- Paarhufer
-
Paarhufer 
Giraffe (Giraffa camelopardalis)
Systematik Überklasse: Kiefermäuler (Gnathostomata) Reihe: Landwirbeltiere (Tetrapoda) Klasse: Säugetiere (Mammalia) Unterklasse: Höhere Säugetiere (Eutheria) Überordnung: Laurasiatheria Ordnung: Paarhufer Wissenschaftlicher Name Artiodactyla Owen, 1848 Unterordnungen - Schweineartige (Suina)
- Schwielensohler (Tylopoda)
- Wiederkäuer (Ruminantia)
- Flusspferde (Cetancodonta)
Die Paarhufer, auch Paarzehige Huftiere oder Paarzeher (Artiodactyla oder Paraxonia), sind in der klassischen Systematik eine Ordnung der Säugetiere (Mammalia). Es handelt sich um überwiegend pflanzenfressende Tiere, die im Gegensatz zu den Unpaarhufern meist durch eine gerade Anzahl von Zehen (zwei oder vier) charakterisiert sind. Zu dieser Gruppe zählen einige der wirtschaftlich bedeutendsten Säugetiergruppen wie Rinder, Schweine, Kamele, Ziegen und Schafe, aber auch andere bekannte Tiere wie Giraffen, Flusspferde, Hirsche oder Antilopen. Molekulare Untersuchungen haben das Ergebnis gebracht, dass die Paarhufer wahrscheinlich paraphyletisch in Bezug auf die Wale sind. Das heißt, dass einige Gruppen (insbesondere die Flusspferde) näher mit den Walen als mit den übrigen Vertretern dieser Ordnung verwandt sind. Moderne phylogenetische Systematiken fassen entsprechend Paarhufer und Wale zu einem gemeinsamen Taxon Cetartiodactyla zusammen. Die Paarhufer sind demnach ein „Formtaxon“ – eine Gruppe, die zwar keine systematische Einheit bildet, aber durch gemeinsame Merkmale charakterisiert wird.
Paarhufer lassen sich in vier Unterordnungen einteilen:
- Die Schweineartigen (Suina) bestehen aus den Echten oder Altweltlichen Schweinen und den Nabelschweinen oder Pekaris.
- Die Flusspferde gehören in eine eigene Unterordnung, die Cetancodonta.
- Die Kamele – einschließlich der neuweltlichen Arten – sind die einzigen rezenten Vertreter der Unterordnung der Schwielensohler (Tylopoda).
- Die Wiederkäuer (Ruminantia) bestehen unter anderem aus Giraffenartigen, Hirschen und Hornträgern, zu denen beispielsweise die Rinder, die Ziegenartigen und die als Gazellen und Antilopen bezeichneten Tiere zählen.
Inhaltsverzeichnis
Körperbau
Die Paarhufer haben einige gemeinsame Merkmale insbesondere im Bau der Gliedmaßen, die sich bei den heute lebenden (rezenten), hochgradig an das Wasserleben angepassten Walen nicht finden. Als gemeinsame abgeleitete Merkmale (Synapomorphien) der Paarhufer galten ein spezieller Bau des Sprungbeins mit zwei Gelenkrollen (Trochlea tali proximalis und distalis) und der verlängerte letzte untere Vormahlzahn. Allerdings hat die Entdeckung dieses speziellen Sprungbeins bei fossilen Walen im Jahr 2001 den Diskussionen über die Systematik neue Nahrung gegeben (siehe unten).
Allgemeiner Körperbau und Fell
 Die Hirschferkel sind die kleinsten Paarhufer.
Die Hirschferkel sind die kleinsten Paarhufer.
Die Paarhufer sind mittelgroße bis große Tiere, die sich vierfüßig (quadruped) fortbewegen. Es gibt zwei Gestalttypen, die sich deutlich im Körperbau unterscheiden. So zeichnen sich etwa Schweineartige und Flusspferde durch einem stämmigen Rumpf, kurze Beine und einen großen Kopf aus. Schwielensohler und Wiederkäuer weisen hingegen einen eher schlanken Körperbau und lange, dünne Beine auf. Die Größe der Paarhufer variiert beträchtlich. Als kleinste Vertreter gelten die Hirschferkel, die oft nur 45 Zentimeter Kopfrumpflänge und ein Gewicht von 1,5 Kilogramm erreichen. Die größten Vertreter sind – mit bis zu 5 Metern Kopfrumpflänge und 4,5 Tonnen Gewicht – das Flusspferd beziehungsweise – mit bis 5,5 Meter Höhe und 4,7 Meter Kopfrumpflänge – die Giraffe. Hinsichtlich der Größe herrscht bei fast allen Arten ein Sexualdimorphismus, das heißt, dass die Männchen durchweg größer und schwerer werden als die Weibchen. Geschlechtsunterschiede gibt es auch bei den Stirnwaffen, so haben bei Hirschen in der Regel nur die Männchen ein Geweih und die Hörner der Hornträger sind bei den Weibchen meist deutlich kleiner oder fehlen gänzlich. Auch hinsichtlich anderer Aspekte wie der der Bezahnung oder der Fellfärbung kann es Sexualdimorphismen geben. So etwa bei der Hirschziegenantilope, bei der das Fell der Männchen dunkelbraun, das der Weibchen hingegen ockerfarben ist.
Bis auf die nahezu unbehaarten Flusspferde sind alle Vertreter dieser Ordnung mit einem dichten Fell bedeckt, dessen Länge und Färbung je nach Lebensraum variieren. Arten in kühleren Regionen können einen Fellwechsel durchmachen. Es überwiegen unscheinbare, tarnende Fellfärbungen wie Gelb-, Grau-, Braun- oder Schwarztöne.
Gliedmaßen und Bewegungsapparat
Äußerer Bau der Gliedmaßen
Paarhufer tragen ihren Namen, weil sie meist eine gerade Anzahl von Zehen (zwei oder vier) haben – lediglich bei einigen Nabelschweinen ist es bei den Hinterbeinen zu einer Reduktion der Zehenanzahl auf drei gekommen. Die Mittelachse eines Beins liegt bei Paarhufern zwischen der dritten und vierten Zehe. Diese beiden Mittelzehen sind am besten ausgebildet. Die ursprünglich vorhandene erste Zehe fehlt bei heutigen Paarhufern immer, sie ist nur bei fossilen Vertretern belegt. Die zweite und fünfte Zehe sind unterschiedlich ausgebildet: Bei den Flusspferden sind sie nach vorne gerichtet und voll funktionsfähig. Bei den übrigen Paarhufern sind sie rückwärts angeordnet (und werden als „Afterzehe“ bezeichnet) oder vollständig reduziert. Bei den Schweineartigen und Hirschferkeln werden die Afterzehen bei weichem, schlammigen Untergrund noch mitbenutzt und vergrößern die Auftrittsfläche. In den meisten Fällen berühren sie den Boden jedoch nicht mehr. Bei manchen Gruppen wie bei den Kamelen und den Giraffenartigen ist die Rückbildung so weit fortgeschritten, dass die zweite und fünfte Zehe nicht einmal mehr als Rudimente vorhanden sind.
-

Bei Flusspferden sind alle vier Zehen nach vorne gerichtet.
-

Bei Schweinen und vielen anderen Paarhufern sind die zweite und fünfte Zehe als Afterklaue nach hinten gerichtet.
-

Bei Kamelen sind nur mehr zwei Zehen vorhanden, die Klauen sind zu Nägeln umgebildet.
Trotz der Bezeichnung Paar„hufer“ ist die Bezeichnung Huf für das Zehenendorgan genau genommen den Pferden vorbehalten, im Falle der Paarhufer spricht man korrekterweise von Klauen.[1] Diese Klauen sind aus Hornsubstanz gebildet und setzen sich ursprünglich aus drei Teilen zusammen, der Platte (oben und an den Seiten), der Sohle (unten) und dem Ballen (hinten), die jedoch in unterschiedlichem Ausmaß verwachsen sein können. Generell sind die Klauen der Vorderbeine breiter und stumpfer als die der Hinterbeine und klaffen stärker auseinander. Bis auf die Kamele sind alle Paarhufer Zehenspitzengänger, das heißt sie setzen nur die Spitze des vordersten Zehengliedes auf den Boden auf. Bei Kamelen sind die Horngebilde an den Füßen zu Nägeln reduziert, die Zehen ruhen auf einem elastischen Polster aus Bindegewebe, das eine breite Sohlenfläche bildet. Daher stammt auch die Bezeichnung der Unterordnung der Schwielensohler (Tylopoda), deren einzige rezente Vertreter die Kamele sind.
