- Archaebakterium
-
Archaeen 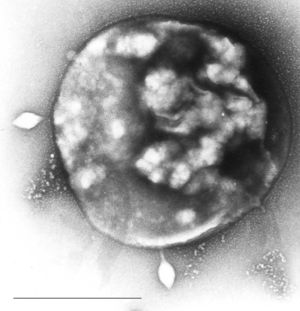
Archaeon des Genus Sulfolobus, infiziert mit dem Sulfolobus-Virus STSV1. Maßstab = 1 μm.
Systematik Klassifikation: Lebewesen Domäne: Archaeen Wissenschaftlicher Name Archaea Otto Kandler & Mark L. Wheelis Abteilungen Archaeen (Archaea, Singular: Archaeon; von griech. ἀρχαῖος archaĩos „uralt“, „ursprünglich“), früher auch Archaebakterien oder Urbakterien genannt, bilden neben den Bakterien (Bacteria) und den Eukaryoten (Eukaryota) eine der drei Domänen, in die alle zellulären Lebewesen eingeteilt werden.
Inhaltsverzeichnis
Beschreibung
Archaeen sind einzellige Organismen mit einem meist in sich geschlossenen DNA-Molekül (auch als Chromosom bezeichnet), das in einem kleinen Volumen angeordnet ist und in dieser Form als Kernäquivalent bezeichnet wird. Sie gehören also zu den Prokaryoten und besitzen weder ein Cytoskelett noch Zellorganellen.
Die separate Stellung der Archaeen als eigenständige Domäne ist durch deutliche Unterschiede in der Sequenz der in den Ribosomen enthaltenen RNA, und zwar der kleinen ribosomalen Untereinheit (16S rRNA) und durch weitere genetische, physiologische, strukturelle und biochemische Eigenschaften begründet.
Entdeckt und beschrieben wurden die Archaebakterien Ende der 1970er Jahre von den US-amerikanischen Mikrobiologen Carl Woese und George Fox. In der Sequenz der ribosomalen RNA entdeckten die Forscher auffällige Unterschiede zu den anderen Prokaryoten, dem Reich der Eubakterien; auch die Struktur der Zellen und deren Eigenheiten im Stoffwechsel ließen auf eine separate Gruppe von Prokaryoten schließen. Diese Ergebnisse wurden in den folgenden Jahren bestätigt und weitreichende Fortschritte in der molekularen Biologie machten eine generelle Änderung der Taxonomie notwendig: Eubakterien und Archaebakterien wurden in Bacteria und Archaea umbenannt und 1990 als eigenständige Domänen neben der Domäne der Eukarya beschrieben (Drei-Domänen-System).
Viele kultivierte Arten der Archaeen sind an extreme Milieubedingungen angepasst. So gibt es Arten, die bevorzugt bei Temperaturen von über 80 Grad Celsius wachsen (hyper-thermophil), andere leben in hoch konzentrierten Salzlösungen (halophil) oder in stark saurem Milieu (pH-Wert bis 0; acidophil) bzw. stark basischem Milieu (pH-Wert bis >10; alkaliphil). Picrophilus fumarii z.B. hat ein Optimum von pH 0,7 und kann sogar bei einem pH von -0,6 noch überleben.
Archaeen sind in der Forschung von Interesse, da in ihnen vielleicht Merkmale des frühen Lebens auf der Erde erhalten geblieben sind. Aber auch ihr außergewöhnlicher Stoffwechsel ist von Interesse, zum Beispiel die Fähigkeit, bei 110 °C zu wachsen. Auch im Hinblick auf Anwendungen ist der ungewöhnliche Stoffwechsel interessant, beispielsweise um Archaeen bei der Boden- und Gewässersanierung einzusetzen.
Bislang sind keine Krankheitserreger aus der Gruppe der Archaeen bekannt.
