diamond: Der dritte Schimpanse. Evolution und Zukunft des Menschen. 2. Auflage. Fischer-Taschenbuch-Verlag, Frankfurt am Main 1999, ISBN 3-596-14092-7 (Fischer 14092).
- Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Eine Einführung. Springer Verlag, Berlin u. a. 1999, ISBN 3-540-64831-3.
- G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag, Heidelberg 2008, ISBN 978-3-8274-1915-6.
- Friedemann Schrenk, Timothy G. Bromage, Henrik Kaessmann: Die Frühzeit des Menschen. Zurück zu den Wurzeln. In: Biologie in unserer Zeit. 32, H. 6, 2002, ISSN 0045-205X, S. 352–359.
- James Shreeve: Aus Afrika in die Welt. Was das Erbgut über unsere Herkunft und die Besiedlung der Erde erzählt. In: National Geographic Deutschland. Heft 3, 2006, ISSN 1615-0872, S. 38–53.
- Phillip Tobias u. a. (Hrsg.): Humanity from African Naissance to Coming Millennia. Colloquia in Human Biology and Palaeoanthropology. Firenze University Press, Florenz 2001, ISBN 88-8453-003-2, Volltext.
- Jeffrey H. Schwartz, Ian Tattersall: Fossil evidence for the origin of Homo sapiens. In: American Journal of Physical Anthropology, Band 143, Supplement 51 (= Yearbook of Physical Anthropology), 2010, S. 94–121, doi:10.1002/ajpa.21443
Weblinks
- „Der Ursprung des Menschen: Auf den Spuren der Hominiden.“ – Dossier von spektrumdirekt vom 12. Januar 2010
- „Die Entwicklungsgeschichte des menschlichen Stammbaums“ – Umfangreiche Übersichtsarbeit
- „Evolution des Menschen.“ – Film vom 16. Juli 2008 in der ZDFmediathek
Einzelnachweise
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, S. 4
- ↑ „Ein Fossil kommt vielleicht auf 100, auf 1000 Generationen. Im besten Fall gibt es mit jedem neuen Fund eine neue Hypothese, die etwas wahrscheinlicher ist als die alte. Das ist, als wollten Sie mit zwei Knochen die Geschichte des Römischen Reichs rekonstruieren.“ Friedemann Schrenk in: Spektrum der Wissenschaft, Nr. 9/2010, S. 72
- ↑ Bernard Wood, Terry Harrison: The evolutionary context of the first hominins. In: Nature, Band 470, 2011, S. 347–352, doi:10.1038/nature09709
- ↑ Zum Beispiel in Friedemann Schrenk, Die Frühzeit des Menschen. Der Weg zum Homo sapiens. C. H. Beck, 5., vollständig neubearbeitete und ergänzte Auflage, München 2008 (C.H.Beck Wissen), ISBN 978-3-406-57703-1
Auch in Grzimeks Tierleben wird Homo erectus als „Frühmensch“ bezeichnet; siehe dazu die „Systematische Übersicht“ in: Grzimeks Tierleben, Band 11 (=Säugetiere 2), dtv, 1979, S. 508. Die Gattung Australopithecus wird hier erläuternd als „Gattung Vormensch“ bezeichnet. - ↑ Vincent M. Sarich, Allan C. Wilson: Immunological time scale for hominid evolution. In: Science, Band 158, 1967, S. 1200–1203; doi:10.1126/science.158.3805.1200
- ↑ Masami Hasegawa u. a.: Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. In: Journal of Molecular Evolution, Band 22, 1985, S. 160–174; Zusammenfassung
- ↑ a b Michael Heads: Evolution and biogeography of primates: a new model based on molecular phylogenetics, vicariance and plate tectonics. Zoologica Scripta, Band 39, Nr. 2, 2009, S. 107–127, doi:10.1111/j.1463-6409.2009.00411.x
- ↑ R. L. Stauffer u. a.: Human and Ape Molecular Clocks and Constraints on Paleontological Hypotheses. In: The Journal of Heredity, Band 92, 2001, S. 469–474; Volltext