Bewegungsapparat
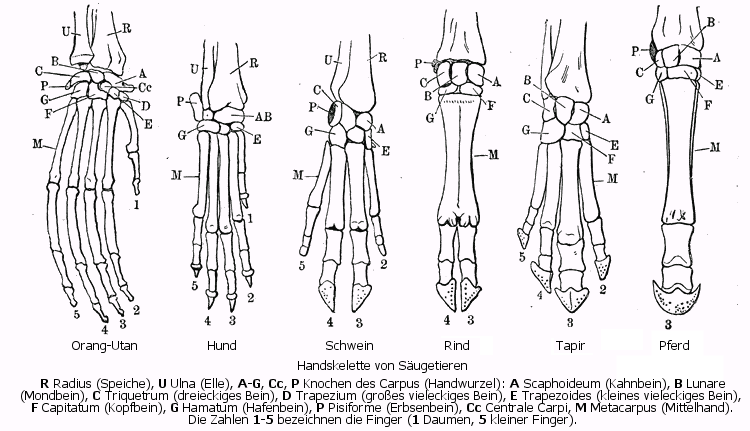
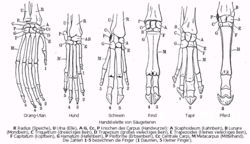 Handskelette verschiedener Säugetiere: Beim Schwein (3. von links) ist die Verkleinerung der 2. und 5. Zehe, beim Rind (4. von links) das Verschmelzen der Mittelhand zum Hauptmittelfußknochen gut zu sehen.
Handskelette verschiedener Säugetiere: Beim Schwein (3. von links) ist die Verkleinerung der 2. und 5. Zehe, beim Rind (4. von links) das Verschmelzen der Mittelhand zum Hauptmittelfußknochen gut zu sehen.Bei den Paarhufern herrscht eine Tendenz zur Verwachsung der Metapodien (Mittelhand- und Mittelfußknochen). Bei den Schweineartigen und Flusspferden sind sie noch getrennt und nur durch ein straffes Bindegewebe gekoppelt. Bei den Kamelen und Wiederkäuern sind 3. und 4. Metapodium zu einer Einheit, dem Hauptmittelfußknochen, verschmolzen, dessen Ursprung aus zwei Knochen oft durch Längsrinnen an der Vorder- und Hinterseite (Sulcus longitudinalis dorsalis und palmaris bzw. plantaris) noch sichtbar ist.[2]
Die Knochen des Stylopodiums (Oberarm- bzw. Oberschenkelknochen) und Zygopodiums (Elle und Speiche bzw. Schien- und Wadenbein) sind meist langgestreckt. Die Muskulatur der Gliedmaßen ist vorwiegend rumpfnah lokalisiert, was dafür sorgt, dass Paarhufer oft sehr schlanke Beine haben. Ein Schlüsselbein ist nie vorhanden, das Schulterblatt ist sehr beweglich, sein Vor- und Zurückschwingungen sorgt für zusätzliche Beweglichkeit bei schnellem Lauf.
Der spezielle Bau der Beine sorgt für eine steife Stellung der unteren Gliedmaßenglieder. Drehbewegungen der Beine sind kaum mehr möglich, die Unbeweglichkeit bewirkt aber eine höhere Stabilität bei schneller Fortbewegung. Zusätzlich haben viele kleinere Paarhufer einen sehr flexiblen Rumpf, dessen Biegsamkeit bei der Flucht zur Vergrößerung der Schrittlänge beiträgt. Neben dem Selektionsdruck zur Erlangung hoher Geschwindigkeiten bei der Flucht erhöht der spezialisierte Bau der Gliedmaßen auch die Energieersparnis bei langsamen Bewegungen, etwa bei der Nahrungsaufnahme.
Kopf und Zähne
Allgemeines
Viele Paarhufer haben einen verhältnismäßig großen Kopf. Der Gesichtsschädel ist langgestreckt und eher schmal, die Nasenbeine laufen nach vorn in einer oder zwei Spitzen aus. Im Hirnschädel ist das Stirnbein nach hinten vergrößert und verdrängt das Scheitelbein, das insbesondere bei den Wiederkäuern nur den Seitenteil des Schädeldachs bildet.
Die Lippen sind beweglich und stark muskularisiert. Bei einigen Gruppen wie den Schweineartigen, aber auch manchen Wiederkäuern wie der Saiga und den Dikdiks, ist es durch Verlängerung der Oberlippe zu einer Rüsselbildung gekommen.
Stirnwaffen

Vier Familien der Paarhufer, die Giraffenartigen, Hirsche, Gabelhornträger und Hornträger, haben Stirnwaffen; Die Gruppe der Stirnwaffenträger (Pecora) umfasst mit den Moschushirschen und Wasserrehen aber auch stirnwaffenlose Vertreter. Dabei handelt es sich meist um Auswüchse des Stirnbeins, die unterschiedlich gebaut sind. Die Giraffenartigen haben Knochenzapfen, die mit behaarter Haut umkleidet sind. Hirsche sind durch ein Geweih gekennzeichnet, das aus zapfenförmigen Knochengebilden („Rosenstöcken“) wächst. Es besteht aus Knochensubstanz und wird jedes Jahr nach der Paarungszeit abgestoßen und neu gebildet. Die Hörner der Hornträger bestehen im Gegensatz dazu aus Hornsubstanz und werden meist ein Leben lang beibehalten. Die Haut, die den Knochenzapfen bedeckt, scheidet Hornzellen ab, die schließlich eine harte Hornscheide bilden. Die ältesten Hornschichten werden dabei immer weiter Richtung Hornspitze verschoben. Mit Ausnahme der Vierhornantilope (und einzelner Haustierrassen) haben alle Hornträger zwei Hörner. Bei den Gabelhornträgern schließlich werden die Hörner ähnlich wie die der Hornträger gebildet, im Gegensatz dazu werden die Hornscheiden aber jährlich abgeworfen.
Die Stirnwaffen können dem Imponiergehabe, dem Kampf um das Paarungsvorrecht und der Verteidigung dienen. In fast allen Fällen sind sie geschlechtsdimorph ausgebildet, das heißt bei Männchen größer als bei Weibchen. Bei einigen Arten wie nahezu allen Hirschen, einigen Waldböcken und dem Okapi fehlt sie den Weibchen generell.
Zähne
Zahnformel I C P M 30–44 = 0–3 0–1 2–4 3 1–3 1 2–4 3 
Die Bezahnung der Paarhufer ist variabel, es lassen sich jedoch zwei Trends erkennen. Die Schweineartigen und Flusspferde haben relativ viele Zähne (bei einigen Echten Schweinen ist sogar die ursprüngliche Zahnzahl der Höheren Säugetiere (44) erhalten geblieben). Das Gebiss ist eher an einen quetschenden Kauvorgang angepasst, was der tendenziell allesfressenden Ernährung dieser Tiere entspricht. Bei den Kamelen und Wiederkäuern ist die Zahnzahl reduziert, es klafft oft eine als Diastema bezeichnete Lücke im Gebiss, und die Backenzähne sind auf eine mahlende Zerkleinerung der Pflanzennahrung ausgerichtet.
Die Schneidezähne sind oft reduziert, bei den Wiederkäuern fehlen sie am Oberkiefer vollständig, stattdessen pressen die unteren Schneidezähne gegen eine Dentalplatte. Die Eckzähne sind unterschiedlich ausgeprägt: Bei den Schweineartigen sind sie vergrößert und hauerartig entwickelt, sie dienen dem Graben im Erdreich und zur Verteidigung. Bei den Wiederkäuern ist der obere Eckzahn bei Männchen von Arten ohne Stirnwaffen (Hirschferkel, Moschushirsche, Wasserreh) vergrößert und wird als Waffe beim Kampf um das Paarungsvorrecht eingesetzt. Bei Arten mit Stirnwaffen fehlt der obere Eckzahn hingegen meist. Die unteren Eckzähne der Wiederkäuer gleichen den Schneidezähnen, sodass diese Tiere acht gleichförmige Zähne im Vorderteil des Unterkiefers haben.
Die Backenzähne der Schweineartigen sind niederkronig und mit wenigen Höckern versehen. Im Gegensatz dazu sind die der Kamele und Wiederkäuer hochkronig und die Höcker sind zu halbmondförmigen Schmelzleisten umgebildet (selenodont).
Sinne
Zur Wahrnehmung der Umwelt dient in erster Linie der Geruchssinn, der wie bei den meisten Säugetieren sehr gut entwickelt ist. Auch der Gehörsinn ist stark ausgeprägt, unterstützt wird dieser durch die bei vielen Arten vorhandenen beweglichen Ohrmuscheln. Im Gegensatz zu vielen anderen Säugern ist auch der Gesichtssinn zumindest bei Wiederkäuern und Kamelen ausgeprägt. Dabei ist vor allem das Bewegungssehen entwickelt, unbewegte Objekte werden eher nicht wahrgenommen. Analog zu vielen anderen Tieren, die ständig aufmerksam gegenüber Räubern sein müssen, sind die Augen seitlich am Kopf angebracht, was einem nahezu vollständigen Rundumblick und dem frühestmöglichen Erkennen von Bedrohungen dient.