Eigenschaften
In vielen molekularbiologischen Eigenschaften sind die Archaeen den Eukaryoten ähnlicher als den Bakterien. Dennoch besitzen sie typisch bakterielle Eigenschaften, z. B. die Zellgröße, das Fehlen eines Zellkerns, die Art der Zellteilung, sie besitzen ein in sich geschlossenes DNA-Molekül, ebenfalls verhältnismäßig einfach aufgebaute Fortbewegungsorgane (Geißeln) und wie die Bakterien Ribosomen mit dem Sedimentationskoeffizienten 70S (allerdings sind die archaeellen Ribosomen komplexer in ihrer Struktur). Die Gene beider Domänen sind in sogenannten Operons organisiert. Archaeen können auch Plasmide tragen, wie beispielsweise ein Archaeon des Genus Sulfolobus.[1]
Die zentralen molekularen Prozesse, zum Beispiel Translation und Transkription, sind dagegen denjenigen der Eukaryoten recht ähnlich: Archaeen benutzen ähnliche, aus mehreren Proteinuntereinheiten zusammengesetzte RNA-Polymerasen (Rifampicin und Streptolydigin-resistent), bei der Translation kommen sehr ähnliche Initiations- und Elongationsfaktoren vor, der Beginn der Transkription wird durch eine so genannte TATA-Box markiert.
Die Archaeen besitzen jedoch auch viele einzigartige Eigenschaften, besonders der Aufbau der Zellwand zeigt deutliche Unterschiede zu den anderen Domänen: Die archaeellen Zellwände enthalten kein Murein (Peptidoglykan) und sind generell sehr vielfältig in ihrem Aufbau: Manchen Archaeen fehlt eine Zellwand völlig (Thermoplasma), andere besitzen hoch komplexe, aus vielen Schichten bestehende Zellwände (Methanospirillum). Aufgrund des anderen Aufbaus sind Archaeen generell gegen Zellwandantibiotika resistent. Auch die Zusammensetzung der archaeellen Plasmamembran unterscheidet sich: In Bakterien und Eukaryoten sind Fettsäuren über eine Ester-Bindung an die Glycerol-Moleküle gebunden, bei Archaeen findet man Glycerol-Diether oder sogar Glycerol-Tetraether (einschichtige Membran, Monolayer) und verzweigte Isopren-Einheiten statt einfacher Fettsäuren. Hyperthermophile Archaeen besitzen häufig stabilere, einschichtige Membranen (Glycerol-Tetraether).
Habitate
Die meisten der kultivierten Archaeen sind Extremophile, d. h., sie sind besonders an extreme Biotope angepasst. Viele Vertreter besitzen die Fähigkeit, bei sehr hohen Temperaturen (d. h. über 80 °C, Hyperthermophile), sehr niedrigen und hohen pH-Werten (Acidophile bzw. Alkaliphile), hohen Salzkonzentrationen (Halophile) oder hohen Drücken (Barophilie) zu leben. Hyperthermophile Archaeen findet man häufig in marinen und terrestrischen vulkanischen Gebieten (Black Smoker, Geysire, Solfatarenfelder); viele dieser Archaeen hat man z. B. aus vulkanisch geprägten Habitaten des Yellowstone National Parks isoliert. Halophile gedeihen gut in Umgebungen mit hohem Salzgehalt, z. B. im Toten Meer oder auch in natürlich vorkommenden marinen Brines. Auch methanogene Archaeen sind in gewisser Weise „extrem“: Sie wachsen ausschließlich unter anoxischen Bedingungen und benötigen häufig molekularen Wasserstoff für ihren Stoffwechsel. Sie sind relativ weit verbreitet: in Süßwasser, Meer, Boden, aber auch als Symbionten im Darmtrakt von Tieren und Menschen.