Andere Autoren berechneten auch in jüngerer Zeit wiederum 26,8 Millionen Jahre, siehe Jan E. Janečka u. a.: Molecular and Genomic Data Identify the Closest Living Relative of Primates. In: Science, Band 318, 2007, S. 792–794, doi:10.1126/science.1147555 - ↑ Gen Suwa u. a.: A new species of great ape from the late Miocene epoch in Ethiopia. In: Nature, Band 448, 2007, S. 921–924; doi:10.1038/nature06113
- ↑ a b c d e f g Terry Harrison: Apes Among the Tangled Branches of Human Origins. In: Science, Band 327, 2010, S. 532–534, doi:10.1126/science.1184703, Volltext (PDF)
- ↑ Zum gleichen Befund kam 2010 eine weitere Studie, in die sowohl paläontologische als auch molekularbiologische Daten einbezogen wurden, siehe Richard D. Wilkinson u. a.: Dating Primate Divergences through an Integrated Analysis of Palaeontological and Molecular Data. In: Systematic Biology, Band 60, Nr. 1, 2011, S. 16–31, doi:10.1093/sysbio/syq054 (Volltext (PDF)). Das Entstehen der Primaten wurde in dieser Studie in die Zeit vor 84,5 Millionen (69,5–103,5) Jahren datiert, die Trennung der Gibbons und der Menschenaffen in die Zeit vor 19,2 (15,1–24,1) Millionen Jahren.
- ↑ C. Owen Lovejoy u. a.: The Great Divides: Ardipithecus ramidus Reveals the Postcrania of Our Last Common Ancestors with African Apes. In: Science, Band 326, 2009, S. 73, doi:10.1126/science.1175833. Die Abzweigung der Orang-Utans datierte er in die Zeit vor rund 16 Millionen Jahren, die Abzweigung der Gorillas in die Zeit vor knapp 8 Millionen Jahren. Zu ähnlichen Ergebnissen für die Trennung der Hominini von den Schimpansen – 4,98 bis 7,02 Millionen Jahre – kamen auch: Sudhir Kumar u. a.: Placing confidence limits on the molecular age of the human–chimpanzee divergence. In: PNAS, Band 102, Nr. 52, 2005, S. 18842–18847, doi:10.1073/pnas.0509585102 (Volltext); Bernard Wood und Terry Harrison nannten 2010 die Zeitspanne zwischen 6 und 4 Millionen Jahren als die „wahrscheinlichste“: The evolutionary context of the first hominins. In: Nature, Band 470, 2011, S. 347–352, doi:10.1038/nature09709; vergl. dazu: „Als sich die Stammbäume von Schimpansen und Menschen teilten.“ – Wissenschaft-Online vom 20. Dezember 2005
- ↑ Jan E. Janečka u. a.: Molecular and Genomic Data Identify the Closest Living Relative of Primates. In: Science, Band 318, 2007, S. 793, doi:10.1126/science.1147555
- ↑ vereinfacht nach: Michael J. Benton: Vertebrate Palaeontology. 3. Aufl., Wiley-Blackwell, Malden 2004, ISBN 978-0-632-05637-8
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, S. 55
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen, S. 58
- ↑ Fiorenzo Facchini: Die Ursprünge der Menschheit. Konrad Theiss Verlag, 2006, S. 58
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen, S. 67
- ↑ Terry Harrison: A new species of Micropithecus from the middle Miocene of Kenya. In: Journal of Human Evolution, Band 18, Nr. 6, 1989, S. 537–557, doi:10.1016/0047-2484(89)90017-1
- ↑ Die Darstellung der ökologischen Einflüsse auf den Artenwandel folgt Terry Harrison, Apes Among the Tangled Branches of Human Origins, in Science, Band 327, 2010, S. 532–534.
