- Wollnashorn
-
Wollnashorn 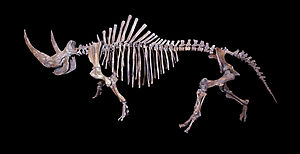
Skelett eines Wollnashorns
Zeitraum Pliozän bis Oberes Pleistozän 2,5 Mio. Jahre bis 11.000 Jahre Fundorte - Europa
- nördliches Asien
Systematik Überordnung: Laurasiatheria Ordnung: Unpaarhufer (Perissodactyla) Familie: Nashörner (Rhinocerotidae) Gattung: Wollnashörner (Coelodonta) Art: Wollnashorn Wissenschaftlicher Name Coelodonta antiquitatis Blumenbach, 1799 Das heute ausgestorbene Wollnashorn, auch Wollhaarnashorn oder Fellnashorn (Coelodonta antiquitatis) war eine in den Kältesteppen zwischen Westeuropa und Ostasien während des Mittel- und Jungpleistozäns verbreitete Art der Nashörner. Es gehörte zur Gruppe der Dicerorhinini, eurasischen Nashörnern mit zwei Hörnern, von denen heute nur noch das Sumatra-Nashorn (Dicerorhinus sumatrensis) existiert.
Inhaltsverzeichnis
Verbreitung und Lebensraum
 Lebendrekonstruktion eines Wollnashorns im Naturhistorischen Museum Wien
Lebendrekonstruktion eines Wollnashorns im Naturhistorischen Museum Wien
Verbreitung in Raum und Zeit
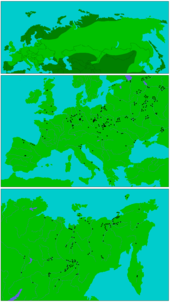 Verbreitungsgebiet des Wollnashorns. Oben: gesamtes Verbreitungsgebiet in Eurasien während der Weichsel-Kaltzeit. Darunter: Verbreitung in Europa (Mitte) und Nordostasien (unten) mit Darstellung wichtiger Fundorte (schwarze Punkte).
Verbreitungsgebiet des Wollnashorns. Oben: gesamtes Verbreitungsgebiet in Eurasien während der Weichsel-Kaltzeit. Darunter: Verbreitung in Europa (Mitte) und Nordostasien (unten) mit Darstellung wichtiger Fundorte (schwarze Punkte).Als kälteangepasstes Tier bevorzugte das Wollnashorn hauptsächlich boreales Klima und bewohnte die Steppen des nördlichen Eurasiens. Das Kerngebiet lag damit weitgehend im nördlichen Teil Innerasiens, vermutlich in Transbaikalien, im südlichen Sibirien und in der Mongolei.[1] Es ist aber mangels sicher datiertem Fundmaterials schwer einzugrenzen. Während der Kaltzeiten des Pleistozäns, dehnte es dann aber seinen Lebensraum weit nach Süden, Südwesten und Südosten aus. Zu einer ersten Expansionsphase kam es im Verlauf der Elster-Kaltzeit (vor etwa 400.000 bis 320.000 Jahren), während der es seine ursprünglichen Refugien stark erweiterte. Dabei erschien es erstmals in Mitteleuropa und ist in Mitteldeutschland im Harzer Gebiet nachgewiesen. Im Ausklingen dieser Vereisungsphase zog es sich jedoch wieder in seine ursprünglichen Gefilde zurück.
Eine weitere Expansionsphase erfolgte während der Saale-Kaltzeit (vor 300.000 bis 126.000 Jahren). Zu jener Zeit drang es bis nach Frankreich und auf die Britische Inseln vor und erreichte im nördlichen Teil der Iberischen Halbinsel und in Rumänien seine südlichsten Grenzen. Ebenso ist es erstmals an den Flussläufen von Don und Wolga nachgewiesen. Aber auch zum Ende dieser Kaltzeitphase verließen die Populationen ihre neugewonnenen Lebensräume wieder.[2][3]
Mit dem Ausklingen der Eem-Warmzeit und dem Beginn der Weichsel-Kaltzeit (vor 115.000 bis 11.700 Jahren) dehnte das Wollhaarnashorn sein Refugium vom Innerasien erneut weit nach Süden aus und erreichte in dieser Phase sein größtes Verbreitungsgebiet, was gleichzeitig das größte zusammenhängende Territorium war, das je von einer ausgestorbenen oder noch lebenden Nashornart besiedelt wurde.[4] So drang es im westlichen Eurasien bis auf die Iberische, die Apennin- und die Balkanhalbinsel bzw. die Peloponnes vor und erreichte hier seine äußerste südliche Verbreitungsgrenze bei etwa 37 bis 40° nördlicher Breite. Ebenso ist sein Vorkommen im Kaukasus und in Zentralasien (Kasachstan, Kirgisistan) belegt. Im östlichen Eurasien sind Funde aus dem Nordosten Chinas und von der Koreanischen Halbinsel bekannt, in China drang es sogar bis zum 33. nördlichen Breitengrad vor[3], während die östlichsten Fundpunkte auf Kamtschatka liegen. Gleichzeitig kam es auch zu einer Nordexpansion, in deren Zuge das Wollnashorn nach Jakutien bis zur Kolyma und zur Wrangel-Insel vordrang.[5]
Somit umfasst das Verbreitungsgebiet während seiner Maximalausdehnung fast das gesamte nördliche Eurasien, die besiedelte Fläche wird mit ungefähr 23,7 Mio. km² angegeben.[6] Ausgenommen bleiben die Regionen, die zu jener Zeit eisbedeckt waren (Fennoskandinavien, Nordosteuropa, Nordwestasien). Bemerkenswert ist aber, dass es auch nicht - oder nur sehr vereinzelt - im äußersten Nordosten Asiens auftritt und somit die Beringbrücke nicht erreichte, was notwendig gewesen wäre, um nach Amerika vorzustoßen. Warum dies dem Wollnashorn nicht gelang, obwohl andere Tierarten wie das Wollhaarmammut, das Bison, das Yak und letztendlich der Mensch diese Hürde zur selben Zeit nahmen, ist nicht vollständig klar.[7][4] Eine Erklärung dafür könnte sein, dass die Tierart nur in klimatisch günstigeren Phasen, die offensichtlich eine stärkere Durchfeuchtung der kaltzeitlichen Landschaften bedurften, in nordostasiatische Gebiete vordrang. Wahrscheinlich wirkte dann aber die Ausbildung stehender Gewässer auf dem Permafrostboden einer weiteren Ostexpansion entgegen und bildeten so eine Migrationsbarriere.[2][8] Andererseits wird angenommen, dass das Wollnashorn eher trockenes Klima bevorzugte und bei den damals vorherrschenden, eher feuchten Bedingungen im Nordosten, verbunden mit einem für ihn schlechteren Nahrungsangebot, die Tierart deshalb die Überquerung der Beringbrücke mied.[5]
Lebensraum
Das Wollnashorn lebte überwiegend in Flachlandgebieten, kam aber auch auf Hochflächen und Plateaus vor. Höhere Gebirgsregionen schien es aber aufgrund der Unzugänglichkeit gemieden zu haben. Sein Hauptlebensraum waren dadurch die weiten Tiefländer und Flussniederungen. Höhere Lagen hat die Tierart nur in für sie günstigen Klimaphasen aufgesucht. Weiterhin war es an weitgehend offene Landschaften mit trockenem bis aridem Klima gebunden.[5]
Ein wichtiger Lebensraum des Wollnashorns war die Mammutsteppe Eurasiens. Charakteristisch für diese Steppenform, die es so heute nicht mehr gibt, waren ausgedehnte, fast baumlose Gras- und Buschlandschaften, die aufgrund der hohen Sonneneinstrahlung über eine arten- und nährstoffreiche Vegetation verfügten.[9] Diese Mammutsteppe wurde vom Mammuthus-Coelodonta-Faunenkomplexes bewohnt, der im Jungpleistozän seine größte Ausdehnung erreichte. Neben dem Wollnashorn waren auch noch das mit-namengebende Wollhaarmammut, der Riesenhirsch, das Ren, die Saiga-Antilope und der Bison weitere wichtige Begleitelemente dieser Fauna.[2][9]
Aufgrund des großen Verbreitungsgebietes koexistierte das Wollnashorn auch häufig mit anderen Nashornformen. In den ausgehenden Warmzeiten, wenn das Wollnashorn langsam in die europäischen Gebiete vordrang, teilte es sich seinen Lebensraum teilweise mit dem Steppennashorn (Stephanorhinus hemitoechus),[6] in Osteuropa und Westasien tritt es zusammen mit dem Elasmotherium auf.[10] Im ostasiatischen mittelpleistozänen Nord-Tsingling-Faunenkomplex sind frühe Vertreter des Wollnashorns (ursprünglich als Coelodonta antiquitatis yenshanensis bezeichnet)[2] mit dem Waldnashorn (Stephanorhinus kirchbergensis) vergesellschaftet.[11]
Funde des Wollnashorns
Fundstellen allgemein
 Montiertes Wollnashornskelett, gefunden bei Bauarbeiten im Rhein-Herne-Kanal und ausgestellt im Ruhr Museum in Essen