Aufgrund dieser „Extremophilie“ hat man die ökologische Bedeutung der Archaeen bislang als relativ gering eingeschätzt. Doch in den letzten Jahren hat man durch Einsatz neuer molekularer Methoden erkannt, dass Archaeen zu großen Anteilen im (verhältnismäßig kalten) Meerwasser, aber auch in Böden und Süßwasser-Biotopen vorkommen. In bestimmten ozeanischen Bereichen machen z. B. Crenarchaeota bis zu 90 % der vorhandenen Lebewesen aus. Insgesamt schätzt man, dass in den Ozeanen etwa 1,3 × 1028 Archaeen und 3,1 × 1028 Bakterien vorkommen. Die Mehrzahl der isolierten (d. h. als Reinkultur im Labor verfügbaren) Archaeen ist allerdings nach wie vor „extremophil“ und erst in wenigen Fällen ist eine Kultivierung von kälteliebenden Archaeen gelungen. Durch die kultivierungsunabhängigen molekularbiologischen Untersuchungen ist allerdings klar geworden, dass die Archaeen eine entscheidende Rolle im Ökosystem der Erde (Stickstoff-, Kohlenstoff-, Schwefelkreislauf) spielen.
Stoffwechsel
Viele Archaeen sind autotroph, d. h. sie gewinnen den Kohlenstoff zum Aufbau ihrer Körperbestandteile ausschließlich durch Assimilation von Kohlendioxid. Auch Heterotrophie, die Gewinnung des Kohlenstoffs aus organischen Verbindungen, ist weit verbreitet.
Die Mehrzahl der bisher kultivierten Archaeen zeichnet sich durch einen anaeroben Stoffwechsel aus; häufig ist für diese Archaeen Sauerstoff sogar toxisch.
Eine Besonderheit archaeellen Stoffwechsels ist die Methanogenese, die ausschließlich von Methan- produzierenden Archaeen, den sogenannten Methanogenen durchgeführt werden kann. Sie besitzen eine Reihe einzigartiger Cofaktoren, wie z. B. Coenzym F420 oder Methanofuran.
Hyperthermophile Archaeen sind in den meisten Fällen Anaerobier, der energiegewinnende Stoffwechsel ist entweder chemoorganotroph oder chemolithotroph (die Energie wird aus chemischen Umsetzungen organischer bzw. anorganischer Verbindungen gewonnen). Schwefelverbindungen spielen hierbei oft eine große Rolle: Während des Stoffwechsels wird der Schwefel reduziert und dabei Stoffwechselenergie frei.
Halophile Archaeen sind meist aerob-chemoorganotroph, sie gewinnen ihre Energie aus chemischen Umsetzungen von organischen Verbindungen. Unter anoxischen Bedingungen, oder bei Nährstoffmangel, sind viele extrem Halophile sogar zur Nutzung von Lichtenergie fähig: Das Protein Bacteriorhodopsin absorbiert Licht und katalysiert den Protonentransfer durch die Cytoplasmamembran, der dadurch erzeugte elektrochemische Gradient treibt die ATPase und damit die ATP-Synthese an.
Morphologie
Wie die Bakterien sind auch die Archaeen in ihrer Form äußerst divers. Die Größen bzw. Längen der archaeellen Zellen liegen etwa zwischen 0,4 („Nanoarchaeum equitans“) bis zu 100 µm (Methanospirillum hungatei), durchschnittlich sind die Zellen etwa 1 µm groß. Die Zellen zeigen verschiedenste Formen, z. B.: Kokken (z. B. Methanococcus jannaschii), Stäbchen (Thermoproteus neutrophilus), Spirillen-förmig (Methanospirillum hungatei), gelappte Kokken (Archaeoglobus fulgidus), Scheiben (Thermodiscus maritimus), lange Filamente (Thermofilum pendens) oder sogar quadratisch (Haloquadratum walsbyi). Sie besitzen oft Geißeln (Flagellen) zur Fortbewegung, oder auch fadenartige Anhängsel (Pili) zur Anheftung an Oberflächen.