Weichteilanatomie
Verdauungstrakt
 Echte Schweine haben im Gegensatz zu den anderen Paarhufern einen einfachen, sackförmigen Magen.
Echte Schweine haben im Gegensatz zu den anderen Paarhufern einen einfachen, sackförmigen Magen. Hirsche haben wie alle Wiederkäuer einen mehrkammerigen Magen, der der besseren Verwertung der Pflanzennahrung dient.
Hirsche haben wie alle Wiederkäuer einen mehrkammerigen Magen, der der besseren Verwertung der Pflanzennahrung dient.Als Anpassung an die schwer verdauliche Pflanzennahrung haben die Paarhufer einige Besonderheiten des Verdauungstraktes entwickelt, die vor allem bei den Wiederkäuern stark ausgeprägt sind. Im Bereich des Mundes sind oft zusätzliche Speicheldrüsen vorhanden und die Mundschleimhaut ist häufig stark verhornt, um Verletzungen durch harte Pflanzenteile zu vermeiden und den leichteren Transport der grob zerkauten Nahrung zu ermöglichen.
Im Bau des Magens zeigen die Paarhufer die wohl höchsten Spezialisierungen unter allen Säugetieren, wobei es mehrfach unabhängig voneinander zur Entwicklung mehrerer Magenabschnitte gekommen ist. Am einfachsten ist er bei den Echten Schweinen gebaut, die noch einen einfachen, sackförmigen Magen besitzen. Bei Nabelschweinen und Flusspferden kam es zur Entwicklung mehrerer Blindsäcke, in denen die Nahrung durch Mikroorganismen zersetzt wird. Der Magen der Kamele ist dreigegliedert und teilt sich in zwei Vormägen, die aber im Gegensatz zu denen der Wiederkäuer mit Drüsen ausgestattet sind, und dem eigentlichen oder Labmagen. Kamele können wiederkäuen, werden aber nicht zum Taxon der Wiederkäuer gerechnet.
Der Magen der Wiederkäuer schließlich teilt sich in drei oder vier Abschnitte: den Pansen (Rumen), den Netzmagen (Reticulum), den Blättermagen (Omasum) und den Labmagen (Abomasum). Den innerhalb der Wiederkäuer als besonders urtümlich eingestuften Hirschferkeln fehlt der Blättermagen, ansonsten zeigen alle Arten dieser Unterordnung den gleichen Bau und die gleiche Verdauungsweise. Die Vormägen sind drüsenfrei, hier wird die grob zerkaute Nahrung durch Mikroorganismen zersetzt, und in kleinen Portionen wieder in die Mundhöhle gefördert, wo sie wiedergekäut wird, bevor sie in den eigentlichen Magen kommt und weiter verdaut wird. (Ausführliches siehe im Artikel Wiederkäuer). Diese Verdauungsweise bietet zwei Vorteile: zum einen kann dabei die schwer verdauliche Pflanzennahrung bestmöglich aufgeschlüsselt und verwertet werden. Zum anderen wird die Dauer der eigentlichen Nahrungsaufnahme – zumal mit der für die Wahrnehmung der Umwelt ungünstigen Körperhaltung mit dem Kopf nahe beim Boden – verkürzt, was im Hinblick auf die Bedrohung durch Fressfeinde von Vorteil ist; das Wiederkäuen kann dann an geschützten Plätzen erfolgen.
Der Darm der Paarhufer ist wie bei vielen pflanzenfressenden Säugetieren generell sehr lang, der Dünndarm ist stark in Schleifen gelegt. Im Gegensatz zu den Unpaarhufern, bei denen die Fermentation erst im Darm stattfindet, sind Blind- und Grimmdarm einfacher gebaut und weniger voluminös.
Harn- und Geschlechtsapparat
Der Bau des Harn- und Geschlechtsapparates der Paarhufer zeigt ebenfalls einige Auffälligkeiten. Der Penis ist im Ruhezustand s-förmig gebogen und ruht in einer Hauttasche am Bauch. Er ist fibroelastisch, das heißt die Schwellkörper sind nur gering entwickelt und die Erektion bewirkt vor allem eine Streckung dieser Krümmung und damit eine Verlängerung, kaum aber eine Verdickung des Penis. Dieser Bau des Penis findet sich in ähnlicher Weise bei den Walen und stellt ein Anzeichen für die nahe Verwandtschaft dar. Die Hoden liegen im Hodensack und damit außerhalb der Bauchhöhle. Die Eierstöcke vieler Weibchen machen einen Abstieg (Descensus ovarii) durch – vergleichbar dem Hodenabstieg vieler männlicher Säuger – und liegen nahe dem Beckeneingang auf Höhe des vierten Lendenwirbels. Die Gebärmutter ist zweihörnig ausgebildet (Uterus bicornis).[3]
Sonstiges
 Bei diesen Japanischen Serauen sind die Präorbitaldrüsen (Drüsen vor den Augen) gut zu erkennen.
Bei diesen Japanischen Serauen sind die Präorbitaldrüsen (Drüsen vor den Augen) gut zu erkennen.Einige Paarhufer haben Besonderheiten im Kreislaufsystem entwickelt. Das Herz der Echten Schweine weist eine paarige Knorpeleinlagerung zwischen Vorhöfen und Kammern auf. Bei einigen Wiederkäuern sind zwei Herzknochen (Ossa cordis) ausgebildet, die die Aortenöffnung stabilisieren.
Die Anzahl der Milchdrüsen ist variabel und korreliert wie bei allen Säugetieren ungefähr mit der Wurfgröße. Ursprünglich waren vermutlich zwei Reihen von Zitzen vom Achsel- bis in den Leistenbereich vorhanden. Diese ursprüngliche Anordnung findet sich noch bei einigen Echten Schweinen, die auch die höchste Wurfgröße aller Paarhufer haben. In den meisten Fällen ist es jedoch zu einer Reduktion der Zitzenanzahl gekommen, die übrigen Paarhufer haben nur noch ein oder zwei Paar Zitzen. Diese bilden bei einigen Arten ein Euter in der Leistenregion.
Sekretdrüsen in der Haut sind bei nahezu allen Arten vorhanden und können an den unterschiedlichsten Stellen lokalisiert sein, etwa vor den Augen, hinter den Hörnern, am Nacken oder Rücken, an den Füßen oder in der Analregion.
Verbreitung und Lebensraum
Paarhufer sind mit Ausnahme des australisch-ozeanischen Raums, der Antarktis und vieler abgelegener Inseln weltweit verbreitet. Der Artenschwerpunkt liegt heute in Afrika und Asien. In Amerika ist die Gruppe relativ artenarm, insbesondere in Südamerika, wo nur Nabelschweine, Neuweltkamele (Lamas und Vikunjas) und Trughirsche vorkommen. Hier haben andere Gruppen wie die ausgestorbenen Südamerikanischen Huftiere und einige Nagetiere (zum Beispiel Capybaras, Pampashasen oder Agutis) ähnliche ökologische Nischen besetzt. Der Mensch hat verschiedene Paarhuferarten als Haus- oder Jagdtiere weltweit verbreitet, sodass diese Tiere heute fast überall zu finden sind, wo es Menschen gibt.
Paarhufer bewohnen nahezu alle Lebensräume, von tropischen Regenwäldern und Steppen bis Wüstengebiete und Hochgebirgsregionen. Die größte Artenvielfalt herrscht aber in offenen Habitaten wie Grasländern und lichten Wäldern. Diese Tiere sind ausgesprochene Bodenbewohner, nur wenige Arten führen eine semiaquatische (teilweise im Wasser stattfindende Lebensweise), etwa die Flusspferde. Einige Arten haben das Hochgebirge besiedelt und können ausgezeichnet klettern.
Lebensweise
Sozialverhalten und Aktivitätszeiten
 Paarhufer wie diese Impalas leben oft in Gruppen.