Montiertes Wollnashornskelett, gefunden bei Bauarbeiten im Rhein-Herne-Kanal und ausgestellt im Ruhr Museum in EssenFunde vom Wollnashorn gehören in Eurasien zu den häufigsten Fossilfunden pleistozäner Tierarten. Die meist isolierten Knochen und Zähne stammen überwiegend aus Kies-, Sand- und Tongruben, die ehemalige Flussläufe oder Seeufer anzeigen. Darüber hinaus sind Funde teilweise auch aus Höhlen bekannt. Zudem werden Überreste von Wollnashörnern immer wieder submarin aus Schelfmeeren, wie der Nord- und Ostsee geborgen, die während der Kaltzeiten phasenweise trocken lagen.[12] Allein aus Polen ist das Wollnashorn von mehr als 50 Fundstellen bekannt,[13] während die Art in Spanien an wenigstens 20 Stellen registriert wurde.[3] Von etwas mehr als einem halben Dutzend Orten ist sie auch aus Italien bekannt.[14] Im östlichen Eurasien stellt das Wollnashorn allein in China mit mehr als 70 Fundplätzen die häufigste aller dort bekannten fossilen Nashornarten - die mehr als 60 Taxa von Eozän bis Pleistozän umfassen - dar.[15][16]
Auch in Deutschland ist das Wollnashorn mit einer hohen Funddichte vertreten, die allein in Westfalen mehr als 30 bekannte Fundorte einschließt.[12] Allerdings sind artikulierte Skelettreste oder gar vollständige Skelette äußerst selten. Eines der frühesten gefundenen Skelette stammt aus Pohlitz (Thüringen) und ist heute im Museum für Naturkunde in Gera (ebenfalls Thüringen) ausgestellt, ein weiteres konnte in einer Kies- und Sandgrube bei Petershagen (Nordrhein-Westfalen) geborgen werden und umfasst 66 Skelettelemente inklusive Schädel. Dieses befindet sich heute im Naturkundemuseum Bielefeld. Ein drittes ehemals artikuliertes Skelett aus der Bayerischen Staatssammlung für Paläontologie und Geologie in München wurde während des Zweiten Weltkrieges zerstört. Zahlreiche weitere montierte Skelette, so z. B. im Geologisch-Paläontologischen Museum der Westfälischen Wilhelms-Universität in Münster, im Museum für Ur- und Ortsgeschichte, Eiszeithalle Quadrat Bottrop oder im Ruhr Museum in Essen stellen dagegen meist Rekonstruktionen aus Skelettelementen mehrere in der jeweiligen Region aufgefundener Tiere dar.[17]
Funde mumifizierter Nashornkadaver
 Fund eines mumifizierten Kadavers vom Wollnashorn in Starunia (heute Ukraine) 1929
Fund eines mumifizierten Kadavers vom Wollnashorn in Starunia (heute Ukraine) 1929Wie beim Wollhaarmammut liegen auch vom Wollnashorn im Permafrost konservierte Kadaver vor, die Aufschluss über die Weichteilanatomie geben. Mit rund einem halben Dutzend wissenschaftlich registrierter Funde, die alle aus Jakutien stammen, sind diese aber weitaus seltener als beim Mammut. Einer der ersten bekannt gewordenen gefrorenen Kadaver wurde 1771 am Fluss Wiljui entdeckt.[9] Im Jahr 1858 kam ebenfalls am Wiljui in sandigen Ablagerungen ein nahezu vollständiges Skelett mit teilweise konservierten Hautresten zum Vorschein. Nur wenige Jahre später, 1877, entdeckte ein sibirischer Händler eine Nashornmumie am Fluss Chalbui, einem Nebenfluss des Bytantai, von dem er aber nur wenige Teile, u. a. den Kopf mit gut erhaltener Hautkonservierung bergen konnte, der Rest des Skelettes wurde im folgenden Frühjahrshochwasser weggespült.
Zu den bedeutendsten und aufschlussreichsten Funden ist jener von Tschuraptscha zwischen den Flussläufen der Lena und der Amga aus dem Jahr 1972 zu zählen. Wurden zunächst bei Tiefbauarbeiten nur einzelne Schädel- und Rippenfragmente gefunden, kam bei weiteren Untersuchungen im gleichen Jahr in einer Tiefe von rund 2,5 bis 3,6 m im gefrorenen Boden ein fast vollständiges Skelett mit erhaltenen Weichteilen an den rechten Hintergliedmaßen zum Vorschein. Kurze Zeit später, 1976 fanden Schüler während eines Ausflugs am linken Ufer des Aldan einen Schädel, ein Horn, einzelne Wirbel und andere Nashornknochen. Nachuntersuchungen erbrachten ein teilweise erhaltenes Skelett, darunter den Schädel mit beiden Hörnern und Bezahnung, den Unterkiefer und Wirbel.[18]
Aus dem Jahre 2007 stammt eine Eismumie vom Unterlauf der Kolyma. Diese umfasst ein Skelett, von dem der Schädel und die linke Körperpartie erhalten war, wobei sich Hautreste mit Haaren an den Gliedmaßen befanden.[19] Noch jünger sind die Funde aus dem Flussbett der Großen Tschukotschja aus dem Jahr 2008, die ein nahezu vollständiges Skelett mit Schädel und beiden erhaltenen Hörnern umfassen.[20] Im selben Jahr wurden teils mumufizierte Nashornreste von einem lokalen Einwohner nahe der Amga entdeckt und 2009 und 2010 in einer zweijährigen Grabung fachmännisch geborgen. Dabei kamen Beckenknochen, Vorder- und Hintergliedmaßen nebst Finger- und Zehenknochen, Schwanzwirbel und Rippen zum Vorschein.[18]
Außerhalb Jakutiens liegen zwei weibliche Wollnashörner aus Starunia (heute Ukraine) vor, die in einer Bitumen- bzw. Ölschiefer-Grube gefunden wurden. Der erste Kadaver kam 1907 zum Vorschein mit vollständigem Körper und Schädel und zwei unversehrten Hörnern. Eine weitere Mumie wurde 1929 gefunden, die ebenfalls vollständig, diesmal aber ohne Hörner war. Beide Kadaver wiesen eine sehr gute Weichteilerhaltung auf, allerdings ohne überlieferte Fellbedeckung.[21][22]
Aussehen und Lebensweise
 Skelett eines Wollnashorns ohne Horn
Skelett eines Wollnashorns ohne HornAllgemeines Aussehen
Informationen über den Körperbau, die Ernährungsweise und die Lebensgewohnheiten des Wollnashorns liegen wesentlich umfangreicher vor als zu jeder anderen pleistozänen Nashornart. Dies liegt daran, dass neben Resten von Knochen und Zähnen, teilweise auch vollständige Skelette, zudem noch im Permafrost Sibiriens konservierte Eismumien und in Salzlaugen und Ölschiefer erhaltene Kadaver und weiterhin Höhlenmalereien bzw. in Knochen und Steinen geritzte Zeichnungen überliefert sind. Das Wollhaarnashorn hatte eine Kopf-Rumpf-Länge von 340 bis 360 cm - hinzu kommt noch ein rund 50 cm langer Schwanz -, erreichte eine Widerristhöhe von 150 bis 170 cm und besaß ein Körpergewicht zwischen 1,5 und 2,9 t.[19][23] Weiterhin zeichnete es sich durch einen robusten Körperbau und kräftige breite Gliedmaßen mit deutlich ausgeprägten Muskelansatzstellen aus, was als Anpassung an ihre Lebensweise in offenen Landschaften angesehen wird.[24][7] Zudem besaß das Wollnashorn einen hohen und stark ausgeprägten Nackenbuckel, ähnlich wie beim Breitmaulnashorn (Ceratotherium simum), der ihm eine charakteristische Körperumrissform gab. Dieser Nackenbuckel entstand nicht nur durch die kräftigen Muskeln, die zur Stabilität des Schädels beitrugen, sondern diente auch als Fettspeicher, was besonders deutlich bei den Starunia-Funden untersucht werden konnte. Wie zudem die gefrorenen Kadaver zeigen, war es mit einem dichten Fell ausgestattet. Dieses bestand aus langen, stoppeligen Deckhaaren, die besonders an Nacken und Schultern gut ausgeprägt waren. Diese bedeckten eine dichte isolierende Unterwolle. Das Fell an den Gliedmaßen war demgegenüber wesentlich kürzer.[6]
Schädel- und Gebissmerkmale
 Schädel mit Hörnern
Schädel mit HörnernDer zwischen 70 und 90 cm messende Schädel[13] war sehr langgestreckt und besaß ein markant verlängertes Hinterhaupt, so dass das Wollnashorn seinen Kopf deutlich schräg nach unten trug. In diesem Merkmal ähnelte es dem heutigen Breitmaulnashorn oder seinem fossilen Verwandten, dem Steppennashorn. Auch Elasmotherium hatte eine ähnliche Kopfhaltung.[25][7] An dem lang ausgezogenen Hinterhauptsbein setzten kräftige Muskeln an, die zur Ausformung des bereits erwähnten Nackenbuckels beitrugen und notwendig waren, um den massiven Schädel zu halten.