Systematik
Die Platzierung der Archaeen im System der Taxonomie ist nicht ganz unumstritten. Anfangs nur durch Aussehen und Physiologie klassifiziert, wird heute aufgrund neuer Möglichkeiten allgemein die Einteilung mittels phylogenetischer Analyse akzeptiert, wie es Carl Woese (1977,1990) vorgeschlagen hat.[2][3]
Für die Beschreibung von Genus und Art gibt es eine festgelegte Prozedur.[4] Durch Publikation oder Validierung im International Journal of Systematic and Evolutionary Microbiology (IJSEM)[5] sind Gattung und Art festgelegt. Höhere Taxa können hier auch beschrieben werden. Der aktuelle Stand kann in der List of Prokaryotic names with Standing in Nomenclature (LPSN),[6] gepflegt durch Jean Euzéby, eingesehen werden. Dies entspricht dem internationalen Code der Nomenklatur von Bakterien (ICNB).[7] Taxa, die diesem Standard nicht entsprechen, werden in Anführungszeichen dargestellt.
Darüber hinaus wurde die globale Einteilung innerhalb der Archaeen und Bakterien reformiert, mittels phylogenetischer Analyse des 16S rRNS Gens[8]. Eine aktuelle Zusammenstellung der Taxa aus dieser und zahlreichen weiterführenden Publikationen erscheint in Bergey's Manual,[9] und in Taxonomic Outline of the Bacteria and Archaea[10], wobei mittlerweile zusätzlich zum 16S-rRNS-Gen teilweise weitere phylogenetische Markergene hinzugezogen werden. Einige dieser Taxa haben ihre Berechtigung, sind aber bis heute nicht valide publiziert oder werden generell nicht vom ICNB erfasst. Solche Taxa werden in Anführungszeichen dargestellt.
Die hier wiedergegebene Systematik enthält die Taxa von Phylum bis Familie. Bei manchen Taxa gibt es widersprüchliche Einträge. Diese wurden anhand von Originalliteratur und einer phylogenetischen Analyse auf Stichhaltigkeit geprüft.[11][12][13]
Vor einigen Jahren wurde die Beschreibung der zusätzlichen Phyla „Korarchaeota“ und „Nanoarchaeota“ veröffentlicht. Ein Vertreter des vorgeschlagenen Phylums „Nanoarchaeota“ konnte erfolgreich co-kultiviert[14] und sein Genom sequenziert werden,[15] das sogenannte „Nanoarchaeum equitans“. Vom vorgeschlagenen Phylum „Korarchaeota“, zunächst anhand seiner 16S rRNA-Gen-Basensequenzen in Proben heißer Quellen nachgewiesen,[16] gibt es Anreicherungskulturen. Daraus konnte nun die komplette Basensequenz des Genoms veröffentlicht werden,[17] mit dem informellen Namen „Candidatus Korarchaeum cryptofilum“ versehen. Ohne isolierte Stämme haben die Vertreter dieser Phyla nach den derzeitigen Regeln des ICSB keinen validierten Platz in der Taxonomie,[18] stellen jedoch zwei von vier bekannten Phyla der Archaeen dar.