Paarhufer wie diese Impalas leben oft in Gruppen.Das Sozialverhalten der Paarhufer ist variabel. Generell herrscht aber eine Tendenz, sich zu größeren Gruppen zusammenzuschließen, es gibt aber auch einzelgängerisch oder dauerhaft in Paaren lebende Tiere (etwa bei den Hirschferkeln). Bei den in Gruppen lebenden Arten entwickelt sich oft eine Hierarchie, sowohl unter den Männchen als auch den Weibchen. Etliche Arten leben aber auch in Haremsgruppen, das heißt, dass ein einzelnes Männchen einige Weibchen und den gemeinsamen Nachwuchs um sich schart und keine Nebenbuhler duldet. Bei anderen Arten bilden die Weibchen und die Jungtiere während des größten Teils des Jahres eigene Gruppen, während die Männchen einzelgängerisch oder in Junggesellengruppen leben und nur zur Paarungszeit die Weibchengruppen aufsuchen. Bei vielen Paarhufern kommt es während der Paarungszeit zu erbitterten Kämpfen um das Paarungsvorrecht zwischen den Männchen, die mit den Stirnwaffen, den hauerartigen Eckzähnen oder auf andere Weise ausgetragen werden.
Viele Paarhufer sind territorial und markieren ihr Revier beispielsweise mit Drüsensekreten oder Urin. Neben ganzjährig standorttreuen Arten gibt es auch Tiere, die jahreszeitliche Wanderungen auf der Suche nach besseren Nahrungsplätzen unternehmen.
Über die Aktivitätszeiten lassen sich keine generellen Aussagen machen. Es gibt sowohl tag-, dämmerungs- und nachtaktive Vertreter als auch Arten, bei denen das Tag-Nacht-Schema je nach Jahreszeit oder Lebensraum variieren kann.
Ernährung
Die meisten Paarhufer sind Pflanzenfresser, deren Nahrungsspektrum je nach Art und Lebensraum variieren kann. Oft werden Gräser, Kräuter oder Blätter, aber auch andere Pflanzenteile wie Knollen oder Früchte verzehrt. Echte Schweine, Nabelschweine und in geringem Ausmaß auch Hirschferkel sind Allesfresser, die ihre Nahrung mit Insekten, Würmern und manchmal auch kleinen Wirbeltieren ergänzen. Die meisten Arten sind auf eine tägliche Wasseraufnahme angewiesen, einige in trockenen Habitaten lebende Arten können jedoch wochenlang ohne Trinken überleben – bekanntestes Beispiel sind die Kamele.
Fortpflanzung und Lebenserwartung
 Wie diese jungen Streifengnus kommen fast alle Paarhufer behaart und mit geöffneten Augen zur Welt und sind Nestflüchter.
Wie diese jungen Streifengnus kommen fast alle Paarhufer behaart und mit geöffneten Augen zur Welt und sind Nestflüchter.Generell herrscht bei den Paarhufern eine Tendenz zu langer Trächtigkeitsdauer, geringer Wurfgröße und hohem Entwicklungsgrad der Neugeborenen. Wie bei vielen anderen Säugetieren haben Arten in gemäßigten oder polaren Regionen eine feste Paarungssaison, während es bei Arten in tropischen Gebieten oft ganzjährig zur Fortpflanzung kommen kann. Entsprechend der Lebensweise überwiegt ein polygynes Paarungsverhalten, ein Männchen paart sich also oft mit mehreren Weibchen. Die Begattung erfolgt üblicherweise durch das säugertypische „Aufreiten“, nur bei den Kamelen wird sie im Liegen vollzogen.
Die Länge der Tragzeit variiert zwischen 4 und 5 Monaten bei Schweineartigen, Hirschferkeln und Moschushirschen, 6 bis 10 Monaten bei Flusspferden, Hirschen und Hornträgern, 10 bis 13 Monaten bei Kamelen und 14 bis 15 Monaten bei den Giraffenartigen. In der Regel kommen ein bis zwei Jungtiere zur Welt, bei einigen Schweinen können es aber bis zu zehn sein.
Die Neugeborenen aller Arten sind Nestflüchter und kommen mit geöffneten Augen und, mit Ausnahme der generell haarlosen Flusspferde, behaart zur Welt. Typisch für einige Paarhufer (etwa Schweine oder Hirsche) ist das gestreifte oder gepunktete Fellkleid der Jungtiere, das der Tarnung dient und sich im Aufwachsen verliert. Die Jungtiere einiger Arten verbringen ihre ersten Wochen mit der Mutter an einem geschützten Lagerort, andere können bald nach der Geburt laufen und der Herde binnen weniger Stunden oder Tage folgen.
Die Lebenserwartung beträgt in der Regel 20 bis 30 Jahre, wie bei vielen Säugetieren haben kleinere Arten eine oft kürzere Lebensspanne als große Arten. Am ältesten werden Tiere wie Flusspferde, Rinder und Kamele, die 40 bis 50 Jahre erreichen können.
Freßfeinde und Parasiten
Je nach Größe und Lebensraum haben Paarhufer unterschiedliche natürliche Feinde, oft sind es jedoch Raubtiere, etwa Katzen, Hunde oder Bären, die Jagd auf diese Tiere machen. Andere Fressfeinde sind beispielsweise Krokodile, große Greifvögel und bei kleinen Arten und Jungtieren auch Riesenschlangen.
Zu den bei Paarhufern parasitierenden Tieren zählen etwa Band- und Fadenwürmer, Dasselfliegen, Flöhe, Tierläuse oder Saugwürmer, die sich jedoch nur bei starkem Befall schwächend auf die Tiere auswirken.
Mensch und Paarhufer
Nutzung
 Einige Paarhufer wie das Hausschaf sind seit Jahrtausenden domestiziert.
Einige Paarhufer wie das Hausschaf sind seit Jahrtausenden domestiziert. Manche Arten wie das Dromedar werden auch als Last- und Reittier eingesetzt.
Manche Arten wie das Dromedar werden auch als Last- und Reittier eingesetzt.Schon seit frühester Zeit sind Paarhufer vom Menschen aus verschiedensten Gründen gejagt worden: um ihr Fleisch zu verzehren, ihr Fell zu Kleidung zu verarbeiten und ihre Stirnwaffen, Knochen und Zähne als Waffen oder Werkzeug zu verwenden. Später beschränkte sich der Mensch nicht nur auf die Jagd, sondern versuchte auch, einige Arten in seiner Nähe zu halten und nachzuzüchten. Die Domestizierung von Nutztieren begann zumindest im achten Jahrtausend v. Chr., als Wildziege, Wildschaf und Wildrind, etwas später auch das Wildschwein, zu Hausziege, Hausschaf, Hausrind und Hausschwein domestiziert worden sind. Nutztiere dienten zunächst vorwiegend als Nahrungsmittellieferanten, später wurden dann auch Tiere zur Arbeitstätigkeit eingesetzt, so seit rund 3000 vor Christus das Dromedar und das Lama. Der Prozess der Domestizierung verlief vielschichtig, genetische Studien deuten an, dass bei vielen Haustieren in unterschiedlichen Regionen dieser Schritt mehrmals unabhängig voneinander vonstatten ging.
Heute werden Paarhufer aus verschiedensten Gründen gehalten. Dies sind vorrangig der Genuss ihres Fleisches, die Gewinnung von Milch und die Verarbeitung ihrer Haut oder ihres Felles zu Leder und anderer Bekleidung oder die Schur zur Gewinnung der Wolle. Auch als Arbeits-, Zug-, Reit- oder Tragtiere werden manche Arten eingesetzt, etwa das Hausrind, der Wasserbüffel, der Yak oder verschiedene Kamele.
Hinsichtlich der Domestikation lassen sich zwei Grundtypen unterscheiden. Zum einen sind das Tiere, die in verschiedenen Rassen gezüchtet wurden, die weltweit verbreitet sind und die sich teilweise erheblich von der Wildform unterscheiden, etwa Hausrind, Hausschwein, Hausziege und Hausschaf. Andere Haustiere sind weitgehend in ihrem Ursprungsgebiet geblieben und gegenüber der Wildform wenig verändert, etwa das Rentier, einige Rinder (wie der Wasserbüffel, der Banteng, der Gaur oder das Yak) und Kamele (wie das Dromedar, das Trampeltier, das Lama oder das Alpaka).
Einige wildlebende Paarhuferarten werden nicht nur zum Nahrungserwerb, sondern auch aus jagdsportlichen Gründen erlegt. Solche Praktiken, die nicht aus Notwendigkeit, sondern zur Gewinnung von Trophäen durchgeführt werden, stehen unter teils heftiger Kritik und haben manche Arten, etwa den Alpensteinbock oder die Arabische Oryx, an den Rand der Ausrottung gedrängt.
Bedrohung
 Das Wildrind, die Stammform des Hausrindes, ist im 17. Jahrhundert ausgerottet worden.