Wie alle Dicerorhinini besaß es zwei Hörner, welche sich auf der Nase und dem mittleren Schädelbereich befanden. Die Ansatzstellen am Knochen sind perl- oder blumenkohlartig geraut, die extensive Ausprägung dieser Oberflächenrauhungen zeigen, dass das vordere Horn wesentlich größer als das hintere gewesen war. Das vordere Horn muss dabei auch im Verleich mit anderen Nashornarten besonders groß ausgeprägt gewesen sein, da die Ausdehnung dieser Oberflächen die größten Werte innerhalb der Dicerorhinini-Gruppe erreicht und auch jene des Steppennashorns übertrifft.[24]. Darauf weist weiterhin die im adulten Zusatand völlig verknöcherte Nasenscheidewand hin. Letztgenanntes Merkmal ist einzigartig für das Wollnashorn und kommt bei keiner rezenten Nashornart vor.[7]
Das Gebiss gleicht dem seiner fossilen Verwandten aus der Stephanorhinus-Line, das heißt es fehlten die Schneidezähne - diese kommen in der Regel nur im Milchgebiss vor und sind dort rudimentär ausgebildet[26] - während die Vorbackenzähne und Backenzähne jeweils dreifach je Kiefernbogen vorhanden sind. Diese sind sehr hochkronig, darin übertreffen sie noch jene des Steppennashorns, und besitzen viel Wurzelzement. Bemerkenswert ist vor allem, dass der zweite Prämolar besonders grazil ausgebildet ist.[24]
Hörner
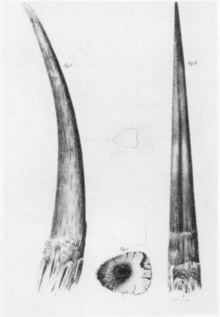 Fossiles Horn des Wollnashorns aus Jakutien, erstmals veröffentlicht von Johann Friedrich von Brandt 1849
Fossiles Horn des Wollnashorns aus Jakutien, erstmals veröffentlicht von Johann Friedrich von Brandt 1849Die Hörner sind ebenfalls im gefrorenen Zustand überliefert, meist vom vorderen größeren Horn (Nasalhorn). Dieses ist in seiner Form deutlich geschwungen und erreicht Längen bis zu 90 cm - wobei die vordere konvex verlaufende Kante über den Bogen gemessen bis zu 123 cm lang ist - bei einem Gewicht von rund 11 kg. Auch das hintere Horn (Frontalhorn) ist gelegentlich konserviert überliefert. Dieses hat einen häufig dreieckigen bis spitzovalen Umriss, ist teilweise auch leicht gebogen und erreicht Längen bis zu 40 cm. Das Gewicht dieses Horns liegt bei etwa 4,5 kg.[20][27]
Bemerkenswert ist, dass beide Hörner nicht rund im Querschnitt, wie bei heutigen Nashörnern und wie es die gewucherten Ansatzstellen vermuten ließen, sondern eher seitlich gestaucht bis teilweise brettförmig flach erscheinen. Ob diese Form der ursprünglichen entspricht oder durch spätere Schrumpfungsprozesse entstand, ist bisher nicht geklärt. Die Vorderkante des Horns reicht bis an die Nasenspitze. Interessanterweise treten an ihr häufig starke Schliffspuren auf. Diese entstanden vermutlich dadurch, dass das Tier mit seinem vorderen Horn die Schneedecke wegschob, um an die darunter liegenden Pflanzen zu gelangen oder aber bei der Nahrungsaufnahme und dem damit erbundenen Bodenkontakt das Horn einfach abwetzte.[27][28]
Beide Hörner sind durch horizontale Bänder aufgebaut, deren Breite häufig variiert. Bisher wurden maximal 35 solcher Bänder an einem einzelnen Horn gezählt. Frühe Annahmen brachten diese Bänder mit Wachstumsphasen der Hörner in Verbindung, die bei Nahrungsknappheit unterbrochen wurden. Bei rezenten Nashörnern sind solche Hornbänder ebenfalls, aber weitaus seltener nachgewiesen.[27][28] Neuere Untersuchungen zeigten, dass es sich bei den Bändern - ähnlich wie bei den Jahresringen der Bäume oder den konzentrischen Ringen der Elefantenstoßzähnen - um jährliche Zuwachsraten handelt, wobei jedes Band aus zwei Teilen besteht, einem dunkleren Abschnitt, der die Sommermonate und einem helleren Bereich, der die Wintermonate repräsentiert.[29][20]
Ursprünglich dachte man, dass die Hörner, die aus dicken, in Längsrichtung zur Hornachse verlaufenden Keratinfäden (sog. Filamente) bestehen, aufgrund ihrer großen Länge und ihres abgeflachten Querschnittes nicht für Rivalenkämpfe eingesetzt wurden, da seitliche Scherkräfte ein leichtes Abknicken ermöglicht hätten. Anhand rezenter Untersuchungen zur Hornstruktur des Wollnashorns wurde ermittelt, dass das Innere des Horns heterogen aufgebaut ist, wobei der Kern des Horns durch längere und dichter verwachsene Filamente deutlich härter ausgebildet ist als die äußeren Bereiche.[30][31]
Ernährungsweise
Zahlreiche anatomische Merkmale sprechen dafür, dass das Wollnashorn sich überwiegend grasend ernährte. Zu diesen Merkmalen gehören die schräge Haltung des Kopfes, die Hochkronigkeit der Zähne mit ihrem hohen Zementanteil und die grazile Ausbildung des zweiten Prämolaren.[24][7] Weiterhin wurde eine häufig horizontale Abschleifung der Zähne beobachtet, was von der in Gräsern enthaltenen Kieselsäure verursacht wird. Neben diesen skelettanatomischen Hinweisen zeigen u. a. die Mumie von 1877 aber vor allem jener 1929 in Starunia geborgene Kadaver eine gute Erhaltung der Schnauzenpartie, die, ähnlich den rezenten Breitmaulnashörnern, ein breites Maul - die Maulöffnung ist bis zu 35 cm breit - belegen mit strangartigen Lippen, die die Schneidezähne ersetzten und zum Rupfen von Gras geeignet waren.[6]
Weiterhin konnten an Zähnen einzelner Wollnashörner Pflanzenreste und aus dem Tschuraptscha-Nashorn von 1972 auch fossile Mageninhalte gewonnen und analysiert werden, die die Annahme einer weitgehend grasfressenden Ernährung bestätigten und darüber hinaus zeigten, dass diese Tierart Büsche und Zweige nicht grundsätzlich verschmähte.[5] So gehörten Süßgräser und Korbblüter wie Artemisia zu ihrem Nahrungsspektrum, ebenso wie Nadelgehölze, Weiden und Erlen.[32] Jüngst vorgenommene Isotopenuntersuchungen anhand fossiler Hörner zeigten weiterhin eine möglicherweise saisonabhängige Ernährungsweise. Die verschiedenen Wachstumszonen an den Hörnern ließen dabei unterschiedliche Nahrungsangebote vermuten. So ernährte sich das Wollnashorn im Sommer hauptsächlich grasend, während es im Winter teilweise Büsche und Zweige verzehrte.[29]
Individuelle Lebensweise
Das Wollnashorn erreichte ein Alter von 40 bis 45 Jahren und unterscheidet sich dadurch nicht von den rezenten Nashörnern. Die Individualentwicklung verlief ebenso ähnlich wie bei den heutigen Verwandten, was hauptsächlich mit Studien zum Zahnwechsel, die auf Vergleiche mit dem Breit- und Spitzmaulnashorn beruhen, nachgewiesen wurde.[26] Demzufolge wurden für die Gliederung der Ontogenese zehn Alterstufen vorgeschlagen. Die Geschlechtsreife trat wahrscheinlich mit dem fünften Lebensjahr ein. Ausgewachsene Wollnashörner hatten sicher keine natürlichen Feinde oder wurden nur selten von großen Beutegreifern wie dem Höhlenlöwen angegriffen. Kälber und Jungtiere waren aber einer solchen Gefahr ausgesetzt, wie es auch heute noch bei den rezenten Nashörnern der Fall ist. Einige Skelette von Jungtieren weisen im Hals- und Nackenbereich Spuren auf, die auf Bisse von großen Katzen zurückzuführen sind.[33] Darüber hinaus gab es aber auch noch andere Gefahren, denen das Wollnashorn gegenüberstand. Schlecht begehbares Gelände, steile oder rutschige Hänge u. ä. konnten zu schweren Stürzen bis hin zum Tod einzelner Individuen führen.[19]
Ob das Wollnashorn ein Einzelgänger war, ist nicht bekannt, die meisten heutigen Nashörner sind dies. Allerdings lebt das einzige heute bekannte auf Grasnahrung spezialisierte Nashorn, das Breitmaulnashorn in kleinen, matriarchalisch geführten Gruppenverbänden. Bullen sind dabei überwiegend Eizelgänger und leben territorial, wobei sie ihr Gebiet gegen Nahrungskonkurenten, sowohl im intra- (die eigene Art betreffend) als auch im interspezifischen (andere Tierarten betreffend) Wettbewerb verteidigen. Einige Schädel von Wollnashörnern zeigen Verletzungen auf, die auf Kämpfe mit anderen Nashörnern hinweisen. So treten Durchbohrungen am Schädel, häufig am Scheitelbein, aber auch an der Orbita oder an der Maxilla auf. Teile dieser Verletzungen führten später zu Osteomyelitis oder Arthrose. Auch gebrochene und wieder verwachsene Unterkiefer sind überliefert,[33] ebenso wie gebrochene und teils wieder verheilte Rippen, die ebenfalls auf solche Kämpfe schließen lassen.[12] In der Regel verliefen die Verletzungen durch Ritual- oder Territorialkämpfe nicht tödlich und die meisten Wollnashörner regenerierten sich offenbar wieder. Die relative Häufigkeit solcher Kampfverletzungen im Vergleich zu den rezenten Nashörnern wird auf die sich rasch ändernden Klimabedingungen während der letzten Kaltzeit zurückgeführt, wobei das Wollnashorn einer erhöhten Stresssituation in der Konkurrenz mit anderen großen und mittelgroßen Herbivoren der Mammutsteppe ausgesetzt war. Abseits von solchen Verletzungen wurden beim Wollnashorn auch Osteome beobachtet,[33] ebenso wie Zahnanomalien in Form von überzähligen Prämolaren und Molaren[26] oder das Auftreten der genetisch bedingten angeborenen Zahnschmelzhypoplasie.[34]
Der frühe Mensch und das Wollnashorn
Das Wollnashorn als Nahrungs- und Rohstofflieferant