Phylum „Crenarchaeota“
- Klasse Thermoprotei
- Ordnung „Caldisphaerales“
- Familie „Caldisphaeraceae“
- Ordnung Cenarchaeales
- Familie „Cenarchaeaceae“
- Ordnung Desulfurococcales
- Familie Desulfurococcaceae
- Familie Pyrodictiaceae
- Ordnung Sulfolobales
- Familie Sulfolobaceae
- Ordnung Thermoproteales
- Familie Thermoproteaceae
- Familie Thermofilaceae
- Ordnung „Caldisphaerales“
Phylum „Euryarchaeota“
- Klasse Archaeoglobi
- Ordnung Archaeoglobales
- Familie Archaeoglobaceae
- Ordnung Archaeoglobales
- Klasse Halobacteria
- Ordnung Halobacteriales
- Familie Halobacteriaceae
- Ordnung Halobacteriales
- Klasse Methanobacteria
- Ordnung Methanobacteriales
- Familie Methanobacteriaceae
- Familie Methanothermaceae
- Ordnung Methanobacteriales
- Klasse Methanococci
- Ordnung Methanococcales
- Familie Methanocaldococcaceae
- Familie Methanococcaceae
- Ordnung Methanococcales
- Klasse „Methanomicrobia“
- Ordnung Methanocellales
- Familie Methanocellaceae
- Ordnung Methanomicrobiales
- Familie Methanocorpusculaceae
- Familie Methanomicrobiaceae
- Familie Methanospirillaceae
- Ordnung Methanosarcinales
- Familie Methanosaetaceae
- Familie Methanosarcinaceae
- Familie Methermicoccaceae
- Ordnung Methanocellales
- Klasse Methanopyri
- Ordnung Methanopyrales
- Familie Methanopyraceae
- Ordnung Methanopyrales
- Klasse Thermococci
- Ordnung Thermococcales
- Familie Thermococcaceae
- Ordnung Thermococcales
- Klasse Thermoplasmata
- Ordnung Thermoplasmatales
- Familie Ferroplasmataceae
- Familie Picrophilaceae
- Familie Thermoplasmataceae
- Ordnung Thermoplasmatales
Humanpathogene Archaea?
Archaea wurden beim Menschen im Darm (Colon), im Mund (Zahnflora) und in der Vagina nachgewiesen. Hierbei treten vor allem Archaea auf, die der Gattung Methanobrevibacter zugehören, im Speziellen Methanobrevibacter smithii. Diese zählen zu den methanogenen Archaea. Jedoch kommen nicht bei allen Menschen M. smithii im Darm vor, bei Säuglingen unter zwei Jahren wurden bisher noch nie Archaea identifiziert.
Es wurde zwar eine Korrelation zwischen Erkrankung und Anzahl von Methanogenen nachgewiesen. Je mehr methanogene Archaea im (entzündeten) Zahnfleisch vorhanden waren, desto stärker war eine entsprechende Parodontitis ausgeprägt. Auch bei Patienten mit Darmkrebs bzw. Divertikulose war die Menge methanogener Archaea in jenen Bereichen erhöht. Dennoch tragen diese Archaea nur indirekt zur Erkrankung bei, indem sie das Wachstum echt pathogener Bakterien fördern.
Biotechnologisches Potential
Archaeelle Stoffwechselleistungen, Zellbestandteile oder Enzyme werden industriell angewendet. Vor allem die Extremophilen besitzen viele Eigenschaften, die sich biotechnologisch nutzen lassen. Einige Beispiele, die sich bereits in der Entwicklungsphase oder Anwendung befinden:
- Biotechnologie
- Biogas- Gewinnung
- Mikrobielle Erzlaugung („microbial ore leaching“ oder „bioleaching“): Bei diesem Prozess werden niederwertige, sulfidische Erze ausgelaugt (die Sulfidanteile werden mikrobiell zu Sulfat oxidiert und dadurch die Schwermetalle in einen löslichen Zustand überführt); dies wird zum Beispiel zur Gewinnung von Kupfer, Zink und Nickel angewendet.
- Medizin: Verwendung von Zellwandbestandteilen (sogenannte S-Layer) als Träger für Impfstoffe
- Nanotechnologie
- Verwendung der S-Layer für die Ultrafiltration
- Bacteriorhodopsin/ Purpurmembran phototropher halophiler Archaeen als Biosensoren
- Biologie
- Gewinnung hitzeresistenter Enzyme, z. B. α-Amylasen, proteolytische Enzyme, DNA-Polymerasen
- Gewinnung neuer Restriktionsenzyme
Einzelnachweise
- ↑ Schleper C, Holz I, Janekovic D, Murphy J, Zillig W (1995) A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating. J Bacteriol. 177: 4417–26. PMID 7635827.