Das Wildrind, die Stammform des Hausrindes, ist im 17. Jahrhundert ausgerottet worden.Der Gefährdungsgrad der einzelnen Paarhuferarten ist unterschiedlich. Einige Arten haben als Kulturfolger (wie etwa das Wildschwein) ihren Lebensraum ausbreiten können oder wurden vom Menschen als Parktiere oder entlaufene Haustiere in Regionen gebracht, in denen sie vorher nicht heimisch waren. Manche Paarhufer haben auch davon profitiert, dass ihre Fressfeinde (vorrangig Raubtiere) als Nahrungskonkurrenten der Viehzüchter von diesen teils erheblich dezimiert wurden.
Im Gegenzug sind viele Paarhufer in ihrem Bestand deutlich zurückgegangen und einige Paarhufer wurden sogar ausgerottet. Die Gründe dafür liegen in der Bejagung und in jüngerer Zeit auch in der zunehmenden Zerstörung ihres Lebensraumes. Ausgestorben sind mehrere Gazellenarten (die Arabische Gazelle und die Algerische Gazelle), mehrere madagassische Flusspferdarten, der Blaubock und der Schomburgk-Hirsch. Auch das Wildrind, die Stammform des Hausrindes, ist im 17. Jahrhundert verschwunden. Zwei Arten, die Arabische Oryx und die Saudi-Gazelle werden von der IUCN als in freier Wildbahn ausgestorben (extinct in the wild) geführt, das heißt, dass nur noch die Bestände in Nachzuchtprogrammen oder Tiergärten existieren. 14 Arten gelten als „vom Aussterben bedroht“ (critically endangered), darunter die Mendesantilope, der Kouprey, die Wildform des Trampeltiers, der Davidshirsch, die Przewalski-Gazelle, die Saiga, das Vietnamesische Waldrind und das Zwergwildschwein. Weitere 24 Arten werden als stark gefährdet (endangered) und 36 Arten als gefährdet (vulnerable) gelistet.[4]
Systematik und Stammesgeschichte
Die Systematik der Paarhufer wird heftig diskutiert. Grund dafür ist, dass sie einerseits eine morphologisch (von ihrem Körperbau) eindeutig definierte Ordnung sind, sich andererseits aber die Wale aus ihnen entwickelt haben und einige Gruppen (insbesondere die Flusspferde) näher mit diesen als mit den übrigen Paarhufern verwandt sind. Das macht die Paarhufer in phylogenetischen (über die Stammesentwicklung definierten) Systematiken – die in jüngerer Zeit maßgeblicher werden – zu einem paraphyletischen Taxon, das heißt zu einer Gruppe, die zwar von einer gemeinsamen Stammform abstammt, aber nicht alle Nachkommen dieses Vorfahren umfasst. Da die phylogenetische Systematik nach Möglichkeit nur monophyletische Taxa anerkennt, das heißt Gruppen, die von einer gemeinsamen Stammform abstammen und alle Nachkommen dieses Vorfahren umfassen, müssen die Paarhufer mit den Walen zu einem als Cetartiodactyla bezeichneten Taxon zusammengefasst werden. Hier soll zunächst die traditionelle Systematik vorgestellt und dann die phylogenetische Sicht der Paarhufer erläutert werden.
Die traditionelle Systematik
Die traditionelle Systematik der Paarhufer
 Richard Owen prägte den Begriff Paarhufer.
Richard Owen prägte den Begriff Paarhufer.Schon Carl von Linné postulierte eine enge Verwandtschaft zwischen Kamelen und Wiederkäuern. Henri de Blainville erkannte den ähnlichen Bau der Gliedmaßen dieser Tiere mit denen der Schweine und Flusspferde und der englische Zoologe Richard Owen prägte 1848 die Bezeichnung „even-toed ungulates“ („Paarzehige Huftiere“) und den wissenschaftlichen Namen Artiodactyla.
Seit dieser Zeit war die Zusammensetzung dieser Gruppe klar und wurde kaum jemals in Zweifel gezogen. Für die innere Systematik dienten der Bau des Magens und der Backenzähne. So haben Schweine, Pekaris und Flusspferde niederkronige Backenzähne und einen einfachen Magen, sie verdauen direkt, ohne wiederzukäuen. Darum wurden sie als Nicht-Wiederkäuer (Nonruminantia) oder Schweineartige im weiteren Sinn (Suina oder Neobunodontia) zusammengefasst. Alle anderen Paarhufer haben hochkronige Backenzähne mit selenodontem Bau (halbmondförmigen Schmelzleisten) und besitzen die Fähigkeit zum Wiederkäuen, sie bilden deshalb die Gruppe der Selenodontia. Unterschiede im Bau des Magens ließen erahnen, dass sich die Fähigkeit zum Wiederkäuen zweimal unabhängig voneinander entwickelt hat; deshalb werden die Kamele nicht zu den eigentlichen Wiederkäuern (Ruminantia) gezählt, sondern diesen als Schwielensohler (Tylopoda) gegenübergestellt. Innerhalb der Wiederkäuer stehen die urtümlichen, stirnwaffenlosen Hirschferkel allen anderen Gruppen gegenüber, die als Stirnwaffenträger (Pecora) zusammengefasst werden.
Aus rein morphologischen Gesichtspunkten ergaben sich daher folgende vermutete Abstammungsverhältnisse, die bis Ende des 20. Jahrhunderts weitgehend anerkannt waren:[5]
Paarhufer Schweineartige im weiteren Sinn (Suina/Neobunodontia) Schweineartige im engeren Sinn (Suoidea)
Flusspferde (Hippopotamidae)
Selenodontia Schwielensohler (Tylopoda)
Wiederkäuer (Ruminantia) Hirschferkel (Tragulidae)
Stirnwaffenträger (Pecora)
Die traditionelle Stellung der Wale
 Die Mesonychia, fleischfressende Huftiere aus dem frühen Känozoikum, galten lange Zeit als Vorfahren der Wale.
Die Mesonychia, fleischfressende Huftiere aus dem frühen Känozoikum, galten lange Zeit als Vorfahren der Wale.Die rezenten Wale sind hochangepasste Meeresbewohner, die äußerlich wenig Gemeinsamkeiten mit anderen Säugetieren haben – Ähnlichkeiten mit anderen Meeressäugern wie Robben und Seekühen beruhen ausschließlich auf Konvergenz. Es liegt aber nahe, dass sie sich aus landbewohnenden Säugern entwickelt haben müssen. Als wahrscheinlichste Kandidaten für die Vorfahren der Wale galten lange Zeit die Mesonychia. Das waren zum Teil riesenhafte, fleischfressende Tiere aus dem frühen Känozoikum (Paläozän und Eozän), die an den Füßen Hufe statt Krallen trugen. Ihre Gliedmaßen wiesen nicht den paarhufertypischen Bau des Sprungbeins auf, der auch von fossilen Walen bis vor kurzem nicht bekannt war. Ihre Backenzähne waren an eine tierische Ernährung angepasst und ähneln den Zähnen heutiger Zahnwale, die für eine fischfressende Nahrung ausgerichtet sind und im Gegensatz zu den übrigen Säugetieren einen gleichförmigen (homodonten) Bau aufweisen.
Man hielt die Mesonychia für nahe Verwandte der Paarhufer, sodass durchaus anerkannt war, dass Paarhufer und Wale die jeweils nächsten lebenden Verwandten voneinander sind. Diese nahe Verwandtschaft konnte auch durch morphologische Gemeinsamkeiten, etwa im Bau des Penis oder der Anordnung der Bronchien bestätigt werden. Die vermuteten Abstammungsverhältnisse lassen sich wie folgt wiedergeben:[6]
Paraxonia Paarhufer (Artiodactyla)
Cete Mesonychia †
Wale (Cetacea)
Die phylogenetische Systematik
Forschungsgeschichte
 Durch molekulare und auch morphologische Studien wurde bestätigt, dass die Wale die nächsten lebenden Verwandten der Flusspferde sind.
Durch molekulare und auch morphologische Studien wurde bestätigt, dass die Wale die nächsten lebenden Verwandten der Flusspferde sind.In den 1990er-Jahren wurde begonnen, die biologische Systematik nicht nur nach Gesichtspunkten des Körperbau und des Fossilbefundes, sondern auch mittels molekularbiologischer Studien zu erarbeiten. Dabei wird versucht, durch Sequenzierung der DNA und RNA genetische Informationen zu erlangen und mit den Daten anderer Lebewesen zu vergleichen, um anhand des Ähnlichkeitsgrades Hinweise auf den Verwandtschaftsgrad zu eruieren. Diese Methode wurde und wird bei zahlreichen Lebewesen angewandt und hat die Systematik vieler Taxa deutlich verändert. Auch bei den Paarhufern und Walen wurden diese Methoden durchgeführt, mit dem überraschenden Ergebnis, dass die nächsten Verwandten der Wale die Flusspferde und die Paarhufer somit eine paraphyletische Gruppe sind.