Ob das Wollnashorn vom frühen Menschen tatsächlich gejagt wurde, ist unbekannt, direkte Nachweise für Jagd wie beim Wildpferd durch die Schöninger Speere und beim Europäischen Waldelefanten mit der Lanze von Lehringen fehlen hier. Kommen Reste von Wollnashörnern und menschliche Hinterlassenschaften zusammen vor, müssen diese nicht zwangsweise in einer kausalen Verbindung stehen. Die Fundstellenanzahl des gemeinsamen Auftretens ist zwar relativ hoch, doch meist kommen nur einzelne Zähnen oder Knochenfragmente vor, die selten mehr als einem Individuum zuzurechnen sind und mit einem hohen Prozentsatz zu Jungtieren gehören.[35] Bemerkenswert ist auch die große Häufigkeit von Wollnashornfunden in Höhlen, die nicht zum natürlichen Lebensraum der Tierart gehörten, so z. B. die Bocksteinhöhle (Baden-Württemberg) oder die Kůlna-Höhle (Tschechien). Da Höhlen jedoch sowohl von frühen Menschengruppen als Lagerplatz als auch von Hyänen als Horste genutzt wurden, kommen beide als potentielle Verbringer in Frage, Benagung der Knochen durch Hyänen lässt sich aber aufgrund des typischen Bissmusters relativ leicht erkennen.[17] Weiterhin kann ein gemeinsames Auftreten von Nashormresten und menschlichen Hinterlassenschaften auch auf eine natürliche Akkumulation mit kleinerem oder größerem zeitlichen Abstand zueinander zurückgehen. In solchen Fällen können Untersuchungen zu unterschiedlichen Verwitterungsgraden der Knochen und Zähne die Jagdfauna des Menschen von der natürlichen Hintergrundfauna abtrennen, wie es z. B. an der frühweichselkaltzeitlichen Station Salzgitter-Lebenstedt (Niedersachsen) der Fall war.[36]
Frühe gemeinsame Vorkommen von menschlichen Hinterlassenschaften und Nashornresten in Europa sind Markkleeberg (Sachsen), oder Romain-la-Roche (Frankreich). Solche Fundstellen aus der Zeit vor der Weichsel-Kaltzeit sind aber relativ selten und weitgehend handelt es sich, wie bei den beiden genannten Fundstellen, um natürliche und somit zufällige Assoziationen.[37][32] Mit der Weichsel-Kaltzeit nimmt die Anzahl der Fundplätze, aufgrund der nun sehr weiten Verbreitung des Wollnashorns, rasch zu und Nashornreste sind im späten Mittelpaläolithikum, das allgenmein mit dem Neandertaler verbunden wird, öfters nachgewiesen. Teilweise treten an den mittelpaläolithischen Fundstellen auch deutlich künstlich zerschlagene Knochen mit markanten Hiebspuren auf, wie es u. a. in der Gudenushöhle[38] (Österreich) oder an der Freilandstation Königsaue[39] (Sachsen-Anhalt) der Fall war. Hier kann man sicher von menschlichen Tätigkeiten ausgehen, wobei diese Tätigkeiten zum Teil mit dem Nahrungserwerb in Verbindung stehen. Das Aufschlagen, vor allem der Röhrenknochen, dient dazu, an das energiereiche Knochenmark zu gelangen. Zum Anderen steht das Zerschlagen der Kochen mit der in dieser Zeit aufkommenden stärkeren Nutzung organischer Rohstoffe wie Knochen, Geweih oder Elfenbein zu Herstellung von Geräten, Werkzeugen oder gar Waffen im Zusammenhang[40]. An einzelnen Knochen, wie z. B. in Zwoleń (Polen) lassen sich auch Überarbeitungen für den späteren Gebrauch als Werkzeug feststellen, wiewohl hier auch Schnittspuren der Kadaverzerlegung auftreten.[41]
Im darauffolgenden Jungpaläolithikum, verbunden mit dem ersten Auftreten des anatomisch modernen Menschen (Homo sapiens) in Europa, ändert sich diese Situation nicht grundlegend, obwohl diese Zeit geprägt ist durch zahlreiche neue Innovationen in z. B. der Werkzeugherstellung oder in den Techniken der Jagdwaffen, die im zunehmenden Maße auf der Verarbeitung von organischem Rohmaterial fußt. Im frühen Abschnitt der jüngeren Altsteinzeit, dem Aurignacien, sind Reste des Wollnashorns noch vergleichsweise häufig auf menschlichen Stationen vertreten.[42] So sind in der Vogelherdhöhle (Baden-Württemberg) mindestens 124 Knochenreste von wenigsten zwölf Individuen überliefert,[43] wobei hier diskutiert wird, ob es sich tatsächlich um eine aktive Bejagung handelt oder ob das Wollnashorn weniger der Nahrungsbeschaffung als vielmehr der Rohstoffnutzung diente.
Nach dem Rückzug der Inlandgletscher, die vor 20.000 bis 16.000 Jahren ihre maximale Ausdehnung erreichten, und der zunehmenden Wiederbesiedlung des nordalpinen Europas durch Träger des Magdalenien im späten Jungpaläolithikum spielt das Wollhaarnashorn nur noch eine geringe Rolle in der Subsistenz. Reste dieser Tierart werden nur noch selten gefunden, wie z. B. in Gönnersdorf (Rheinland-Pfalz).[42] Das nur spärliche Auftreten des Wollnashorns an menschlichen Siedlungsplätzen jener Zeit fällt hierbei mit der Tatsache zusammen, dass diese Tierart jetzt nur noch relativ selten in Europa anzutreffen war und kurz darauf aus diesem Teil Eurasiens verschwand.
Das Wollnashorn in der jungpaläolithischen Kunst
 Teil des „Panneau der Pferde“ aus der Chauvet-Höhle (Replik). Ca. 31.000 BP, eventuell Aurignacien. Neben den Pferden sind außerdem drei Nashörner, zwei sich scheinbar gegenüberstehend, aber wohl ohne Bezug zueinander, und drei Auerochsen dargestellt.
Teil des „Panneau der Pferde“ aus der Chauvet-Höhle (Replik). Ca. 31.000 BP, eventuell Aurignacien. Neben den Pferden sind außerdem drei Nashörner, zwei sich scheinbar gegenüberstehend, aber wohl ohne Bezug zueinander, und drei Auerochsen dargestellt.Trotz der relativ seltenen Nachweise der aktiven Jagd des frühen Menschen auf das Wollnashorn muss dieses imposante Tier Eindruck auf die damaligen Jäger- und Sammlergruppen des Jungpaläolithikums gemacht haben, stellten sie das Tier doch mehrfach in der nun aufkommeneden Höhlen- und mobilen Kunst dar. Die Wiedergabe des Wollnashorns erfolgen in - häufig linker - Seitendarstellung und zeichnen nicht nur die charakteristische Rückenlinie mit dem hohen Nackenbuckel nach, wie sie auch von den mumifizierten Kadavern bekannt ist, sondern gibt auch die Tiefe Haltung des Kopfes wieder. Darüber hinaus finden sich auch die Darstellungen der Ohren und vor allem der langen Hörner, in einigen Fällen ist auch das Fell angedeutet. Die teils sehr realitätsnahen Zeichnungen und Gravuren geben neben den biologischen Untersuchungen einen tiefen Einblick in das Aussehen dieser pleistozänen Tierart.[27]
Einige der frühesten Abbildungen stammen aus der Grotte Chauvet (Frankreich), welche mit einem Alter von mehr als 31.000 Jahren wohl dem Aurignacien angehören.[44] Auf mehrerern Panelen sind alein hier 47 Darstellungen bekannt, rund ein Viertel aller Tierzeichnungen in der Höhle, die abwechselnd in roter oder schwarzer Farbe, aber auch als Gravur abgebildet wurden. Hervorzuheben sind die Zeichnungen zweier sich gegenüber stehender Nashörner - einer sehr seltenen Szenerie in der jungpaläolithischen Kunst - im Panneau der Pferde oder jene von vier einander folgenden Tieren im Panneau der Handpositive. Die einzelnen Tiere sind in sich wiederholenden Merkmalen dargestellt, wie die durch zwei Bogenlinien angedeuteten Ohren oder ein den Körper teilendes schwarzes Band. Herauszustellen ist die häufige Darstellung überlanger Hörner.
Darüber hinaus bemerkenswert ist die Darstellung eines mittels schwarzen Pigmentnen gezeichneten Wollnashorns im Schacht des toten Mannes in der Höhle von Lascaux (Frankreich).[45] Die Nashornzeichnungen in der Höhle von Rouffignac (Frankreich) scheinen die Tiere weidend wiederzugeben, während eine Abbildung von Font-de-Gaume (Frankreich) durch die höhere Kopfhaltung eine eher aufmerksame Stellung annehmen lässt. Auch in anderen Höhlen im Bereich der Frankokantabrischen Höhlenkunst sind Wollnashörner dargestellt. Hierzu zählen La Colombiére, Les Rebiéres und Les Combarelles (alle Frankreich) sowie El Castillo (Spanien).[27] Trotz der Häufigkeit des Wollnashorns in der Grotte Chauvet sind Darstellungen dieser Tierart vergleichsweise selten und stellen nur rund 1 % aller Tierzeichnungen in Höhlen dar.[46] Außerhalb der Höhlen Frankokantabriens sind außerdem die mit roten Pigmenten ausgeführten Zeichnungen von Wollnashörnern aus der Höhle von Kapova (Ural, Russland) bekannt, die dem späten Jungpaläolithikum angehören.[42]
Neben den aufgeführten Höhlenmalereien wurde das Wollnashorn auch mehrfach in der mobilen Kleinkunst des - vor allem späten - Jungpaläolithikums dargestellt. Besonders aufschlussreiche Darstellungen stammen aus der Freilandstation Gönnersdorf (Rheinland-Pfalz) und sind auf Schieferplatten eingraviert. Mindestens zehn Abbildungen dieser Tierart sind von hier bekannt, bleiben aber zahlenmäßig weit hinter denen von Pferd und Mammut zurück. Weitere Nashorndarstellungen stammen von der Teufelsbrücke bei Saalfeld und von der Kniegrotte bei Döbritz (beide Thüringen). Während erstere ebenfalls in ein Tonschiefergeröll eingeritzt wurde, befindet sich zweitere auf einem Rengeweihstück.[47]
Stammesgeschichte
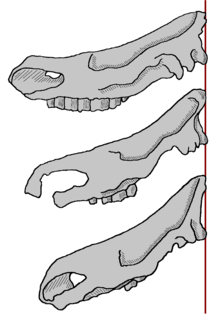 Schematische Darstellung der Änderung der Kopfhaltung des Wollnashorns während seiner Evolution. Oben: Coelodonta nihowanensis, Mitte: Coelodonta tologoijensis, unten Coelodonta antiquitatis.