- ↑ Salvador Moyà-Solà u. a.: A unique Middle Miocene European hominoid and the origins of the great ape and human clade. PNAS, Band 106 (24), 2009, S. 9601–9606, doi:10.1073/pnas.0811730106
- ↑ J. Agusti, S. Moyà-Solà: Mammal extinctions in the Vallesian (Upper Miocene). In: Lecture Notes in Earth Sciences, Band 30, 1990, S. 425–432, doi:10.1007/BFb0011163
- ↑ J. Agustí u. a.: Explaining the end of the hominoid experiment in Europe. Journal of Human Evolution, Band 45 (2), 2003, S. 145–153, doi:10.1016/S0047-2484(03)00091-5
- ↑ Kevin T. Uno u. a.: Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. In: PNAS, Online-Vorabveröffentlichung vom 4. April 2011, doi:10.1073/pnas.1018435108
- ↑ Caro-Beth Stewart, Todd R. Disotell: Primate evolution – in and out of Africa. Current Biology, Band 8 (16), 1998, S. R582–R588, doi:10.1016/S0960-9822(07)00367-3
- ↑ Yutaka Kunimatsu u. a.: A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans. PNAS, Band 104 (49), 2007, S. 19220–19225, doi:10.1073/pnas.0706190104
- ↑ Nick Patterson, Daniel J. Richter, Sante Gnerre, Eric S. Lander, David Reich (2006): Genetic evidence for complex speciation of humans and chimpanzees. In: Nature, Bd. 441 (29. Juni 2006), S. 1103–1108.
- ↑ Alexey V. Fedorov u. a.: Tropical cyclones and permanent El Niño in the early Pliocene epoch. In: Nature, Band 463, 2010, S. 1066–1070, doi:10.1038/nature08831
- ↑ Thure E. Cerling et al.: Woody cover and hominin environments in the past 6 million years. In: Nature, Band 476, 2011, S. 51–56, doi:10.1038/nature10306
- ↑ a b c Friedemann Schrenk, Timothy Bromage: Der Hominiden-Korridor Südostafrikas. In: Spektrum der Wissenschaft, Nr. 8/2000, S. 51–51
- ↑ Fiorenzo Facchini: Die Ursprünge der Menschheit. Konrad Theiss Verlag, 2006, S. 110
- ↑ Bernard Wood, Mark Collard: The Human Genus. Science, Band 284, 1999, S. 65–71, doi:10.1126/science.284.5411.65
- ↑ G. J. Sawyer, Viktor Deak: Der lange Weg zum Menschen. Lebensbilder aus 7 Millionen Jahren Evolution. Spektrum Akademischer Verlag, Heidelberg, 2008, S. 85
- ↑ Winfried Henke, Hartmut Rothe: Stammesgeschichte des Menschen. Springer Verlag, Berlin 1999, S. 156
- ↑ Friedemann Schrenk: Die Frühzeit des Menschen. Der Weg zum Homo sapiens. C. H. Beck, München 1997, S. 70
- ↑ Johannes Krause, Qiaomei Fu, Jeffrey M. Good, Bence Viola, Michael V. Shunkov, Anatoli P. Derevianko und Svante Pääbo: The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. In: Nature, Band 464, 2010, doi:10.1038/nature08976
- ↑ David Reich u. a.: Genetic history of an archaic hominin group from Denisova Cave in Siberia. In: Nature, Band 468, Nr. 7327, 2010, S. 1053–1060, doi:10.1038/nature09710
- ↑ Maria Mednikova: A proximal pedal phalanx of a Paleolithic hominin from denisova cave, Altai. In: Archaeology, Ethnology and Anthropology of Eurasia, Band 39, Nr. 1, 2011, S. 129–138, doi:10.1016/j.aeae.2011.06.017 oder [1]
- ↑ Lluís Quintana-Murci u. a.: Genetic evidence of an early exit of Homo sapiens sapiens from Africa through eastern Africa. In: Nature Genetics, Band 23, 1999, S. 437–441, doi:10.1038/70550