Zu den ersten, die zu diesem Ergebnis kamen, zählten Dan Graur und Desmond Higgins mit einer 1994 veröffentlichten Studie. [7] Allerdings berücksichtigten sie die Flusspferde noch nicht und hielten die Wiederkäuer für die Schwestergruppe der Wale. Nachfolgende Untersuchungen kamen dann zu dem Ergebnis, dass die Flusspferde die nächsten lebenden Verwandten der Wale darstellen, dies wurde unter anderem anhand von Caseingenen[8], SINEs[9], Fibrinogen-Sequenzen[10], Cytochrom- und rRNA-Sequenzen [11], IRBP- und vWF-Gensequenzen[12], Adrenorezeptoren[13] und Apolipoproteinen[14] bestätigt. In einer dieser Studien wurde von Claudine Montgelard, Francois M. Catzeflis und Emmanuel Douzery 1997 erstmals der Name Cetartiodactyla vorgeschlagen, der sich aus den wissenschaftlichen Bezeichnungen der Wale (Cetacea) und Paarhufer (Artiodactyla) zusammensetzt.
Im Jahr 2001 sorgten zwei außergewöhnliche Fossilfunde für Aufsehen. In Pakistan fand man Teile des Gliedmaßenskeletts des etwa wolfgroßen Pakicetus und des fuchsgroße Ichthyolestes, zwei urtümlichen Walen aus dem Eozän vor rund 48 Millionen Jahren, die als Pakicetidae zusammengefasst werden. Diese Funde zeigten nicht nur, dass die frühen Wale in stärkerem Ausmaß als bisher angenommen landgebunden waren, sondern wiesen eindeutig auch den speziellen Bau des Sprungbeins mit einer doppelt gerollten Gelenkfläche auf. Dieses Merkmal galt bislang als Exklusivmerkmal der Paarhufer und da es nun auch bei frühen Walen entdeckt wurde, konnte die enge Verwandtschaft beider Gruppen auch morphologisch belegt werden. (Bei den späteren Walen kam es zu einer so umfassenden Reduktion der Hintergliedmaßen, sodass aus dem Bau der Hinterbeine dieser Tiere keine Rückschlüsse auf mögliche Abstammungen mehr gezogen werden können.) Die Mesonychia zeigen diesen speziellen Bau des Sprungbeins nicht, somit war eine Abstammung der Wale von ihnen ausgeschlossen.
Der spezielle Bau des Sprungbeins belegte zwar eine enge Verwandtschaft zwischen Paarhufern und Walen, kann aber die Frage, ob die Paarhufer paraphyletisch sind, nicht beantworten. Darum wurden morphologische Untersuchungen durchgeführt, um den molekularbiologischen Befund der Nahverwandtschaft von Flusspferden und Walen zu unterstützen. In der Anordnung der Höcker der Backenzähne, im Bau der Mittelfußknochen und des Schädels konnten Übereinstimmungen gefunden werden,[15] die ein Schwestergruppenverhältnis dieser zwei Taxa unterstützen. Ob die auffälligste Gemeinsamkeit, der Verlust des Felles und der Talgdrüsen, ein gemeinsames Merkmal oder eine unabhängig voneinander entwickelte Anpassung an die wasserbewohnende Lebensweise ist, ist umstritten.
 Die Flusspferde sind eine erdgeschichtlich junge Gruppe, was Fragen über ihre Herkunft aufwirft.
Die Flusspferde sind eine erdgeschichtlich junge Gruppe, was Fragen über ihre Herkunft aufwirft.Als problematisch erweist sich dabei, dass der älteste Vertreter der Wale im frühen Eozän (vor rund 53 Millionen Jahren), das älteste bekannte Flusspferd aber erst im Miozän (vor rund 15 Millionen Jahren) gelebt hat. Da der gemeinsame Vorfahr von Walen und Flusspferden vor den ersten Walen gelebt haben muss, ergibt sich eine 40 Millionen Jahre lange Lücke der Fossilgeschichte der Flusspferde. In Anbetracht der vergleichsweise guten Fossilfundrate der Paarhufer, erscheint es unwahrscheinlich, dass es ausgerechnet von Vorfahren der Flusspferde keine Überreste gibt. (Manche Untersuchungen erklärten das späte Auftauchen der Flusspferde damit, dass sie sich aus Verwandten der Nabelschweine entwickelt hätten, das erscheint aber aufgrund des molekularen Befundes als unwahrscheinlich.) Das Augenmerk der Forschung richtete sich daher auf die Anthracotheriidae, einer vom Eozän bis in das Miozän verbreiteten Paarhufergruppe, die bereits bei ihrer Entdeckung im 19. Jahrhundert als „flusspferdähnlich“ beschrieben wurde. Eine Studie aus dem Jahr 2005 [16] zeigte, dass die jüngsten Vertreter der Anthracotheriidae starke Ähnlichkeiten mit den Flusspferden aufweisen. Als mögliches Szenario gilt dementsprechend, dass die Wale und die Anthracotheriidae von einem gemeinsamen Vorfahren abstammen und die Flusspferde sich aus den Anthracotheriidae entwickelten.
Innere Systematik
Der überwältigende molekulare Befund und auch einige morphologische Hinweise sprechen dafür, dass die Paarhufer paraphyletisch in Bezug auf die Wale sind und mit ihnen ein Taxon Cetartiodactyla bilden, wobei die Monophylie der Cetartiodactyla insgesamt durch molekulare und anatomische Hinweise gut abgesichert ist. Moderne Systematiken teilen die Cetartiodactyla in fünf untergeordnete Taxa, die jeweils mit hoher Wahrscheinlichkeit ebenfalls monophyletisch sind: die Schwielensohler (Tylopoda), die Schweineartigen (Suina), die Wiederkäuer (Ruminantia), die Flusspferde (Ancodonta) und die Wale (Cetacea).
Als Schwestergruppe der Wale gelten die Flusspferde – unter Einbeziehung fossiler Taxa dürfte eine Klade aus Flusspferden und Anthracotheriidae die nächste Verwandtschaftsgruppe dieser Meeressäuger darstellen. Auch die Wiederkäuer dürften näher mit Walen und Flusspferden als mit den übrigen Paarhufern verwandt sein – dies wurde bislang nur molekularbiologisch, aber nicht morphologisch untersucht und ist daher umstritten. Als älteste rezente Seitenlinie der Paarhufer gelten die Kamele, womit die Vermutung von der konvergenten Entwicklung der Fähigkeit zum Wiederkäuen bei Kamelen und Wiederkäuern bestätigt wird. Die vermuteten Abstammungslinien innerhalb der Cetartiodactyla lassen sich in folgendem Kladogramm wiedergeben[17]:
Cetartiodactyla Schwielensohler (Tylopoda)
N.N. Schweineartige (Suina)
Cetruminantia Wiederkäuer (Ruminantia) Hirschferkel (Tragulidae)
Stirnwaffenträger (Pecora)
Cetancodonta/Whippomorpha Flusspferde (Hippopotamidae)
Wale (Cetacea)
 Die Kamele gelten heute als Schwestergruppe aller übrigen Paarhufer.
Die Kamele gelten heute als Schwestergruppe aller übrigen Paarhufer. Der Gabelbock ist der einzige rezente Vertreter der Gabelhornträger.
Der Gabelbock ist der einzige rezente Vertreter der Gabelhornträger.Die vier als Paarhufer zusammengefassten Taxa der Cetartiodactyla teilen sich in zehn rezente Familien:
- Die Schwielensohler (Tylopoda) umfassen nur eine Familie, die Kamele (Camelidae). Es handelt sich um eine artenarme Gruppe von Tieren, die gut an extreme Habitate angepasst sind – die Altweltkamele an Wüsten und die Neuweltkamele an Hochgebirgsregionen.
- Die Schweineartigen (Suina) setzen sich aus zwei Familien zusammen:
- Die Echten oder Altweltschweine (Suidae) sind auf die Alte Welt beschränkt. Dazu gehören das Wildschwein und dessen domestizierte Form, das Hausschwein.
- Die Nabelschweine oder Pekaris (Tayassuidae) sind nach Drüsen an ihrem Bauch benannt und kommen heute in Mittel- und Südamerika vor.
- Die Wiederkäuer (Ruminantia) bestehen aus sechs Familien:
- Die Hirschferkel (Tragulidae) sind die kleinsten Paarhufer und die urtümlichsten Wiederkäuer, sie bewohnen Wälder in Afrika und Asien.
- Die Giraffenartigen (Giraffidae) setzen sich aus zwei äußerlich unterschiedlichen Arten zusammen, der Giraffe und dem Okapi.
- Die Moschushirsche oder Moschustiere (Moschidae) sind eine in Ostasien lebende Gruppe hirschähnlicher, aber geweihloser Tiere.