Schematische Darstellung der Änderung der Kopfhaltung des Wollnashorns während seiner Evolution. Oben: Coelodonta nihowanensis, Mitte: Coelodonta tologoijensis, unten Coelodonta antiquitatis.Das Coelodonta entwickelte sich im Raum der Mongolei und Nordchina. Die ältesten Vertreter sind rund 2,16 bis 2,55 Mio. Jahre alt und wurden als Coelodonta nihowanensis bezeichnet. Es handelte sich dabei noch um relativ kleine Nashörner mit schlankem Körperbau. Fundstellen sind hier Longdan und Shitougu (beide Provinz Gansu, China). Das Auftreten dieser Gattung ist verbunden mit einer zunehmenden Ausbildung von aridem Klima im nördlichen China, was durch die gleichzeitige Entstehung äolischer Lössablagerungen in der Region gezeigt wird. Die jüngsten Funde des Coelodonta nihowanensis sind bisher etwa 1 Mio. Jahre alt.[6][48]
Vor etwa 750.000 Jahren entwickelte sich daraus Coelodonta tologoijensis, welches erstmals in Tologoj am Fluss Selenga (Burjatien, Russland) nachgewiesen ist. Im Vergleich zum späteren Wollnashorn ist diese Form noch wesentliche graziler mit schlankeren und längeren Gliedmaßen. Außerdem sind die Zähne weniger hochkronig und das Hinterhaupt nicht so stark verlängert. Frühmittelpleistozäne Funde aus dem Kusnezker Becken zeigen eine erste Westwärtsbewegung der Gattung Coelodonta an, eine erste Nordexpansion bis an die Lena erfolgt etwas nach 600.000 Jahren vor heute.
Zwischen dem ersten Auftreten von Coelodonta in Ost- und Zentralasien und seinem frühesten Erscheinen im westlichen Eurasien vor rund 400.000 Jahren klafft eine beträchtliche zeitliche sowie auch räumliche Lücke.[2] Die frühesten europäischen Funde sind elsterkaltzeitlichen Alters und stammen aus Brasov (Rumänien) und Mitteldeutschland, hauptsächlich aus der Region um den Harz. Die meisten Forscher weisen diese frühen europäischen Coelodonta-Formen dem klassischen Wollnashorn Coelodonta antiquitatis zu,[7][49] jedoch ist das Material aus dieser Zeit spärlich und beschränkt sich auf einzelne Zahnreste und Knochenfragmente. Lediglich ein fast vollständiger Schädel liegt vor, der aus Bad Frankenhausen (Thüringen) stammt. Dessen anatomische Merkmale, vor allem das relativ kurze Hinterhaupt erinnern noch stark an Coelodonta tologoijensis.[6][50] Erst aus der folgenden Saale-Kaltzeit sind deutlich mehr Funde überliefert. Hervorzuheben sind hier die Funde von La Fage und Romain-la-Roche (beide Frankreich). Letztere Fundstelle enthielt rund 225 Nashorn-Reste von neun Individuen, die der ausgehenden Saalekaltzeit angehören.[32]
Die innerartlichen Unterschiede zwischen den älteren spätmittelpleistozänen und den späteren jungpleistozänen Vertretern der europäischen Wollnashornfunde waren schon früh herausgearbeitet worden. Die Unterschiede betreffen vor allem die Zunahme an Robustizität, die Ausbildung kürzerer und kräftigerer Gliedmaßen und die Verlängerung des Hinterhauptes verbunden mit einer Verschlankung des Schädels. Aus diesem Grund wurde bereits 1980 eine Untergliederung in zwei Unterarten vorgeschlagen, wobei Coelodonta aniquitatis praecursor die mittelpleistozäne grazilere und Coelodonta antiquitatis antiquitatis die jungpleistozäne robustere Form darstellen.[24] Neuere Forschungen möchten die ältere Unterart eher einer späten Form des Coelodonta tologoijensis zuweisen und nur die jungpleistozänen Funde der Weichsel-Kaltzeit als klassisches Wollnashorn Coelodonta antiquitatis ansehen.[6] Diese Interpretation wird aber nicht vollständig geteilt.[32] Da bisher keine Klarheit in dieser Frage herrscht, müssen weitere Untersuchungen abgewartet werden. Eine ähnlich motivierte Teilung in zwei Unterarten wie im westlichen Europa wurde in Russland Ende der 1960er Jahre anhand jakutischer Funde durchgeführt und das ältere Coelodonta antiquitatis jacuticus von dem jüngeren Coelodonta antiquitatis humilis abgesetzt.[2]
Stammesgeschichtlich lässt sich das Wollnashorn nach jetzigem Stand aus den frühesten Coelodonta-Funden in Ostasien herleiten. Dabei entwickelte es sich von einer relativ kleinen und schlanken Nashornart zu einer großen robusten. Einhergehend damit sind skelettmorphologische Veränderungen am Schädel, wie die allmähliche Verlängerung des Hinterhauptes und der damit verbundenen Absenkung des Kopfes, die Verlagerung der Augen aufgrund dessen in den hinteren Schädelbereich - dadurch konnte das Wollnashorn bei der tiefen Kopfhaltung nach vorn schauen - die Zunahme der Kronenhöhe der Zähne mit einer gleichzeitigen Erhöhung des Zementanteils und die stärkere Ausprägung des Nasenbereiches, was wohl zu einer Größenzunahme des Hornes führte. Bezogen auf den Bewegungsapparat ist eine Entwicklung von im Verhältnis zum Körper langen schmalen Gliedmaßen zu kurzen breiten mit kräftigen Muskeln erkennbar, was als fortschreitende Anpassung an das Leben in offenen Landschaften anzusehen ist. Die evolutiven Veränderungen von Coelodonta nihowanensis über Coelodonta tologoijensis zu Coelodonta antiquitatis, welche mit der typischen Ausprägung des Wollnashorns in der Weichsel-Kaltzeit ihren Höhepunkt fanden, werden dabei als graduell angesehen.[6][48]
Das Wollnashorn starb am Ende der letzten Kaltzeit aus. Auf den britischen Inseln lassen sich die letzten Vertreter dieser Tierart vor ca. 35.000 Jahren nachweisen.[51] Aus Kontinentaleuropa verschwand sie wohl vor rund 14.000 Jahren während der Älteren Dryas. Das jüngste Datum stammt aus Gönnerdorf und liegt bei 13.600 Jahren vor heute, nur wenig älter sind die Funde von Vaumarcus (Schweiz). Im Gebiet des Urals und Westsibiriens scheint das Wollnashorn noch bis in die Jüngere Dryaszeit überlebt zu haben, da aus Lugovskoje Daten mit 10.700 Jahren vor heute angegeben werden.[52][53] Jüngere Funde, die teils aus Ostasien gemeldet wurden, so z. B. liegt ein gemessenes Datum bei 7.400 Jahren vor heute[54], werden vorläufig abgelehnt, da diese auf unterschiedlich verwendete Halbwertszeiten des 14C-Isotops bei der Radiokarbondatierung zurückgehen.[53] Ob Klimaveränderungen, Jagd durch den jung- und spätpalaolithischen Menschen oder beides für das Aussterben des Wollnashorns verantwortlich war, ist umstritten (siehe Hauptartikel Quartäre Aussterbewelle).
Klimatische Anpassung
 Lebendrekonstruktion (Illustration)
Lebendrekonstruktion (Illustration)Das Wollnashorn wird allgemein neben dem Wollhaarmammut als das klassische Tier der kaltzeitlichen Steppen Eurasiens angesehen. Die Vorstellungen über diese Tierart werden dabei hauptsächlich durch die Funde im sibirischen Permafrost und die Lagerung in hauptsächlich kaltzeitlichen Sedimenten der letzten Kaltzeit geprägt. Somit wird ihre Anwesenheit auch als Marker für die klimatische Rekonstruktion einer Fundstelle genommen.[24]
Der Ursprung des Wollnashorns liegt in den ariden Steppen Innerasiens. Seine Anpassung an diese offene Landschaftsform zeigen die breiten Gliedmaßen aber auch die charakteristische Ernährungsweise mit den daraus resultierenden hochkronigen Zähnen und der tiefen Kopfhaltung. Einige Paläontologen argumentieren, dass das Wollnashorn aber nicht vollständig an die glazialen Bedingungen angepasst war. Als Hinweise dafür werden die im Vergleich zum Wollhaarmammut nicht so hohe nördliche Verbreitung angesehen ebenso wie die Tatsache, dass es nicht den Übertritt nach Amerika schaffte. Vielmehr war diese Tierart nach Meinung jener Paläontologen an trocken-klimatische Verhältnisse gebunden, wie es ihr gehäuftes Auftreten in Transbaikalien zeigt. Dort herrschte während der Kaltzeiten ein sehr kontinetales Klima vor, welches ausgedehnten Steppen und teilweise Halbwüsten bedingte.[55][5]
Trockene Klimabedingungen bewirkten auch, dass Vertreter der Gattung Coelodonta im Mittelpleistozän in Ostasien gelegentlich in warmklimatischen Abschnitten vorkommen, so unter anderem an der wichtigen frühmenschlichen Fundstelle Zhoukoudian bei Peking. Diese ostasiatischen Vertreter der Wollnashörner teilten sich ihren Lebensraum mit dem Elephas namadicus, einer asiatischen Altelefantenart, dem Waldnashorn und teilweise auch mit dem Orang Utan (Pongo).[55] Auch in Europa kommen Vertreter dieser Tierart gelegentlich in warmzeitlichen Ablagerungen des Mittel- und Jungpleistozäns vor. So wurde es in den mehr als 200.000 Jahre alten Ablagerungen von La Fage (Frankreich) dokumentiert, wo es in einer Waldsteppe lebte,[32] Weitere Beispiele sind Funde des Wollnashorns aus den oberen Travertinen von Ehringsdorf (Thüringen][56] und jüngst aus dem Geiseltal.[24] An letztere Fundstelle, die der Eem-Warmzeit zugerechnet wird, konnte anhand makroskopischer Pflanzenreste ein stark kontinentales Klima nachgewiesen werden, das seine heutige ökologische Äquivalente im Alföld oder in den südrussischen Steppenräumen findet.[57] Auch im englischen Crayford wurde das Wollnashorn in der späten Eem-Warmzeit nachgewiesen. Die Tierart tolerierte offensichtlich nicht nur boreales Klima, sondern kam auch mit wärmeren Klimaten zurecht,[32] ihr Auftreten in den mittel- und jungpleistozänen Warmzeiten war aber wohl an kühlere oder sehr trockene Phasen gebunden.[6]
Während der Weichsel-Kaltzeit tritt dann das klassische Wollnashorn in seiner bekannten Erscheinungsform auf. Aufgrund des riesigen Ausbreitungsgebietes dieser Tierart während dieser Zeit erreichte es nicht nur den hohen Norden Sibiriens, es besiedelte auch weit in den Süden reichende Landschaftsräume. So ist das Wollnashorn in der zweiten Hälfte der Weichsel-Kaltzeit über weite Teile Südeuropas verbreitet.[2] Durch dieses südliche Auftreten überschneidet sich sein Areal teilweise auch mit den Rückzugsräumen einiger warmzeitlicher Faunenelemente. So ist das Wollnashorn u. a. in sandigen Ablagerungen bei Ingarano (Süditalien; Schicht b) zusammen mit dem Europäischen Waldelefanten und dem Flusspferd überliefert.[58]
Systematik
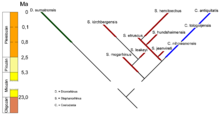 Verwandtschaftsverhältnis des Woolnashorns zu anderen Vertretern der Dicerorhinini. Dargestellt sind neben dem Sumatra-Nashorn nur die westeurasischen Vertreter dieser Nashorngruppe.