- ↑ C. R. Woese, G. E. Fox: Phylogenetic structure of the prokaryotic domain: the primary kingdoms. In: Proc. Natl. Acad. Sci. U.S.A. Bd. 74, Heft 11, 1977, S. 5088-5090. PMID 270744
- ↑ C. R. Woese, O. Kandler, M. L. Wheelis: Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. In: Proc. Natl. Acad. Sci. U.S.A., Bd. 87, Heft 12, 1990, S. 4576-4579. PMID 2112744
- ↑ B. J. Tindall, P. Kämpfer, J. P. Euzéby, A. Oren: Valid publication of names of prokaryotes according to the rules of nomenclature: past history and current practice. In: Int J Syst Evol Microbiol. Bd. 56, Heft 11, 2006, S. 2715-2720. PMID 17082418. DOI: 10.1099/ijs.0.64780-0)
- ↑ International Journal of Systematic and Evolutionary Microbiology (IJSEM)
- ↑ J. P. Euzéby: List of bacterial names with standing in nomenclature: a folder available on the Internet. In: Int. J. Syst. Bacteriol. Bd. 47, 1997, S. 590-592. PMID 9103655 - List of Prokaryotic names with Standing in Nomenclature (LPSN)
- ↑ S. P. Lapage, P. H. Sneath, V. B. D. Skerman, E. F. Lessel, H. P. R. Seeliger, W. A. Clark: International Code of Nomenclature of Bacteria, 1990 Revision. (Bacteriological Code). ASM Press, Washington, D.C. 1992, ISBN 155581039X
- ↑ C. R. Woese, E. Stackebrandt, T. J. Macke, G. E. Fox: A phylogenetic definition of the major eubacterial taxa. In: Syst. Appl. Microbiol. Bd. 6, 1985, S. 143-151. PMID 11542017
- ↑ Bergey's Manual Trust, Department of Microbiology, 527 Biological, Sciences Building, University of Georgia, Athens, GA 30602-2605, USA - Bergey's Manual
- ↑ George M. Garrity, Timothy G. Lilburn, James R. Cole, Scott H. Harrison, Jean Euzéby, Brian J. Tindall: Taxonomic Outline of the Bacteria and Archaea. Release 7.7, March 6, 2007, Michigan State University Board of Trustees, www.taxonomicoutline.org
- ↑ W. Ludwig, O. Strunk, R. Westram, L. Richter, H. Meier, and 27 other authors: ARB: a software environment for sequence data. In: Nucleic Acids Res. Bd. 32, 2004, S. 1363-1371.
- ↑ E. Pruesse, C. Quast, K. Knittel, B. Fuchs, W. Ludwig, J. Peplies, F. O. Glöckner: SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. In: Nuc. Acids Res. Bd. 35, No. 21, 2007, S. 7188-7196.
- ↑ P. Yarza, M. Richter, J. Peplies, J. Euzeby, R. Amann, K. H. Schleifer, W. Ludwig, F. O. Glöckner, R. Rossello-Mora: The All-Species Living Tree Project: a 16S rRNA-based phylogenetic tree of all sequenced type strains. In: Syst. Appl. Microbiol. 2008, doi:10.1016/j.syapm.2008.07.001.
- ↑ Huber H, Hohn MJ, Rachel R, Fuchs T, Wimmer VC, Stetter KO (2002) A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. Nature. 417: 63-7. PMID 11986665.
- ↑ Waters E, Hohn MJ, Ahel I, Graham DE, Adams MD, Barnstead M, Beeson KY, Bibbs L, Bolanos R, Keller M, Kretz K, Lin X, Mathur E, Ni J, Podar M, Richardson T, Sutton GG, Simon M, Soll D, Stetter KO, Short JM, Noordewier M (2003) The genome of Nanoarchaeum equitans: insights into early archaeal evolution and derived parasitism. Proc Natl Acad Sci U S A. 100: 12984-8. PMID 14566062.