- Die Gabelhornträger (Antilocapridae) umfassen nur eine Art, den in Nordamerika lebenden Gabelbock.
- Die Hirsche (Cervidae) setzen sich aus rund 45 Arten zusammen, die durch ein Geweih charakterisiert sind, das in der Regel nur die Männchen tragen. Zu den auch in Europa verbreiteten Arten zählen unter anderem das Reh, Dam- und Rothirsch, der Elch und das Rentier.
- Die Hornträger (Bovidae) sind die artenreichste und vielgestaltigste Paarhufergruppe. Zu ihnen zählen die Rinder, die Ziegenartigen, die Gazellenartigen und mehrere als Antilopen bezeichnete Gruppen.
- Die Flusspferde (Hippopotamidae) umfassen zwei Arten, das (Eigentliche) Flusspferd und das Zwergflusspferd. Sie werden aus den oben genannten Gründen in einer eigenen Unterordnung, Ancodonta, geführt.
- Für die interne Systematik der Wale siehe Wale#Systematik.
Das größte systematische Problem innerhalb der untergeordneten Taxa betrifft die Wiederkäuer. Allgemein anerkannt ist, dass die Hirschferkel die Schwestergruppe der übrigen fünf Familien sind, die als Stirnwaffenträger (Pecora) zusammengefasst werden. Innerhalb der Stirnwaffenträger ist die Systematik unübersichtlich. Zwar wurden traditionell Hirsche, Moschushirsche und Gabelhornträger als Hirschartige (Cervioidea) zusammengefasst, verschiedene molekulare Untersuchungen liefern jedoch andere – und uneinheitliche – Ergebnisse, sodass die Frage nach einer phylogenetischen Systematik der Stirnwaffenträger zum gegenwärtigen Zeitpunkt nicht beantwortet werden kann.
Im Dezember 2007 stellte Hans Thewissen, Professor am Department of Anatomy der Northeastern Ohio Universities Colleges of Medicine and Pharmacy, eine alternative Stammbaumhypothese vor. Seinen Untersuchungen entsprechend waren die nächsten Verwandten der frühen Wale eine ausgestorbene Paarhufergruppe namens Raoellidae und beide Taxa stellen zusammen die Schwestergruppe der restlichen Paarhufer dar, inklusive der Flusspferde:
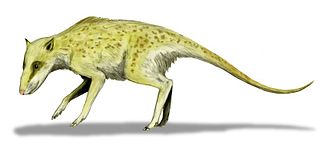 Indohyus in einer Lebendrekonstruktion
Indohyus in einer LebendrekonstruktionCetartiodactyla übrige Paarhufer (incl. Flusspferde)
N.N. Raoellidae (u.a. Indohyus und Khirharia)
Seine Erkenntnisse stammen aus der Untersuchung eines neuen Skeletts aus der Kaschmir-Region in Pakistan. Dabei handelte es sich um einen Vertreter der Gattung Indohyus, welche den Raoellidae zugeordnet wird. Vor allem aufgrund eines knöchernen Rings am Felsenbein (Bulla tympanica), dem so genannten Involucrum, welcher bislang nur von Walen bekannt war, sowie weiteren Merkmalen der Vorbackenzähne (Prämolare) und der Knochenstruktur, wurde die nahe Verwandtschaft begründet.[18]Äußere Systematik
Die Cetartiodactyla werden innerhalb der Höheren Säugetiere in die Überordnung der Laurasiatheria eingeordnet, eine Säugergruppe, die nach ihrem vermutlichen Ursprungsort, dem Kontinent Laurasia, benannt ist. Mit welchen Großgruppen innerhalb der Laurasiatheria die Cetartiodactyla näher verwandt sind, ist immer noch ungeklärt, als gesichert gilt allerdings, dass die früher vorgeschlagene Gruppe der „Huftiere“ (Ungulata) keine natürliche Gruppe darstellt. Verschiedene molekulare Untersuchungen lassen zumindest unterschiedliche mögliche Abstammungslinien erkennen: Nach einer Hypothese bilden die Cetartiodactyla gemeinsam mit den Unpaarhufern und den Ferae (Schuppentiere und Raubtiere) ein gemeinsames Taxon Fereuungulata – wobei unklar bleibt, ob die Unpaarhufer näher mit den Cetartiodactyla oder den Ferae verwandt sind. Eine andere, in jüngerer Zeit aufgestellte Hypothese fasst hingegen Unpaarhufer, Ferae und Fledertiere zu einem Taxon Pegasoferae zusammen und sieht die Cetartiodactyla als deren Schwestergruppe.[19]
Stammesgeschichte
Die ältesten Fossilien der Paarhufer stammen aus dem frühen Eozän (vor rund 55 Millionen Jahren). Da diese Funde nahezu zeitgleich in Europa, Asien und Nordamerika auftauchten, ist es sehr schwierig, den Entstehungsort der Paarhufer genauer zu bestimmen. Diese frühen Formen werden in der Gruppe der Dichobunidae (oder Dichobunoidea) zusammengefasst – ihr bekanntester und am besten erhaltener Vertreter ist Diacodexis. Es handelt sich um kleine (anfangs nur etwa hasengroße) Tiere mit schlankem Körperbau, langen dünnen Beinen und einen langen Schwanz. Die Hinterbeine waren deutlich länger als die Vorderbeine, die typische Zehenstruktur noch nicht voll entwickelt und die Zähne niederkronig und einfach gebaut. Schon im frühen bis mittleren Eozän kam es zur Radiation und zur Entstehung der Vorfahren der meisten heutigen Unterordnungen. Die Stammesgeschichte der Paarhufer verlief allerdings verhältnismäßig gleichförmig und unspektakulär, riesenhafte oder bizarr anmutende Tiere (wie sie etwa bei den Unpaarhufern entstanden), sind aus dieser Gruppe kaum bekannt.
Die frühesten Wale sind aus dem frühen Eozän (vor rund 53 Millionen Jahren) belegt und haben sich vermutlich auf dem indischen Subkontinent entwickelt. Von da an schlugen sie, bedingt durch die meeresbewohnende Lebensweise, eine gänzliche eigene Entwicklungslinie ein (siehe Evolution der Wale).
 Die Entelodontidae waren stämmige Tiere mit großem Kopf, der durch Knochenhöcker am Unterkiefer gekennzeichnet war.
Die Entelodontidae waren stämmige Tiere mit großem Kopf, der durch Knochenhöcker am Unterkiefer gekennzeichnet war.Zwei einstmals verbreitete, heute aber ausgestorbene Gruppen der Paarhufer waren die Entelodontidae und die Anthracotheriidae. Die Entelodontidae existierten vom mittleren Eozän bis zum frühen Miozän in Eurasien und Nordamerika. Sie hatten einen stämmigen Körper mit kurzen Beinen und einen massiven Kopf, der durch zwei Knochenhöcker am Unterkiefer gekennzeichnet war. Die Antracotheriidae wiesen einen großen, schweineartigen Körperbau auf, die Beine waren kurz und die Schnauze auffallend langgestreckt. Diese Gruppe erschien im mittleren Eozän und verbreitete sich über Eurasien, Afrika und Nordamerika. Sie lebte bis in das Miozän oder Pliozän – aus Asien gibt es allerdings noch einen unsicheren Fund aus dem frühen Pleistozän. Vermutlich waren die Anthracotheriidae die Vorfahren der Flusspferde, vielleicht führten sie eine ähnliche wasserbewohnende Lebensweise und wurden von diesen verdrängt. Die Flusspferde selbst erschienen im späten Miozän und sind aus Afrika und Asien belegt – Amerika haben sie nie erreicht.
Die Schwielensohler (Tylopoda) waren während weiter Teile des Känozoikums auf Nordamerika beschränkt, Frühformen wie die Cainotheriidae sind aber auch aus Europa belegt. Zu den nordamerikanischen Schwielensohlern zählten Gruppen wie die stämmigen, kurzbeinigen Merycoidodontidae, die mit Stirnwaffen ausgestatteten Protoceratidae und die eigentlichen Kamele (Camelidae). Diese erschienen erstmals im späten Eozän und entwickelten in Nordamerika einen großen Artenreichtum. Erst im späten Miozän oder frühen Pliozän wanderten sie in Eurasien und im späten Pliozän in Südamerika ein. In diesen Kontinenten leben sie bis heute, in Nordamerika sind sie aus nicht genau bekannten Gründen vor rund 10.000 Jahren ausgestorben.
Vertreter der Schweineartigen (Suina) sind seit dem Eozän bekannt. Im späten Eozän oder Oligozän bildeten sich die beiden heute noch bestehenden Familien, die Altweltschweine, die stets auf Eurasien und Afrika beschränkt blieben und die Nabelschweine, die in der Alten Welt ausstarben und heute nur mehr in Amerika vorkommen.