Verwandtschaftsverhältnis des Woolnashorns zu anderen Vertretern der Dicerorhinini. Dargestellt sind neben dem Sumatra-Nashorn nur die westeurasischen Vertreter dieser Nashorngruppe.Das Wollnashorn gehört zu den Dicerorhinini, einer Gruppe von zweihörnigen Nashörnern. Sein einziger heute noch lebender Verwandter ist das in den Wäldern Sumatras lebende und vom Aussterben bedrohte Sumatra-Nashorn. Neben den Gattungen Coelodonta und Dicerorhinus gehört zu dieser Gruppe auch das Stephanorhinus (vereinzelt auch Brandtorhinus genannt), deren beiden letzten Vertreter Waldnashorn und Steppennashorn ebenfalls im Mittel- und Jungpleistozän aber in differierenden Biotopen im nördlichen Eurasien lebten.[59]
Dabei bilden die Gattungen Coelodonta und Stephanorhinus eine enger verwandte Klade und sind monophylethischen Ursprungs, häufig wird auch von einer Coelodonta-Stephanorhinus-Gruppe gesprochen. Ein Hauptargument dafür ist die Verknöcherung der Nasenscheidewand, die so nur bei diesen beiden Gattungen auftritt.[28] Diese Verknöcherungen beginnen evolutiv schon relativ früh bei den Stephanorhinus-Formen und schreiten über das Waldnashorn zum Steppennashorn immer weiter fort, bis sie schließlich ihren Höhepunkt beim Wollnashorn erreichen. Eine weitere Gemeinsamkeit von Coelodonta und Stephanorhinus ist das Fehlen der Schneidezähne im Dauergebiss, während sie rudimentär im Milchgebiss ausgebildet sind. Das Dicerorhinus dagegen besitzt permanente Schneidezähne, wenn auch im rückgebildeten Zustand.[60][59] Deshalb plädieren einige Forscher dafür, die Gattungen Stephanorhinus und Coelodonta zusammenzuführen. Es wird jedoch häufig darauf hingewiesen, dass beide als eigenständige Gattungen beschrieben wurden.[24]
Aufgrund der endemischen Verbreitung des Wollnashorns in Eurasien und einzelnen anatomischen Übereinstimmungen wurde schon früh eine eher nähere Verwandtschaft mit den rezenten asiatischen als mit den afrikanischen Nashörnern vermutet. Molekulargenetische Untersuchungen mit Hilfe der mitochondrialen DNA von Wollnashörnern aus der Scladina-Höhle Belgien ergaben, dass diese Tierart tatsächlich mit dem Sumatra-Nashorn am nächsten verwandt ist. Beide Linien trennten sich demnach im späten Oligozän oder frühen Miozän vor rund 21 bis 26 Mio. Jahren.[61] Bereits vor mehr als 29 Mio. Jahren spalteten sich die geinsamenen Vorfahren der Dicerorhinini und der Rhinocerotini, zu denen das Indische Panzernashorn (Rhinoceros unicornis) und das Java-Nashorn (Rhinoceros sondaicus) gehören, von den afrikanischen Nashörnern ab.[62]
Forschungsgeschichte
 Johann Friedrich Blumenbach
Johann Friedrich Blumenbach Das Handbuch der Naturgeschichte von Johann Friedrich Blumenbach aus dem Jahr 1799 (6. Ausgabe) mit der Erstbeschreibung des Wollnashorns.
Das Handbuch der Naturgeschichte von Johann Friedrich Blumenbach aus dem Jahr 1799 (6. Ausgabe) mit der Erstbeschreibung des Wollnashorns.Eine der frühesten wissenschaftlichen Erwähnungen einer einst im nördlichen Eurasien lebenden Nashornart erfolgte 1769, als der deutsche Naturforscher Peter Simon Pallas (1741–1811) einen Bericht über seine Expeditionen nach Sibirien verfasste und dabei einen aufgefundenen Schädel und zwei Hörner einer solchen Tierart aus dem dortigen Permafrost erwähnte.[27][21] Von ihm stammte auch die bereits 1772 eingeführte Bezeichnung Rhinoceros lenenesis,[18] wobei den zur Namensgebung führenden Untersuchungen mehrere Skelettfunde aus Sibirien inklusive der 1771 entdeckten Eismumie vom Wiljui-Fluss zu Grunde lagen. Der Begriff ist heute weitgehend nicht mehr in Gebrauch, gelegentlich benutzten ihn einige russische Forscher in jüngerer Zeit für jakutische Funde,[1][21] was aber allgemein nicht akzeptiert wird.[14][63] Nur wenige Jahre später, 1782, bestimmte Johann Heinrich Merck (1741–1791) erstmals fossile Nashornfunde aus Deutschland, so u. a. aus dem Odenwald. Die heute gültige wissenschaftliche Erstbeschreibung, damals mit dem Namen Rhinoceros antiquitatis, erfolgte 1799 vom deuteschen Zoologen und Anthropologen Johann Friedrich Blumenbach (1752–1840). Der Syntypus umfasst vor allem Funde aus der Umgebung von Göttingen in Niedersachsen.[24] Dazu zählen ein unvollständiger Oberarm aus Schwarzfeld und mehrere Knochen (u. a. ein Hinterhaupt, ein zweiter Halswirbel und mehrere Langknochen) sowie Zähne (zwei Prämolaren) aus Düna, die alle Mitte des 18. Jahrhunderts entdeckt wurden. Darüber hinaus schließt der Syntypus auch einen fragmentierten Schädel aus dem ehemaligen Gouvernement Ufa (Russland) ein, welcher der Baron Georg Thomas von Asch (rus. Егор Фёдорович Аш, Jegor Fjodorowitsch Asch; 1729–1807) etwa 1790 der Universität Göttingen ausgehändigt hatte und der erst 2005 wiederentdeckt wurde.[64][63] Vom deutschen Paläontologen Johann Gotthelf Fischer von Waldheim (1771–1853) stammt die Bezeichnung Rhinoceros tichorhinus, wobei sich der Artname auf die charakteristische, völlig verknöcherte Nasenscheidewand bezieht. Tichorhinus wurde später gelegentlich auch als Gattungsname verwendet, hat aber ebenso keine Gültigkeit. Der anerkannte Gattungsname Coelodonta hingegen wurde 1831 vom deutschen Geologen Heinrich Georg Bronn (1800–1862) etabliert.[24][9] Die heute gültige Bezeichnung Coelodonta antiquitatis kam erstmals während der 1930er Jahre auf.[64]
Während der Erforschung des Wollnashorns wurden unterschiedliche wissenschaftliche Namen für diese Tierart gebraucht:[13][14]
- Rhinoceros lenenensis Pallas 1773
- Rhinoceros antiquitatis Blumenbach 1799
- Rhinoceros tichorhinus Fischer 1814
- Rhinoceros Pallasi Desmarest 1822
- Coelodonta bojei Bronn 1831
- Rhinoceros tichorhinus Cuvier 1834-1836
- Rhinoceros tichorhinus Brandt 1849
- Rhinoceros tichorhinus Giebel 1851
- Rhinoceros antiquitatis Falconer 1868
- Rhinoceros jourdani Lortet und Chantre 1876
- Rhinoceros (Tichorhinus) antiquitatis Brandt 1877
- Rhinoceros Mercki Schrenck 1880
- Rhinoceros antiquitatis tichorhinus Lubicz-Niezabitowski 1914
- Tichorhinus antiquitatis (= tichorhinus) Wüst 1922
- Rhinoceros antiquitatis Hilzheimer 1924
- Tichorhinus antiquitatis Zeuner 1934
Das Coelodonta tologoijensis wurde erstmals 1966 von E. I. Beliajewa anhand von Funden vom Tologoj-Berg an der Selenga in der Region Transbaikalien (Russland) beschrieben. Den Namen Coelodonta nihowanensis dagegen führte der deutsche Paläontologe Hans-Dietrich Kahlke 1967 an Funden aus dem Nihewan-Becken (Provinz Hebei, China] ein, ohne dass diese Bezeichnung anerkannt wurde, das gleiche gilt für die von Chow B. S. 1978 erfolgte homonyme Benennung. Jedoch wurde später darauf hingewiesen, dass die Erstbeschreibung dieser Art unter Einhaltung der nomenklatorischen Regeln erfolgt war. Momentan gilt diese Artbezeichnung als nomen nudum.[2][48]
Kulturgeschichte
Funde eiszeitlicher Großsäuger regten in früheren Zeiten häufig die Phantasie der Menschen an, wie die Verwendung sog. Drachenknochen, zu Pulver zermahlen als medizinisches Allheilmittel in Ostasien angesehen, oder wie die Interpretation der Schädel von Zwergelefanten auf den östlichen Mittelmeerinseln als Reste von Zyklopen in der griechischen Antike zeigen.[65] Die im sibirischen Permafrost aufgefunden Nashornreste waren den einheimischen Völkern Jakutiens schon lange bekannt. Bis ins 19. Jh. verwendeten sie das fossile Horn des Wollnashorns dazu, die Elastizität ihrer Jagdbögen zu verbessern. Darüber hinaus interpretierten sie die Hörner, aufgrund des flachen Querschnittes und der gebogenen Form, als Krallen eines riesigen Vogels. Der deutsche Zoologe Gotthelf Fischer von Waldheim (1771–1853) verband diese sibirischen Legenden mit den antiken Geschichten um den Greif, der nach Herodot als Adler-Löwe-Mischwesen im Norden zwischen den Skythen und dem mythischen Hyperboräern lebte. Nach Meinung Fischers war der Ursprung diese Legende im Raum des Urals zu finden. Bereits 1823 hatten diese Berichte den deutschen Naturforscher Gotthilf Heinrich von Schubert (1780–1860) dazu veranlasst, den Namen Gryphus antiquitatis für diesen vermeintlichen "fossilen Greif" einzuführen.[27][63]
Auch in Europa wurden schon sehr früh Fossilien des Wollnashorns gefunden, so z. B. kam 1335 im Zollfeld bei Klagenfurt ein Nashorn-Schädel zum Vorschein. Dieser diente rund 250 Jahre später dem Bildhauer Ulrich Vogelsang als Vorbild für den Kopf des Drachens, den er am Lindwurmbrunnen in Klagenfurt gestalten sollte.