- ↑ Barns SM, Delwiche CF, Palmer JD, Pace NR (1996) Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences. Proc Natl Acad Sci U S A. 93: 9188-93. PMID 8799176.
- ↑ Elkins JG, Podar M, Graham DE, Makarova KS, Wolf Y, Randau L, Hedlund BP, Brochier-Armanet C, Kunin V, Anderson I, Lapidus A, Goltsman E, Barry K, Koonin EV, Hugenholtz P, Kyrpides N, Wanner G, Richardson P, Keller M, Stetter KO (2008) A korarchaeal genome reveals insights into the evolution of the Archaea. Proc Natl Acad Sci U S A. 105: 8102-7. PMID 18535141.
- ↑ P. De Vos, H. G. Trüper: Judicial Commission of the International Committee on Systematic Bacteriology. IXth International (IUMS) Congress of Bacteriology and Applied Microbiology. Minutes of the meetings, 14, 15 and 18 August 1999, Sydney, Australia. In: Int. J. Syst. Evol. Microbiol. Bd. 50, 2000, S. 2239–2244
Literatur
- Martin Dworkin, Stanley Falkow, Eugene Rosenberg, Karl-Heinz Schleifer, Erko Stackebrandt (Hrsg.) The Prokaryotes, A Handbook of the Biology of Bacteria. 7 Bände, 3. Auflage, Springer-Verlag, New York u. a. O., 2006, ISBN 0-387-30740-0
- Joseph W. Lengeler, Gerhart Drews, Hans G. Schlegel (Hrsg.) Biology of the Prokaryotes. Georg Thieme Verlag, Stuttgart 1999, ISBN 3-13-108411-1
- S. M. Barns, C. F. Delwiche, J. D. Palmers und N. R. Pace: Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences. In: Proc. Natl. Acad. Sci. USA. Bd. 93, 1996, S. 9188–9193
- B. M. Karner, E. F. DeLong, D. M. Karl: Archaeal dominance in the mesopelagic zone of the Pacific ocean. In: Nature. Bd. 409, 2001, S. 507–510
- H. König: Archaea. In: Encyclopedia of Life Sciences. John Wiley & Sons, Inc., 2003, (DOI: 10.1038/npg.els.0000443)
- Michael T. Madigan, John M. Martinko, Paul V. Dunlap, David P. Clark: Brock – Biology of Microorganisms, 12. Auflage. Pearson, San Francisco u. a. O. 2009, ISBN 0-321-53615-0
- George M. Garrity, Timothy G. Lilburn, James R. Cole, Scott H. Harrison, Jean Euzéby, Brian J. Tindall: Taxonomic Outline of the Bacteria and Archaea. Release 7.7, March 6, 2007, Michigan State University Board of Trustees, www.taxonomicoutline.org
- H. Huber, M. J. Hohn, R. Rachel, T. Fuchs, V. C. Wimmer, K. O. Stetter. A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont. In: Nature. Bd. 417(6884), 2002, S. 63–67
- E. Conway de Macario und Alberto JL. Macario (2009): Methanogenic archaea in health and disease: a novel paradigm of microbial pathogenesis. In: Int J Med Microbiol. 299(2); 99–108; PMID 18757236
- Sven P. Thoms: Ursprung des Lebens. Fischer, Frankfurt 2005, ISBN 3-596-16128-2
- C. R. Woese, G. E. Fox: Phylogenetic structure of the prokaryotic domain: The primary kingdoms. In: Proc. Natl. Acad. Sci. USA. Bd. 74, 1977, S. 5088–5090.
- C. R. Woese, O. Kandler, M. L. Wheelis: Towards a natural system of organisms: Proposal of the domains Archaea, Bacteria and Eucarya. In: Proc. Natl. Acad. Sci. USA. Bd. 87, 1990, S. 4576–4579.
Weblinks
- Klasse Thermoprotei
Wikimedia Foundation.