 Die Rindergiraffen (Sivatheriinae) waren eine Gruppe kurzhalsiger Giraffenartiger mit hirschähnlichen Stirnwaffen.
Die Rindergiraffen (Sivatheriinae) waren eine Gruppe kurzhalsiger Giraffenartiger mit hirschähnlichen Stirnwaffen.Auch die Wiederkäuer (Ruminantia) sind mit zahlreichen frühen Formen, die wohl den heutigen Hirschferkeln ähneln, seit dem Eozän bekannt. Diese Frühformen, die als Tragulina zusammengefasst werden, lebten bis zum Miozän in Afrika, Eurasien und Nordamerika, dann starben sie bis auf die rezenten Hirschferkel aus. Im Oligozän oder Miozän entwickelten sich die übrigen heute noch lebenden Vertreter, die mehrheitlich auf die Alte Welt beschränkt blieben. Eine große Radiation erfuhren die Hornträger (Bovidae), die ab dem frühen Miozän Afrika besiedelten, wo sie die früher dominanten Unpaarhufer und Schliefer weitgehend verdrängten und die ökologische Nische der großen Pflanzenfresser besetzten. Innerhalb der Giraffenartigen (Giraffidae) entstanden neben den heutigen Arten auch die Rindergiraffen, die hirschähnliche Stirnwaffen entwickelten. Auch die Hirsche (Cervidae) entwickelten zahlreiche Arten, blieben jedoch eher auf Eurasien beschränkt. Nach Nordamerika kamen Hornträger und Hirsche erst relativ spät (Spätmiozän oder Frühpliozän), auf diesem Kontinent hatte sich aber die Gruppe der Gabelhornträger mit zahlreichen Arten ausgebreitet, von welchen heute nur mehr eine Art, der Gabelbock überlebt hat.
Südamerika wurde von den Paarhufern erst im Pliozän vor rund drei Millionen Jahren besiedelt, als sich die Landverbindung beim Isthmus von Panama schloss. Mit Nabelschweinen, Neuweltkamelen und Trughirschen ist die südamerikanische Paarhuferfauna verglichen mit den anderen Kontinenten aber artenarm geblieben. Vom Aussterben der Großsäuger am Ende des Pleistozäns waren die Paarhufer weniger betroffen als andere Säugergruppen, ein Artensterben größeren Ausmaßes geschah nur bei den nordamerikanischen Kamelen. Ein Grund hierfür liegt darin, dass die größeren Vertreter der Paarhufer in Afrika vorkamen und dort im Gegensatz zu den anderen Kontinenten zur damaligen Zeit kaum Großsäuger ausstarben.
Belege
Zitierte Belege
Die Informationen dieses Artikels entstammen zum größten Teil den unter Literatur angegebenen Quellen, darüber hinaus werden folgende Quellen zitiert:
- ↑ Hendrichs (2004), S. 609
- ↑ Franz-Viktor Salomon: Bewegungsapparat. In: F.-V. Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. Enke, Stuttgart, 2. erw. Aufl. 2008, S. 22–234. ISBN 978-3-8304-1075-1.
- ↑ Uwe Gille: Harn- und Geschlechtsapparat, Apparatus urogenitalis. In: F.-V. Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. Enke, Stuttgart, 2. erw. Aufl. 2008, S. 368–403. ISBN 978-3-8304-1075-1
- ↑ Gefährdungsgrad der einzelnen Arten in der Roten Liste gefährdeter Arten der IUCN. Abgerufen am 12. März 2007
- ↑ etwa noch bei Nowak (1999) oder Hendrichs (2004)
- ↑ folgend etwa Malcolm C. McKenna, Susan K. Bell: Classification of Mammals - Above the Species Level. Columbia University Press, New York 1997. ISBN 0-231-11013-8
- ↑ Dan Graur and Desmond G. Higgins: Molecular Evidence for the Inclusion of Cetaceans within the Order Artiodactyla. In: Molecular Biology and Evolution, 11(3) 357-364 (1994). PDF
- ↑ John Gatesy, Cheryl Hayashi, Mathew A. Cronin, Peter Arctander: Evidence from milk casein genes that cetaceans are close relatives of hippopotamid artiodactyls. In: Molecular Biology and Evolution, 13(7): 954-963 (1996).
- ↑ M. Shimamura et al: Molecular evidence from retroposons that whales form a clade within even-toed ungulates. in: Nature, 388: 666-670 (1997) Abstract
- ↑ John Gatesy: More DNA Support for a Cetacea/Hippopotamidae Clade: The Blood-Clotting Protein Gene y-Fibrinogen. in: Molecular Biology and Evolution, 14(5): 537-543 (1997)
- ↑ Claudine Montgelard, Francois M. Catzeflis und Emmanuel Douzery: Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. In: Molecular Biology and Evolution, 14(5): 550-559 (1997). PDF
- ↑ John Gatesy, Michel Milinkovitch, Victor Waddell und Michael Stanhope: Stability of Cladistic Relationships between Cetacea and Higher-Level Artiodactyl Taxa. In: Systematic Biology, 48(1): 6-20 (1999). Abstract
- ↑ Ole Madsen, Diederik Willemsen, Björn M. Ursing, Ulfur Arnason und Wilfried W. de Jong: Molecular Evolution of the Mammalian Alpha 2B Adrenergic Receptor. In: Molecular Biology and Evolution 19: 2150-2160 (2002).
- ↑ Heather Amrine-Madsen, Klaus.-P. Koepfli, Robert K. Wayne und Mark S. Springer: A new phylogenetic marker, apolipoprotein B, provides compelling evidence for eutherian relationships. In: Mol. Phylogenet. Evol. 28: 225-240 (2003). Abstract
- ↑ Jonathan Geilser und Mark Uhen: Morphological Support for a close Relationship between Hippos and Whales. In: Journal of Vertebrate Paleontology 23(4):991–996 (2003).
- ↑ Jean-Renaud Boisserie, Fabrice Lihoreau und Michel Brunet: The position of Hippopotamidae within Cetartiodactyla. In: Proceedings of the National Academy of Sciences of the USA 102: 1537 – 1541 (2005). Fulltext
- ↑ Nach Robin Beck et al.: A higher-level MRP supertree of placental mammals. In: BMC Evol Biol. 2006; 6: 93. Fulltext
- ↑ J.G.M. Thewissen, Lisa Noelle Cooper, Mark T. Clementz, Sunil Bajpai, B.N. Tiwari: Whales orginated from aquatic artiodactyls in the Eocene epoch of India. Nature 450, 2007; Seiten 1190–1194
- ↑ Hidenori Nishihara, Masami Hasegawa und Norihiro Okada: Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions, in Proceedings of the National Academy of Sciences 103, 2006; Seiten 9929-9934 (Volltext, PDF verfügbar)
Literatur
- Hubert Hendrichs: Artiodactyla (Paraxonia), Paarhufer. In: Wilfried Westheide, Reinhard Rieger (Hrsg.): Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. Spektrum Akademischer Verlag, Heidelberg – Berlin 2004, S. 608–630, ISBN 3-8274-0307-3. (vorwiegend Körperbau und Lebensweise)
- Thomas S. Kemp: The Origin & Evolution of Mammals. Oxford University Press, Oxford 2005, 331 Seiten, ISBN 0-19-850761-5. (vorwiegend Systematik und Stammesgeschichte)
- Kenneth D. Rose und David Archibald: Rise of Placental Mammals: Origins and Relationships of the Major Extant Clades. Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8022-X (vorwiegend Systematik und Stammesgeschichte)
- Don E. Wilson, DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Ausgabe. Johns Hopkins University Press, Baltimore 2005, ISBN 0-8018-8221-4. (Artverzeichnis)
- Ronald M. Nowak: Walker's Mammals of the World. The Johns Hopkins University Press, Baltimore 1999, 2015 Seiten, ISBN 0-8018-5789-9. (allgemeine Informationen)
Weblinks
 Commons: Paarhufer – Sammlung von Bildern, Videos und Audiodateien
Commons: Paarhufer – Sammlung von Bildern, Videos und Audiodateien Wiktionary: Paarhufer – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen
Wiktionary: Paarhufer – Bedeutungserklärungen, Wortherkunft, Synonyme, Übersetzungen- Artiodactyla und Cetartiodactyla auf Animal Diversity Web (allgemeine Informationen)


Dieser Artikel wurde am 12. Februar 2010 in dieser Version in die Liste der exzellenten Artikel aufgenommen.






















Wikimedia Foundation.