- ↑ Max Ingman u. a.: Mitochondrial genome variation and the origin of modern humans. In: Nature, Band 408, 2000, S. 708–713; doi:10.1038/35047064
Ian McDougall u. a.: Stratigraphic placement and age of modern humans from Kibish, Ethiopia. In: Nature, Band 433, 2005, S. 733–736; doi:10.1038/nature03258 - ↑ Brenna M. Henn u. a.: Hunter-gatherer genomic diversity suggests a southern African origin for modern humans. In: PNAS, Band 108, Nr. 13, S. 5154–5162, doi:10.1073/pnas.1017511108
- ↑ Michael F. Hammer u. a.: Genetic evidence for archaic admixture in Africa. In: PNAS, Band 108, Nr. 37, 2011, S. 15123–15128, doi:10.1073/pnas.1109300108
- ↑ Edward Tyson: Orang Outang sive Homo silvestris: Or, The Anatomie of Pygmie Compared with that of a Monkey, an Ape, and a Man. London, 1699; als editierter Reprint unter dem Titel: Edward Tyson: A Philological Essay Concerning the Pygmies of the Ancients. (Hrsg. von Sir Bertram Coghill Alan Windle). Bibliothèque de Carabas Bd. 9, Verlag D. Nutt, 1894
- ↑ Johann Jacob Scheuchzer: Homo diluvii testis. Zürich, 1726
- ↑ William King: The reputed fossil man of the Neanderthal. In: Quarterly Journal of Science, Band 1, 1864. S. 88–97, Online
Zuvor hatte King den Namen bereits an anderer Stelle – quasi inoffiziell – vorgeschlagen, siehe William King: On the Neanderthal Skull, or Reasons for believing it to belong to the Clydian Period and to a species different from that represented by Man. British Association for the Advancement of Science, Notices and Abstracts for 1863, Part II, London, 1864, S. 81 f. - ↑ Charles Darwin: The Descent of Man, and Selection in Relation to Sex. London, John Murray, 1871, Band 1, S. 199: „In each great region of the world the living mammals are closely related to the extinct species of the same region. It is therefore probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere.“
- ↑ Ernst Haeckel: Natürliche Schöpfungsgeschichte. Gemeinverständliche wissenschaftliche Vorträge über die Entwickelungslehre im Allgemeinen und diejenige von Darwin, Goethe und Lamarck im Besonderen, über die Anwendung derselben auf den Ursprung des Menschen und andere damit zusammenhängende Grundfragen der Naturwissenschaft. Berlin, Georg Reimer, 1868, Kapitel 19 (Volltext)
- ↑ Friedemann Schrenk: Die Frühzeit des Menschen. Der Weg zu Homo sapiens. CH Beck, 1997, S. 81
- ↑ athenapub.com The Discovery of Java Man in 1891
- ↑ Raymond A. Dart: Australopithecus africanus: The Man-Ape of South-Africa. In: Nature, Band 115, 1925, S. 195–199, doi:10.1038/115195a0, Volltext (PDF)
- ↑ „an extinct race of apes intermediate between living anthropoids and man.“ R. Dart, in Nature (1925), S. 195 (im Original kursiv)
- ↑ Charles Dawson, Arthur Smith Woodward: On the Discovery of a Palæolithic Human Skull and Mandible in Flint-Bearing Gravel Overlying the Wealden (Hastings Beds) at Piltdown, Fletching (Sussex). In: Quarterly Journal of the Geological Society, Band 69, März 1913


Dieser Artikel wurde am 6. Juni 2010 in dieser Version in die Liste der exzellenten Artikel aufgenommen. Kategorien:- Wikipedia:Exzellent
- Evolution
- Paläoanthropologie










Wikimedia Foundation.