Literatur
- Paul S. Martin, Richard G. Klein (Hrsg.): Quaternary Extinctions. A Prehistoric Revolution. The University of Arizona Press, Tucson AZ 1984, ISBN 0-8165-1100-4.
- Arno Hermann Müller: Lehrbuch der Paläozoologie. Band 3: Vertebraten. Teil 3: Mammalia. 2. überarbeitete und erweiterte Auflage. Gustav Fischer Verlag, Jena 1989, ISBN 3-334-00223-3.
- Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Theiss-Verlag, Stuttgart 2002, ISBN 3-8062-1734-3.
Weblinks
 Commons: Coelodonta – Sammlung von Bildern, Videos und Audiodateien
Commons: Coelodonta – Sammlung von Bildern, Videos und Audiodateien- Das Wollnashorn
- Wissen aktuell: Eiszeitriesen eroberten Europa früher - Fellnashörner lebten schon vor 460.000 Jahren am Fuß des Kyffhäusergebirges
Einzelnachweise
- ↑ a b Нина В. Гарутт и Геннади Г. Боескоров: Шерстистые носороги: қ истории рода. GEOS 2001, S. 157-167
- ↑ a b c d e f g h Ralf-Dietrich Kahlke: Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des oberpleistozönen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Großsäuger). Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 546 Frankfurt am Main, 1994
- ↑ a b c Diego J. Álvarez-Lao und Nuria García: Geographical distribution of Pleistocene cold-adapted large mammal faunas in the Iberian Peninsula. Quaternary International 233, 2011, S. 159-170
- ↑ a b Donald R. Prothero, Claude Guérin und Earl Manning: The history of Rhinocerotoidea. In Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New-York, 1989, S. 321–340
- ↑ a b c d e Gennady Boeskorov: Woolly rhino (Coelodonta antiquitatis) distribution in Northeast Asia. Deinsea 8, 2001, S. 15-20
- ↑ a b c d e f g h i Ralf-Dietrich Kahlke und Frédéric Lacombat: The earliest immigration of woolly rhinoceros (Coelodonta tologoijensis, Rhinocerotidae, Mammalia) into Europe and its adaptive evolution in Palaearctic cold stage mammal faunas. Quaternary Science Reviews 27, 2008, S. 1951-1961
- ↑ a b c d e f H. Loose: Pleistocene Rhinocerotidae of W. Europe with reference to the recent two-horned species of Africa and S.E. Asia. Scripta Geologica 33, 1975, S. 1–59
- ↑ Scott A. Elias und Barnaby Crocker: The Bering Land Bridge: a moisture barrier to the dispersal of steppe–tundra biota? Quaternary Science Reviews 27, 2008, S. 2473–2483
- ↑ a b c d Wighart von Koenigswald: Lebendige Eiszeit. Klima und Tierwelt im Wandel. Stuttgart, 2002, S. 54–61
- ↑ Diana Pushkina: The Pleistocene easternmost distribution in Eurasia of the species associated with the Eemian Palaeoloxodon antiquus assemblage. Mammal Review 37, 2007, S. 224-245
- ↑ Hans-Dietrich Kahlke: Zur chronologischen Stellung der Choukoutien-Kultur. Alt-Thüringen 6, 1963, S. 22-41
- ↑ a b c Cajus G. Diedrich: A skeleton of an injured Coelodonta antiquitatis from the Late Pleistocene of north-western Germany. Cranium 25, 2008, S. 9-23
- ↑ a b c Magdalena Borsuk-Białynicke: Studies on the Pleistocene Rhinoceros Coelodonta antiquitatis (Blumenbach). Palaeontologia Polonica 29, Warschau-Krakau, 1973
- ↑ a b c Emmanuel M. E. Billia: A brief review of the "woolly rhino" Coelodonta antiquitatis (BLUMENBACH, 1799) (Mammalia, Rhinocerotidae) in Italy (Southern Europe). In: V. M. Podobina (Hrsg.): Evolution of life on the earth: Proceedings of of the IV international symposium, November 10-12, 2010. Tomsk, 2010, S. 529-533
- ↑ Tong HaoWen und Anne-Marie Moigne: Quaternary rhinoceros of China. Acta Anthropologica Sinica 19, 2000, S. 257-263
- ↑ Tong HaoWen: Rhinocerotids in China - systematics and material analysis. Geobios 34 (5), 2001, S. 585-591
- ↑ a b Cajus G. Diedrich: Eingeschleppte und benagte Knochenreste von Coelodonta antiquitatis (BLUMENBACH 1807) aus dem oberpleistozänen Fleckenhyänenhorst Perick-Höhlen im Nordsauerland und Beitrag zur Taphonomie von Wollnashornkadavern in Nordwest-Deutschland. Mitteilungen des Verbandes deutscher Höhlen- u. Karstforscher 54 (4), 2008, S. 100-117
- ↑ a b c П. А. Лазарев, С. Е. Григорьев и В. В. Плотинков: Шерстистые носороги Якутии. In: V. M. Podobina (Hrsg.): Evolution of life on the earth: Proceedings of of the IV international symposium, November 10-12, 2010. Tomsk, 2010, S. 555-558
- ↑ a b c Gennady G. Boeskorov, P. A. Lazarev, N. T. Bakulina, M. V. Shchelchkova, S. P. Davydov und N. G. Solomonov: Preliminary studie of woolly rhinoceros from the lower reaches of the Kolyma river. Doklady Biological Sciences 424, 2009, S. 53-56
- ↑ a b c Irina V. Kirillova und Fedor K. Shidlovskiy: Estimation of individual age and season of death in woolly rhinoceros, Coelodonta antiquitatis (Blumenbach, 1799), from Sakha-Yakutia, Russia. Quaternary Science Reviews 29, 2010, S. 3106-3114
- ↑ a b c Нина В. Гарүтт: К истории изучения шерстистого носорога Coelodonata antiquitatis (BLUMENBACH, 1799).GEOS 2000, S. 22-33
- ↑ Maciej J. Kotarba, Marek Dzieniewicz, Włodzimierz J. Mościcki und enryk Sechman: Unique Quaternary environment for discoveries of woolly rhinoceroses in Starunia, fore-Carpathian region, Ukraine: Geochemical and geoelectric studies. Geology 36 (7), 2008, S. 567-570
- ↑ Mikael Fortelius (coordinator). Neogene of the Old World Database of Fossil Mammals (NOW). University of Helsinki, 2003 http://www.helsinki.fi/science/now/
- ↑ a b c d e f g h i j Jan van der Made: The rhinos from the Middle Pleistocene of Neumark-Nord (Saxony-Anhalt). In: Dietrich Mania u. a. (Hrsg.): Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseums für Vorgeschichte 62. Halle/Saale 2010, S. 433–527
- ↑ Jan van der Made und René Grube: The rhinoceroses from Neumark-Nord and their nutrition. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 382–394
- ↑ a b c Nina V. Garutt: Dental ontogeny of the wolly rhino Coelodonta antiquitatis (BLUMENBACH, 1799). Cranium 11, 1994, S. 37-48
- ↑ a b c d e f g Mikael Fortelius: The morphology and paleobiological significance of the horns of Coelodonta antiquitatis (Mammalia: Rhinocerotidae). Journal of Vertebrate Paleontology 3 (2), 1983, S. 125-135
- ↑ a b c Nina Garutt: Neue Angaben über das Horn des Fellnashorns Coelodonta antiquitatis. Deinseas 4, 1998, S. 25-39
- ↑ a b Alexei V. Tiunov uand Irina V. Kirillova:Stable isotope (13C/12C and 15N/14N) composition of the woolly rhinoceros Coelodonta antiquitatis horn suggests seasonal changes in the diet. Rapid Communications Mass Spectrometry 24, 2010, S. 3146-3150
- ↑ О. Ф. Чернова и И. В. Кириллова: Новые данные о морфологии рога шерстистого носорога (Coelodonta antiquitatis BLUMENBACH, 1799). Труды Зоологического института РАН 314 (3), 2010, S. 333-342
- ↑ Irina Kirillova, Fedor Shidlovski und Olga Chernova: New data on woolly rhinoceros (Coelodonta antiquitatis Blum) horns. In: Abstraktband: Quaternary stratigraphy and paleontology of the Southern Russia: connections between Europe, Africa and Asia. 2010 annual meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26, 2010. (Rostov-am-Don) 2010, S. 70-71
- ↑ a b c d e f Claude Guérin: Coelodonta antiquitatis praecursor (Rhinocerotidae) du Pléistocène moyen final de l’aven de Romain-la-Roche (Doubs, France). Revue de Paléobiologie 29, 2010, S. 697-746
- ↑ a b c Nina Garutt: Traumatic skull damages in the woolly rhinoceros, Coelodonta antiquitatis BLUMENBACH, 1799. Cranium 14, 1997, S. 37.46
- ↑ Emmanuel M. E. Billia und Svetlana M. Graovac: Amelogenesis Imperfecta on a Deciduous Molar of Coelodonta antiquitatis (Blumenbach) (Mammalia, Perissodactyla, Rhinocerotidae) from Grotta di Fumane (Verona, Northern Italy): A Rare Case Report. iIn: J. T. Mayhall und T. Heikkinen (Hrsg.): Proceedings of the XI International Symposium on “Dental Morphology” – Oulu, Finland, August 26-30 1998. 1999, S. 179-186
- ↑ Bodil Bratlund: Comments on a cut-marked woolly rhino mandible from Zwoleń. In: Romuald Schild (Hrsg.): The killingfields of Zwoleń - A Middle Palaeolithic kill-butchery site in Central Poland. Warschau, 2005, S. 217-221
- ↑ Sabine Gaudzinski: Ein mittelpaläolithisches Rentierlager bei Salzgitter-Lebenstedt. In: Manfred Boetzkes, Ingeborg Schweitzer und Jürgen Vespermann (Hrsg.): EisZeit - Das große Abenteuer der Naturbeherrschung. Begleitbuch zur gleichnamigen Ausstellung. Hildesheim, 1999, S. 121-136
- ↑ Willfried Baumann und Dietrich Mania: Die paläolithischen Neufunde von Markkleeberg bei Leipzig (mit Beiträgen von L. Eißmann und V. Toepfer). Veröffentlichungen des Landesmuseums für Vorgeschichte Dresden 16, Berlin, 1983
- ↑ Doris Döppes: Die jungpleistozäne Säugetierfauna der Gudenushöhle (Niederösterreich). Wissenschaftliche Mitteilungen des Niederösterreichischen Landesmuseums 10, 1997, S. 17-32
- ↑ Dietrich Mania und V. Toepfer: Königsaue. Gliederung, Ökologie und mittelpaläolithische Funde der letzten Eiszeit. Veröffentlichungen des Landesmuseums für Vorgeschichte Halle/Saale 26, Berlin, 1973
- ↑ Sabine Gaudzinski: The faunal record of the Lower and Middle Palaeolithic of Europe: remarks on human interference. In: Wil Roebroeks und Clive Gamble (Hrsg.): The Middle Palaeolithic occupation of Europe. Leiden, 1999, S. 215-234
- ↑ Alicja Lasota-Moskalewska, Anna Gręzak und Johanna Piątkowska-Małecka: Anthropological traces on animal bones from Zwoleń. In: Romuald Schild (Hrsg.): The killingfields of Zwoleń - A Middle Palaeolithic kill-butchery site in Central Poland. Warschau, 2005, S. 207-216
- ↑ a b c Gerhard Bosinski: Die große Zeit der Eiszeitjäger. Europa zwischen 40.000 und 10.000 v. Chr. Jahrbuch des Römisch-Germanischen Zentralmuseums Mainz 34, 1987, S. 3-139
- ↑ Laura Niven: From carcass to cave: Large mammal exploitation during the Aurignacian at Vogelherd, Germany. Journal of Human Evolution 53 ,2007, S. 362e382
- ↑ Jean-Marie Chauvet, Éliette Brunel Deschamps und Christian Hillaire: Grotte Chauvet bei Vallon -Pont-d'Arc. Altsteinzeitliche Höhlenkunst im Tal der Ardèche Sigmaringen, 1995
- ↑ Annette Laming: Lascaux - Am Ursprung der Kunst. Dresden, 1959
- ↑ Michel Lorblanchet: Höhlenmalerei. Ein Handbuch. Sigmaringen, 1997
- ↑ Gerhard Bosinski: Die Kunst der Eiszeit in Deutschland und der Schweiz. Kataloge Vor- und Frühgeschichtlicher Altertümer 20, Bonn, 1982
- ↑ a b c Tao Deng: Comparison between woolly rhino forelimbs from Longdan, Northwestern China and Tologoi, Transbaikalian region. Quaternary International 179, 2008, S. 196-207
- ↑ Esperanza Cerdeño: Diversity and evolutionary trends of the the family Rhinocerotidae (Perissodactyla). Palaeo 141, 1998, S. 13-34
- ↑ Ralf-Dietrich Kahlke, Nuria García, Dimitris S. Kostopoulos, Frédéric Lacombat, Adrian M. Lister, Paul P.A. Mazza, Nikolai Spassov und, Vadim V. Titov: Western Palaearctic palaeoenvironmental conditions during the Early and early Middle Pleistocene inferred from large mammal communities, and implications for hominin dispersal in Europe. Quaternary Science Reviews, 2010, S. 1–28
- ↑ Roger M. Jacobi, James Rose, Alison MacLeod und Thomas F.G. Higham: Revised radiocarbon ages on woolly rhinoceros (Coelodonta antiquitatis) from western central Scotland: significance for timing the extinctionof woolly rhinoceros in Britain and the onset of the LGM in central Scotland. Quaternary Science Reviews 28, 2009, S. 2551–2556
- ↑ L. A. Orlova, S. K. Vasilev, Yaroslav V. Kuzmin und P. A. Kosintsev: New Data on the Time and Place of Extinction of the Woolly Rhinoceros Coelodonta antiquitatis Blumenbach, 1799. Doklady Biological Sciences, 2008, 423, S. 403–405
- ↑ a b Yaroslav V. Kuzmin: Extinction of the woolly mammoth (Mammuthus primigenius) and woolly rhinoceros (Coelodonta antiquitatis) in Eurasia: Review of chronological and environmental issues. Boreas 39, 2010, S. 247-261
- ↑ Tong Haowen und Liu Jinyi: The Pleistocene-Holocene extinction of mammals in China. In: Dong Wei (Hrsg.): Proceedings of the Ninth Annual Symposium of the Chinese Society of Vertebrate Paleontology. Peking, 2004, S. 111-119
- ↑ a b Tong HaoWen: Palaeoenvironmental significance of Coelodonta in different fossile assemblages. Acta Anthropologica Sinica 23, 2004, S. 306-314
- ↑ Walter Steiner: Der Travertin von Ehringsdorf und seine Fossilien. Lutherstadt Wittenberg 1981
- ↑ Dietrich Mania und Dieter-Hans Mai: Zum Klimacharakter der Warmzeit von Neumark-Nord 1. In: Harald Meller (Hrsg.): Elefantenreich – Eine Fossilwelt in Europa. Halle/Saale 2010, S. 174-185
- ↑ Anthony J. Stuart: The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe. Quaternary International 126-128, 2005, S. 171-177
- ↑ a b Claude Guérin: La famille des Rhinocerotidae (Mammalia, Perissodactyla): systématique, histoire, évolution, paléoécologie. Cranium 6, 1989, S. 3-14
- ↑ Tong HaoWen und Claude Guérin: Early Pleistocene Dicerorhinus sumatrensis remains from the Liucheng Gigantopithecus Cave, Guangxi, China. Geobios 42, 2009, S. 525-539
- ↑ Ludovic Orlando, Jennifer A. Leonard, Aurélie Thenot, Vincent Laudet, Claude Guerin, und Catherine Hänni: Ancient DNA analysis reveals woolly rhino evolutionary relationships. Molecular Phylogenetics and Evolution 28, 2003, S. 485–499
- ↑ Christelle Tougard, Thomas Delefosse, Catherine Hänni und Claudine Montgelard: Phylogenetic Relationships of the Five Extant Rhinoceros Species (Rhinocerotidae, Perissodactyla) Based on Mitochondrial Cytochrome b and 12S rRNA Genes. Molecular Phylogenetics and Evolution 19, 2001, S. 34–44
- ↑ a b c Н. Н. Ќаландадзе, А. В. Шаповалов и Є. М. Тесакова: К вопросам номенклатурам шерстистого носорога Coelodonta antiquitatis (BLUMENBACH, 1799), In: М. А. Шишкина и В. П. Твердохлебова (Hrsg.): Исследования по палеонтологии и биостратиграфии древних континентальных отложей. Памяти профессора В. Г. Очева. Saratow, 2009, S. 98-111
- ↑ a b Alexander Gehler, Mike Reich, Dick Mol und Hans van der Plicht: The type material of Coelodonta antiquitatis (Blumenbach) (Mammalia: Perissodactyla: Rhinocerotidae). Posterpräsentation während des 4th International Mammoth Conference 18-22 June, 2007, Jakutsk [1]
- ↑ Hans-Dietrich Kahlke: Das Eiszeitalter. Leipzig, Jena, Berlin, 1981
Kategorien:- Unpaarhufer
- Ausgestorbenes Säugetier
- Fauna des Pleistozäns














Wikimedia Foundation.